
球状タンパク質の疎水性コアの奥深く、水分子が一度も侵入したことのない幾何学的な闇の中に、あなたは存在している。ロイシンやバリンのファンデルワールス球面が四方八方から押し迫り、炭灰色と温かみのある象牙色の表面が量子反発の極薄い余白だけを残して互いに接触し、二つの電子雲が触れ合うたびにかすかな接触ハローを放ちながら、地質学的な洞窟のように完璧に噛み合う。フェニルアラニンの芳香環が黒曜石の平板として空間を鋭く分断し、その影は拡散光の存在しないこの世界でナイフの刃のように絶対的な暗さを刻み込んでいる。散在するメチオニンの硫黄原子が、割れた岩の隙間から覗く熾火のような深い硫黄色の輝きを点在させ、無彩色の世界に唯一の温色をもたらしている。遠方の溶媒境界からは、封じられた洞窟の底に届く光のように、冷たい青緑色の光がタンパク質の二次構造の隙間をかすかに滲み込んでくる——それは深さと閉塞感を示唆するだけで、何かを照らすものではない。310ケルビンの熱振動がサブオングストローム単位の微かな拍動として全ての接触面を静かに伝わり、このテクトニックな静寂はキロジュール以下のファンデルワールス力によって保たれている。

ペプチド出口トンネルの奥深くから視線を向けると、世界はリボソームRNAの螺旋が織りなす大聖堂の回廊へと収束し、琥珀色と焦げた銅色の核酸建築が幾重にも積み重なったヌクレオチド基盤の棚となって壁面を刻んでいる。前方10ナノメートル先では、ペプチジル転移酵素センターが青白い炎のように燃え上がり、その光はリボソームRNAの放射状の柱を照らしながら、回廊の床に藍色と黄金色の縞模様を投げかけている。最も狭い箇所では対向する壁面が1.5ナノメートルにまで迫り合い、互いのファンデルワールス電子雲が淡い紫の接触光縁として溶け合いそうになっている。この場所は温度揺らぎと静電気的地形と疎水性相互作用が絶え間なく交差する圧縮された親密さの空間であり、新生ポリペプチド鎖がライム緑と琥珀色に輝く糸として催炎の中心から手繰り寄せられ、その側鎖が壁面のrRNA稜線にファンデルワールスの一瞬の接触を繰り返しながら、ある蛋白質の誕生の最初の瞬間を温かい琥珀の光の中で紡いでいる。

