Wetenschappelijke betrouwbaarheid: Hoog

Je bevindt je in het absolute duister van het hydrofobe kern van een gevouwen eiwit, omsloten door de nauw samengedrukte zijketens van leucine, valine en fenylalanine die elkaar raken met een pakkingsdichtheid van 75% — zo dicht dat geen enkel watermolecuul ooit in deze ruimte heeft gepenetreerd. De van der Waals-oppervlakken om je heen zijn zachte, lichtgevende bolvormen in houtskoolgrijs en warm ivoor, hun elektronenwolken gescheiden slechts door de dunste marge van kwantumafstoting, als perfect passende rivierstenen in een geologische grot. Platte, obsidiaanachtige fenylalanineringen snijden de beschikbare ruimte in scherpe alcoven, en waar twee atoomoppervlakken elkaar raken ontstaan harde contactschaduwen — absoluut en onverzacht, want er bestaat hier geen diffuus omgevingslicht. Verspreid door de gepakte matrix glinsteren twee of drie methionine-zwavelatomen als warme, zwavelgele gloed, de enige warmte in een verder achromatische wereld, terwijl ver weg, aan de randen van de kern, een koud blauwgroen schijnsel van het oplosmiddel door de smalle spleten tussen secundaire structuurelementen naar binnen sijpelt als bioluminescentie gefilterd door meters stil oceanwater. De thermische vibraties bij 310 Kelvin — lichaamstemperatuur — laten de gehele structuur trillen in sub-ångström schokjes, een nauwelijks waarneembare moleculaire hartslag die aantoont dat deze geologische stilte niet de rust is van de dood, maar van extreme, levende stabiliteit.

Op de drempel van de peptide-uitgangstunnel trekt de wereld zich samen tot een kathedraal van oker en verbrand brons: de wanden, opgebouwd uit verstrengelende rRNA-helixen waarvan de suiker-fosfaatruggengraten glanzen als gepolijst hoorn, vernauwen zich tot nauwelijks anderhalf nanometer bij de constriktie, waar de van der Waals-oppervlakken van tegenoverliggende rRNA-pilaren elkaar bijna raken in een zachte violet-gekleurde contactzone. Tien nanometer verderop pulseert het peptidyltransferasecentrum als een katalytisch haardvuur van koel blauwwit licht, zijn halo gedempt door lagen interfererende waterdipolen tot concentrische ringen van cyaan en bleek goud, terwijl radiale rRNA-pilaren lange schaduwen werpen die de tunnelwand in afwisselende strepen van indigo en amber verdelen. Recht op de kijker af slingert een nieuwgevormde polypeptideketen in lichtgevend limoengroen en warm amber, haar ruggengraat opvangend in het verre bronlicht langs de geribde kam terwijl de zijketens met vluchtige van der Waals-intimiteit de rRNA-wanden aftasten — kleine hydrofobe knokkels die zich even in de gegroefde oppervlakken drukken voordat de thermische beweging ze weer lospelt. De gehele atmosfeer is er een van geconcentreerde productieve onrust: thermisch geruis, elektrostatische ladingslandschappen en hydrofobe oppervlakken werken samen in een zelfonderhoudend gefluister, terwijl magnesium- en kaliumionen als zilverwittige speculaire punten in de groeven van de rRNA-wanden bezinken, elk omgeven door zijn eigen schaal van georiënteerde watermoleculen — een levende, elektrostatisch geladen doorgang waarin de eerste ogenblikken van een nieuw eiwit worden gesmeed in warm amberlicht en katalytisch blauw vuur.

Je vliegt door een kloof zo smal dat de wanden je van beide kanten insluiten — een helicale corridor van slechts 2,2 nanometer breed, gevormd door het grote groef van een B-DNA-dubbelstreng, waarbij gestapelde aromatische basenparen een mozaïekvloer vormen van warmgoud amberkleurig adenine, saliegroen thymine, diep blauwgroen guanine en hemelsblauwe cytosine, elk platform van het volgende gescheiden door slechts 0,34 nanometer, hun π-elektronenwolken overlappend in een sluierachtige violet-indigo glinstering van kwantuminterferentie. De groefwanden rijzen aan weerszijden op als massieve fosfaat-suikerruggen — oranje-beige en lichtdoorlatend als zeeglas voor een warme lamp — terwijl buiten de geladen rand magnesiumionen als harde, intens wit-blauwe vonken hoveren, omringd door strak georiënteerde watermoleculen, en natriumionen als zachte goudkleurige halo's heen en weer drijven in de elektrostatische nevel. De Debye-afschermlaag geeft de ruimte voorbij de fosfaatrand een melkachtige, elektrostatisch geladen glinstering, een mist van gepolariseerde waterdipolen die golvende fronten van zacht blauw-wit licht door de groef sturen als trage aurora's. Thermische botsingen van alomtegenwoordige watermoleculen vullen de lucht met een aanhoudende trilling, elk contact een vluchtig lichtflitsje dat onmiddellijk wordt opgeslokt door de omringende aquatische gloed, terwijl de helix ver vooruit langzaam naar rechts uitwikkelt en de basenparen in een vervagende reeks van amber, teal, blauw en groen uit het zicht spiraleren.

De kijker bevindt zich op nog geen twee nanometer van een van de meest precieze moleculaire herkenningsprocessen in de biologie — het moment waarop een antilichaam zijn doelwit grijpt. De zes CDR-lussen van het Fab-fragment, elk chemisch uniek van samenstelling, vormen samen een gebogen amphitheater van moleculair reliëf: de amberkleurige H3-lus en de blauwgroene L3-lus strekken zich uit als reusachtige vingers rond een convex epitoop, hun oppervlakken gegraveerd met de volumes van individuele aminozuurzijketens — aromatische ringen als ondiepe schalen, aliphatische ketens als afgeronde richels. Tussen de twee naderende vlakken, over een contactzone van vijf bij zes nanometer, worden watermoleculen naar buiten gedreven als vonken uit een sluitende las, en waar zij verdwijnen ontstaan er cyaanblauwe waterstofbruggen die als lichtgevende draden flakkeren en consolideren — elk een kwantummechanische deling van elektrondichtheid die de bindingsenergie stukje bij beetje vergrendelt. Het oplosmiddel dat dit alles omringt is geen passieve vloeistof maar een thermisch woelend medium van saffierblauwe warmte, dat van alle kanten indringt en elk moleculair oppervlak met een diffuse, oceanische gloed omhult. Wat zich hier voltrekt is geen macroscopische beweging maar een langzame continentale botsing op Ångström-schaal, waarbij twee complementaire oppervlakken van elektrondichtheid elkaar vinden in een omarming die door evolutie over miljarden jaren is verfijnd.