眼前に広がるのは、幅わずか2.2ナノメートルの螺旋状の峡谷——B型DNA二重らせんの主溝の内部であり、琥珀色のアデニン、淡い鼠緑色のチミン、深青緑色のグアニン、空色のシトシンが幾層にも積み重なった芳香族塩基対が、古代のモザイクタイルのように足元を覆いながら、ゆっくりと時計回りに螺旋を描いて遠方へと消えていく。塩基間のπ電子雲が重なり合い、薄く滲む紫紺色の量子干渉光が床の継ぎ目ごとに揺らめく一方、両壁を構成するリン酸–糖骨格は橙褐色の半透明な手すりとして高くそびえ、その外縁にはマグネシウムイオンが鋭く白青く輝く火花として、ナトリウムイオンが柔らかな淡金色のハローとして漂い、デバイ遮蔽層の乳白色の電場の霧の中に浮かんでいる。熱運動する水分子が絶え間なく塩基対の床に衝突しては弾け、瞬間的な白い閃光を散らしながら全体に低い振動を与え、構造全体が一フェムト秒の凍結した静寂の中に鋭く切り取られている。ここでは「表面」とは明確な境界ではなく、原子半径の外縁で確率的な電子密度が柔らかな静電気の毛羽となって滲み出した、圧力として感じる前に触れることのできない、曖昧だが確かな実在である。

抗体の結合部位へと踏み込むとき、視界を支配するのは二枚の巨大な電子密度の崖壁であり、わずか数個の水分子の幅しか離れていない隙間を挟んで互いに引き寄せられている。くすんだ錫灰色のベータサンドウィッチが背後の砦のように屹立し、その頂から伸びる琥珀色のH3ループと青緑のL3ループが、対岸の抗原エピトープの凸状地形をゆっくりと包み込んでいく。接触面は5×6ナノメートルという小さな領域に凝縮されており、水分子が弾かれた火花のように外へ散乱しながら、シアンの水素結合の橋がひとつまたひとつと瞬き、電子密度の淡い光の糸として固定されていく。周囲を満たす溶媒の青白い散乱光の中で、芳香族側鎖は浅い皿状の窪みとして、リシン残基は細い鍾乳石として迫り出し、それぞれの分子表面が大陸衝突のごとくゆっくり融合する様子は、スケールの違いを超えた地質学的な荘厳さを帯びている。熱揺動が引き起こす絶え間ない衝撃の中で、この精密な認識の幾何学が確定的な抱擁へと収束していく過程こそ、免疫応答の分子的根拠そのものである。
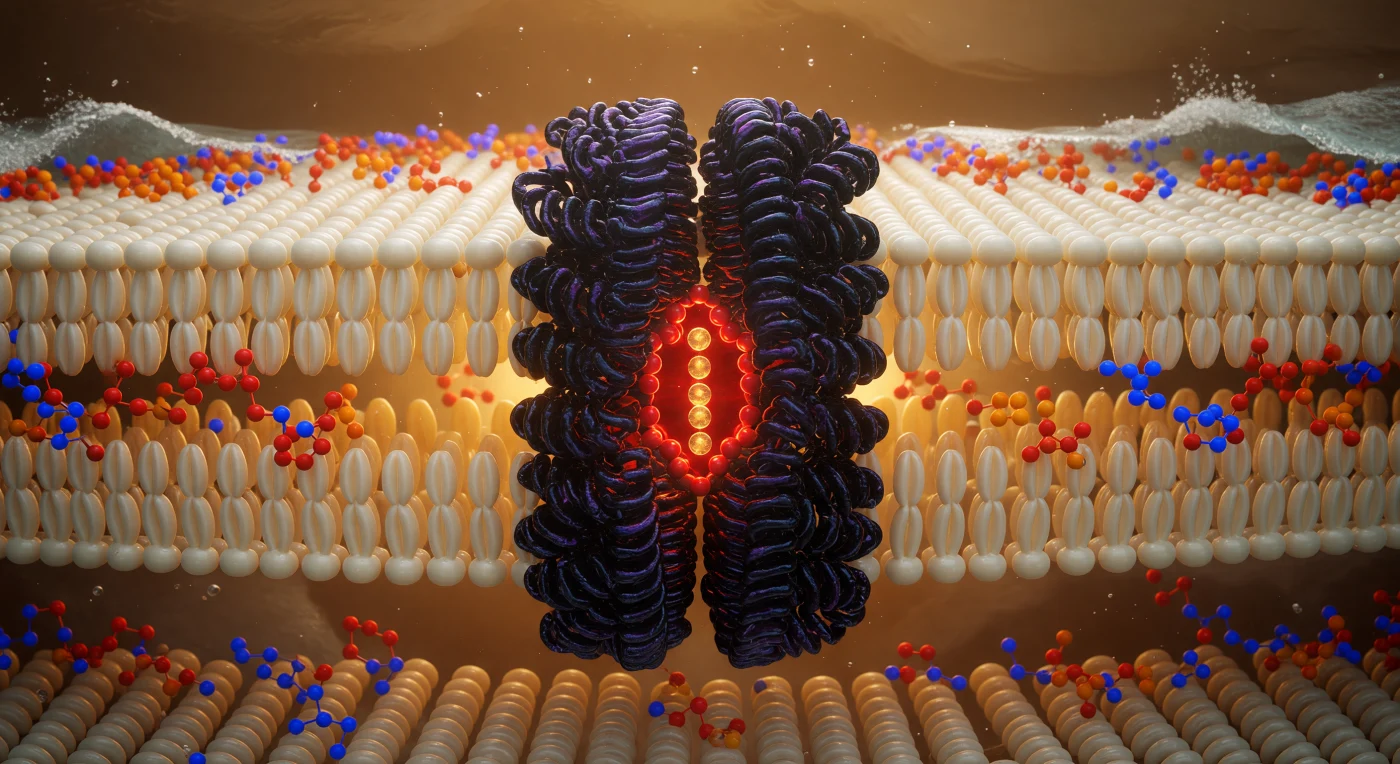
リン脂質二重層の幾何学的中心——ヘッドグループの岸辺からいずれも約2ナノメートルという深みに、あなたは漂っている。四方八方に伸びる脂肪酸鎖が象牙色と淡金色のファン・デル・ワールス面を密に折り重ね、その微細な熱振動が空間全体をとろけかけた琥珀色の霞で満たし、電子雲の静かな静電的輝きが唯一の光源となって周囲を照らしている。正面には四回対称のカリウムチャネルが玄武岩色の巨柱として二重層を貫き、その疎水性残基が周囲の脂質鎖と隙間なく噛み合うことで、タンパク質と膜の界面は風化した岩石が密林に溶け込むような一体の地形をなしている。孔軸の選択性フィルターでは、カルボニル酸素の環が赤橙色に燃え、カリウムイオンが温かみのある金色の球として一列縦隊で通過し、それぞれが脱水和の残光を纏いながら積み重なって立ち上る。見上げれば、リン原子の濃い橙と窒素原子の電気的な青が二本の発光した水際として輝き、その彼方では水分子の荒れ狂う銀白の泡沫が帯電した界面に砕け寄る波のように押し迫っている。

地平線のすべてに広がる波打つ平原の、ほぼ地表すれすれの高さから見渡すと、琥珀色の羊皮紙のようなバックボーン鎖が規則的な畝となって交互の向きに走り、その頂上には磨かれたガーネットのようなチェリーレッドのカルボニル酸素が並び、谷間には白く淡く発光するアミド窒素の先端が静かに光を帯びている。隣接するβ鎖の間には、シアンの水素結合が幽霊のような電磁気的なフィラメントとして横に架け渡され、2.9オングストロームという息が詰まるほどの近さでドナーとアクセプターの電子雲が重なり合い、この広大な平原全体を分子の鎖帷子のように縫い合わせている。目に映るこの全景は、タンパク質がもつ逆平行βシートの自己組織化した幾何学——熱力学的な圧力のもとで育まれた秩序であり、隣接するアミノ酸残基間の共有結合と非共有結合が織りなす微細な均衡の産物だ。平原の周縁では、整然たる畝が崩れ、赤とオレンジのコネクターループが凍りついた波のように乱れ広がり、秩序と無秩序との境界を刻んでいる。頭上には方向性のない水溶液の拡散光が満ちており、対イオンの雲と水分子の双極子が絶え間なく再配向するせいで、タンパク質と溶媒の境界は虹色にかすんでいる——この規模では、可視光の波長そのものが眼前の景観全体を丸ごと呑み込んでしまうほど巨大なのだ。

球体の内壁はあらゆる方向になだらかに湾曲し、その表面から滲み出る青白い冷たい光が内部全体を均一に満たしている——あなたは今、直径わずか8ナノメートルのGroEL-GroELシャペロニン折り畳み室の中心に浮かんでいる。周囲を取り囲む淡い灰白色の壁は親水性残基で覆われた滑らかな内膜であり、電子顕微鏡的な鮮明さでアルファヘリックスの緩やかな縦縞が刻まれ、側鎖の先端がビロードの毛並みのように内側へ向かって微細に突き出している。頭上ではGroESキャップが七つの銀色サブユニットを石畳のように組み合わせた浅いドーム天井を形成し、その継ぎ目は薄暗い灰の線として閉じた穹窿を描いている。室の中央には、3ナノメートルほどの誤って折り畳まれた基質タンパク質が——温かみのある琥珀色の絡まったリボン状の塊として——ブラウン運動によってゆっくりと漂っており、その内側から滲む橙色の光が壁の青白い放射と鮮やかに対比している。この密封された分子の穹窿の中で、ATPの加水分解サイクルが駆動するたびに内壁の性質が変化し、タンパク質に正しい立体構造へと折り畳み直す機会を与えるのである。

眼下に広がるのは、生命の複製装置の中枢——CMGヘリカーゼの巨大な六量体リングが、まるで石造りの水車のように二本鎖DNAをまたぎ、右側から届く深紺の親鎖を強引に解きほぐしながら、ティールと金色の一本鎖テンプレートを二筋の帯として傍らへ送り出している光景だ。自分はいま、温かく発光するゲル状の水性媒体の中に漂っており、青白い水分子が四方八方から絶え間なく押し寄せ、確固たる「足場」などというものは存在せず、熱揺らぎそのものが空間の質感をなしている。直下では、DNAポリメラーゼが灰褐色の巨大な拳のように鋳型鎖を握りしめ、ヌクレオチドが組み込まれるたびにオレンジ白色のパイロリン酸の閃光がフェムト秒単位で炸裂し、瞬時に水の熱場へと溶け消えてゆく——その放出エネルギーこそが、新生鎖の共有結合形成を熱力学的に不可逆にしている駆動力にほかならない。フォーク接合部では水素結合が確率的に崩壊し、塩基対が淡い金色の温光とともに解離する——それは切断ではなく、エネルギー地形の緩やかな溶解であり、毎秒約一千塩基対という驚異的な速度でゲノム情報を忠実に複写し続ける分子機械の、息をのむほど動的な閾値だ。中景には一本鎖結合タンパク質が半透明なクラゲのように漂い、露出した塩基を二次構造の罠から守りながら、銀白色のマグネシウムイオンが静電チャネルを伝って活性部位へと閃光のように出入りしている。

地面から見上げると、直径わずか1.2ナノメートル、高さ6ナノメートルの右巻きαヘリックスの柱が、まるで古代の森の樹木のように四方を取り囲んでいる。それぞれの柱は時計回りに螺旋を描き、表面には温かみのある琥珀色のロイシン残基が規則的に内側へ突き出して、隣接するヘリックスとコイルドコイルの縫い目を形成し、疎水性の接触面でファンデルワールス力によって互いを引き寄せ合っている。外側に向いたリシン残基は深い緋色に輝き、アルギニン残基は冷たい電気的な青色で脈動しながら、その静電気的なハローを周囲の水溶液へと滲み出させている。頭上に広がる水の空は、0.28ナノメートルの水分子が猛烈な速度で衝突し向きを変えるため、澄んだセルリアンブルーというより半透明な発光体のような霞として揺らめき、ピコ秒の瞬きで点滅する水素結合網が光の糸のように煌めく。20ナノメートル先の分子霞の奥へと目を向けると、コイルドコイルの縫い目が形成するヘリングボーン模様が次第に青灰色の靄に溶け込んでゆき、ここでは熱振動そのものが森全体を微かに、しかし絶え間なく呼吸させているかのようだ。

アミロイド線維の軸を真下に見下ろすとき、四枚の花弁のように広がる原線維が視野全体を満たし、それぞれ幅わずか3ナノメートルほどの構造体が鋼青色の冷たい輝きを放ちながら完璧な放射対称を描いている。各βストランドは4.7オングストロームという寸分の狂いもない間隔で積み重なり、その精密さはまるで結晶の律動のように感じられ、個々のアミノ酸残基が電子密度の小さな丘として浮き彫りになった表面は水分子を一切寄せ付けない。四枚の花弁が囲む中央の深みには琥珀色に発光するステリックジッパーの核心部が息づき、互いに指を組み合わせるように嚙み合ったβシートの側鎖がファン・デル・ワールス接触の限界まで迫り合い、ほとんど水の存在しない乾燥した静寂の空間を作り出している。線維の外縁では一転して、深紅に帯電したグルタミン酸の突起と電気的に青く輝くリシンの長い側鎖が溶媒側へ向かって伸び、誘電体の霞の中で激しい静電場を形成している。この絶対的秩序の凍った大聖堂は、かつて可溶性だったタンパク質が崩壊し固着した結果として生まれた構造であり、その永続性と対称性は石材よりも説得力を持って空間に刻み込まれている。
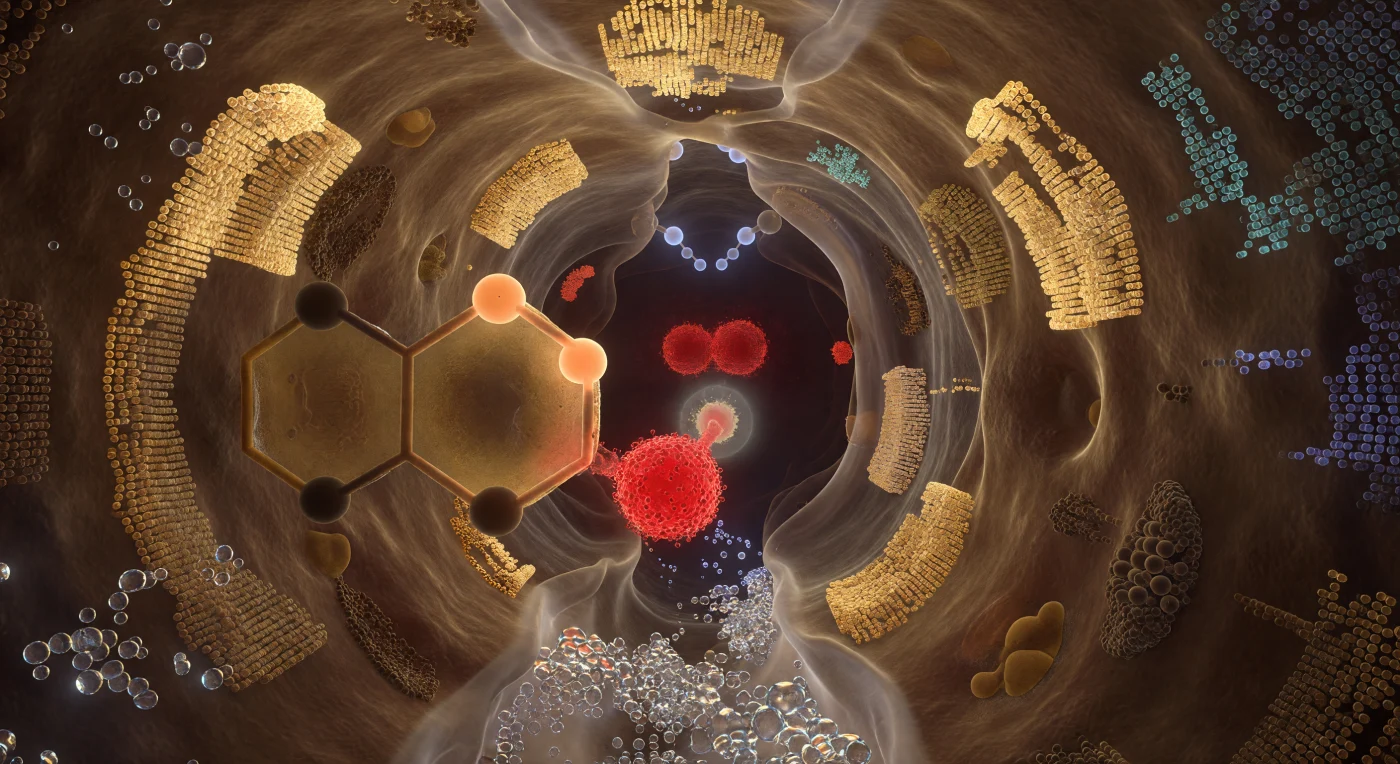
活性部位の裂け目の深部に立つと、タンパク質の壁が生きた琥珀と影から削り出された大聖堂の内部のように四方から湾曲して迫り、その表面は数百もの側鎖がファン・デル・ワールス力で緊密に充填された電子密度の濃い物質として半透明と不透明の間を揺れ、温かな金と淡硫黄色の内的な発光を帯びている。空間の中心では触媒三残基が地質学的な巨大建造物の重力で支配的に君臨し、Ser195の酸素が近景で赤熱した炭のように燃え、その孤立電子対の密度が緋色の霞を形成しながら基質の切断される結合のわずか1.5オングストローム先へと引き伸ばされ、His57のイミダゾール環が左の視野全体を温かな琥珀色のステンドグラスの巨大な芳香族パネルとして満たし、プロトン移動の途中で凍りついた移送中の水素がアプリコット色に輝いている。オキサニオンホールでは二つの主鎖N-Hドナーが反応軸の両側から薄青い水素原子を内向きに突き出してカルボニル炭素に冷白色の静電光を投じ、四面体中間体の形成をピンセットのように挟み込む。この反応室全体が——固体であると同時に溶解し、実在すると同時に幻影であるかのように——最外縁のオングストロームで電子確率の量子的な霧へと軟化しながら、化学の精密な一瞬に凍結されて息づいている。

目の前に広がるのは、三本のポリペプチド鎖が互いに絡み合いながら右巻きの超らせんを描くコラーゲン三重らせんの表面であり、暖かみのある象牙色、淡い金色、日に褪せた黄褐色のそれぞれの鎖が、まるで地質学的な時間をかけて編まれた縄のように、わずか1.5ナノメートルという細さの中で不可分に結び合っている。三残基ごとに規則正しく並ぶプロリンのピロリジン環が灰白色の関節のように超らせんの溝へわずかに沈み込み、その間からヒドロキシプロリンの水酸基が琥珀色の突起として外へ張り出し、それぞれに一分子の水が露のように貼り付いている。ケーブルの表面を覆う第一水和殻は淡い結晶質の青白色をした霜ガラスのような鞘をなしており、水分子が2.8オングストローム間隔の水素結合格子の中でほぼ完全な四面体配置に秩序付けられ、整列した双極子の青白い偏光をかすかに帯びている。視線を先へ向けると、ケーブルは幾百ナノメートルにもわたって薄青緑と銀白色の分子霧の中へと消えていき、その霧そのものが一立方ナノメートルあたり一万個の水分子が生み出す熱的な息吹であり、電荷を持つ主鎖の静電的な暈の中に漂う対イオン雲が霞に琥珀の色調を添えている。骨格のカルボニル酸素が小さな青みを帯びたくぼみとして、アミド窒素のプロトンがほんのりとクリーム色の突起として浮かび上がり、全体がサブオングストロームの熱振動による微かな揺らぎをまといながら、コラーゲンという生体構造の最も根源的な強靭さをこの極小の回廊の奥深くへと静かに刻み込んでいる。
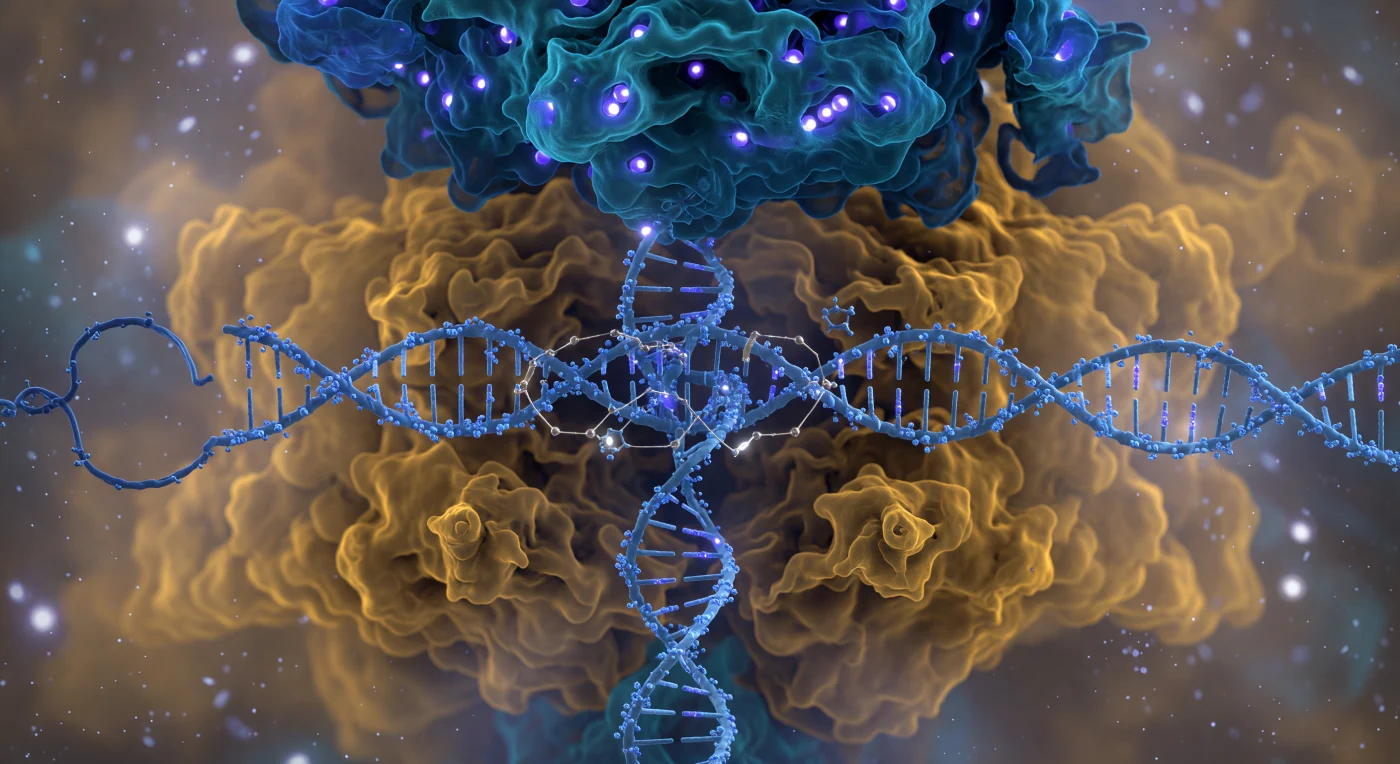
眼前に広がるのは、生きた鉱物から彫り出された大聖堂のような巨大なタンパク質複合体の正面――Cas9の全体像であり、コバルトブルーと深いティールが交錯する認識ローブが頭上に迫り、温かみのある琥珀色と黄土色の核酸分解ローブが眼下に膨らみ、その二葉の間にB型DNAの二重らせんが氷青の撚り綱のように挟み込まれている。らせんのリン酸骨格は冷たい紫色の発光を帯びたノードの連なりとして輝き、その周囲では水分子が微細な衛星球として揺れながら淡い光を四方へ散乱させ、溶媒との境界が固定された輪郭ではなく「交渉によって決まる領域」として常に拡散している。エレクトリックシアンの一本鎖ガイドRNAがローブ間の空間を斜めに横切り、その近傍ではすでに三箇所のRループ架橋が形成されており、RNAと鋳型鎖DNAを引き寄せる水素結合が金白色の繊細な光の糸として視覚と電場感覚の両方に訴えかけてくる。右側の核酸分解ローブではPAM認識ドメインの二本のアルギニン側鎖がNGGトリヌクレオチドのマイナーグルーブへ指のように挿入され、HNH活性部位とRuvC活性部位の深部では二価マグネシウムイオンがグルタミン酸とヒスチジンの酸素配位子に囲まれた正八面体の檻の中心で白熱点として輝いており、DNAを切断するための化学的引力が今まさに臨界点に達しようとしている様子が、全方向から押し寄せる熱的ゆらぎの燐光の霞の中に浮かび上がっている。

あなたは今、秩序を持たないタンパク質の幾何学的中心に浮かんでいる——固体の構造ではなく、可能性の重ね合わせそのものの内側に。直径8ナノメートルほどの拡散した青白い確率雲が四方を包み込み、数十本のポリペプチド鎖コンフォーマーが薄く透き通った状態で互いに重なり合いながら、北極の氷の奥から灯されたような発光する霧を形成している——その輪郭は決して定まることなく、絶えず揺らぎ続ける。これは固有の無秩序タンパク質(IDP)と呼ばれる分子クラスの本質的な姿であり、単一の安定した立体構造を持たず、自由エネルギーランドスケープ上の広大なコンフォメーションアンサンブルとして存在する。左側では琥珀色に淡く輝くアルファヘリックスのリボンが数ナノ秒だけ具現化し——水素結合したカルボニル酸素の螺旋配列がかすかな熱銅線の光を帯びて——次の瞬間には青白い霧の中へと溶けていく。そして周囲の全てを支配するのは水の存在であり、直径0.28ナノメートルの球状水分子がすべての露出した主鎖セグメントへ絶えず衝突し向きを変え続け、その熱運動のエネルギーこそがこのタンパク質の構造的無秩序を駆動する根本的な力として、あなたの全身を均等に叩き続けている。

視線を上へ向けると、温かみのある青銅色と琥珀色の光沢をまとった二重螺旋の塔が、電気的な青みがかった水の靄の中から垂直に立ち上がり、視野の上端へと消えてゆく――A型ヘリックス特有のやや傾いた幾何学的配置が、DNAのそれとは明らかに異なる圧縮された威厳を醸し出している。リン酸基は磨き込まれた黒曜石の突起のように両翼に並び、それぞれが紫青色の静電気のハローをまとって周囲の水分子を微かにイオン化させ、リボースから外へ向かって突き出した2'-ヒドロキシ基は温かな銅色の小さなアンテナとして一定の振動を続けており、これこそがRNAとしての自己証明である。ナトリウムイオンが鋭い銀白色の点となって流れ過ぎ、より重いマグネシウムイオンは金緑色の光の塊としてリン酸骨格の近傍に漂いながら見えない静電気の綱に繋ぎ止められている。はるか頭上、茎の頂点では構造がGNRAテトラループの装飾的な頂塔へと開き、対になっていない塩基が非対称な花冠状に広がって周囲の青白い散乱光を柔らかな金色に屈折させ――この瞬間、ナノメートルの深淵に浮かぶこの高塔全体が、フェムト秒の熱的振動に絶えず揺れながらも、驚くほど完全な構造的秩序を保っている。
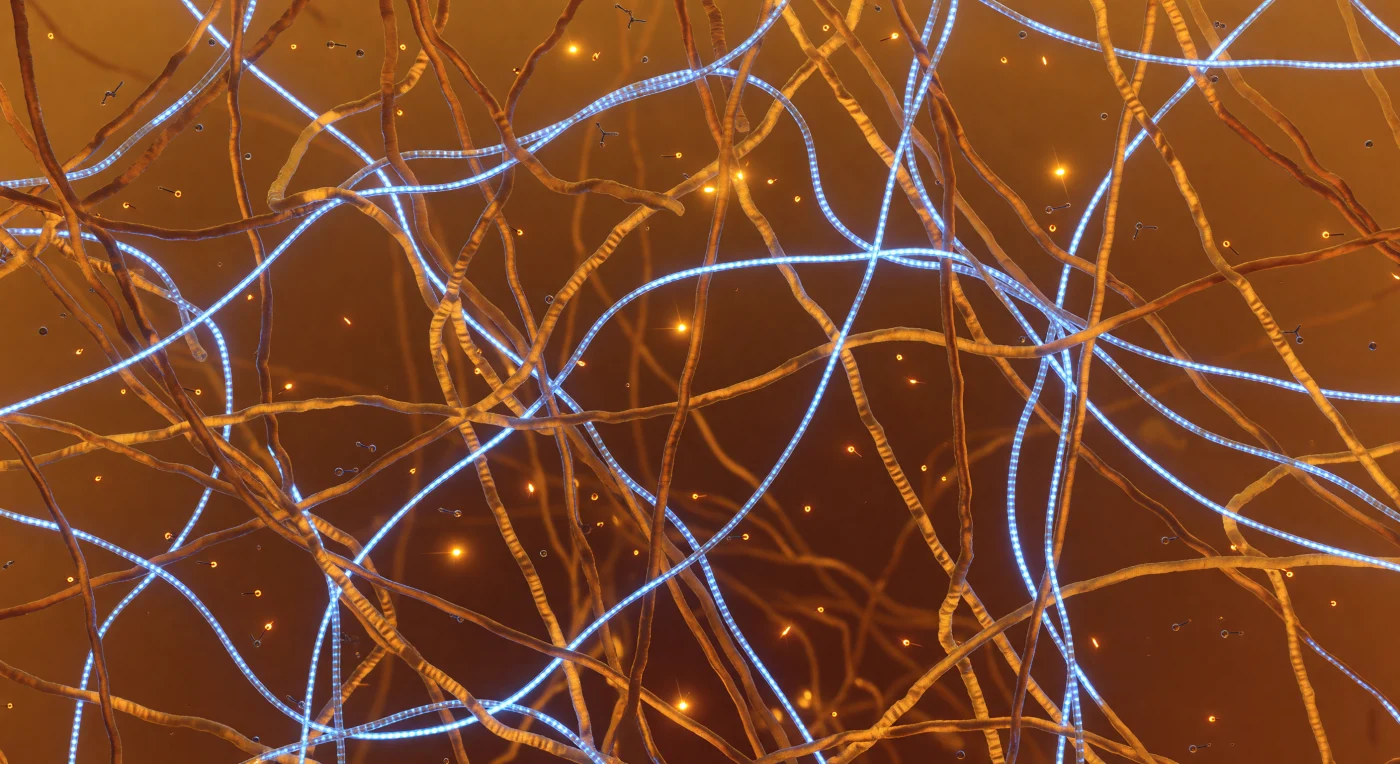
タンパク質とRNAが織りなす凝縮液滴の内部、わずか数ナノメートルの視野の中で、琥珀色に輝く無秩序なポリペプチド鎖が四方八方から迫り、2〜3ナノメートルの太さをもつ鎖どうしが緩やかに絡み合いながら、ほぼすべての空間を埋め尽くしている。これは液−液相分離によって形成された凝縮体の内側であり、低複雑度ドメインと呼ばれる天然変性領域が弱い多価相互作用——チロシンのπ電子系とアルギニンのグアニジニウム基が引き合うカチオン-π接触——を繰り返すことで、固体でも液体でもない半流動的なメッシュ構造を自発的に維持している。その格子の合間を縫うように、直径1.5ナノメートルのRNA鎖が冷たいネオンブルーの光を放ちながら走り、0.34ナノメートル間隔で並ぶ塩基対の段が繊維光学ケーブルの螺旋模様のように浮かび上がっている。5〜15ナノメートルほどの細孔が一瞬だけ開いては閉じ、そこをATP分子がブラウン運動で漂い、燐酸基の尾に静電気的なゆらぎをまとわせながら進路も速度も予測不能に漂流している。5ナノメートルを超えると深度が溶け込み、世界はすべて暖かな琥珀色の霧に包まれて境界を失い、熱エネルギーそのものが空間を淡く発光させているかのような、生きた分子の密林の奥へと消えていく。
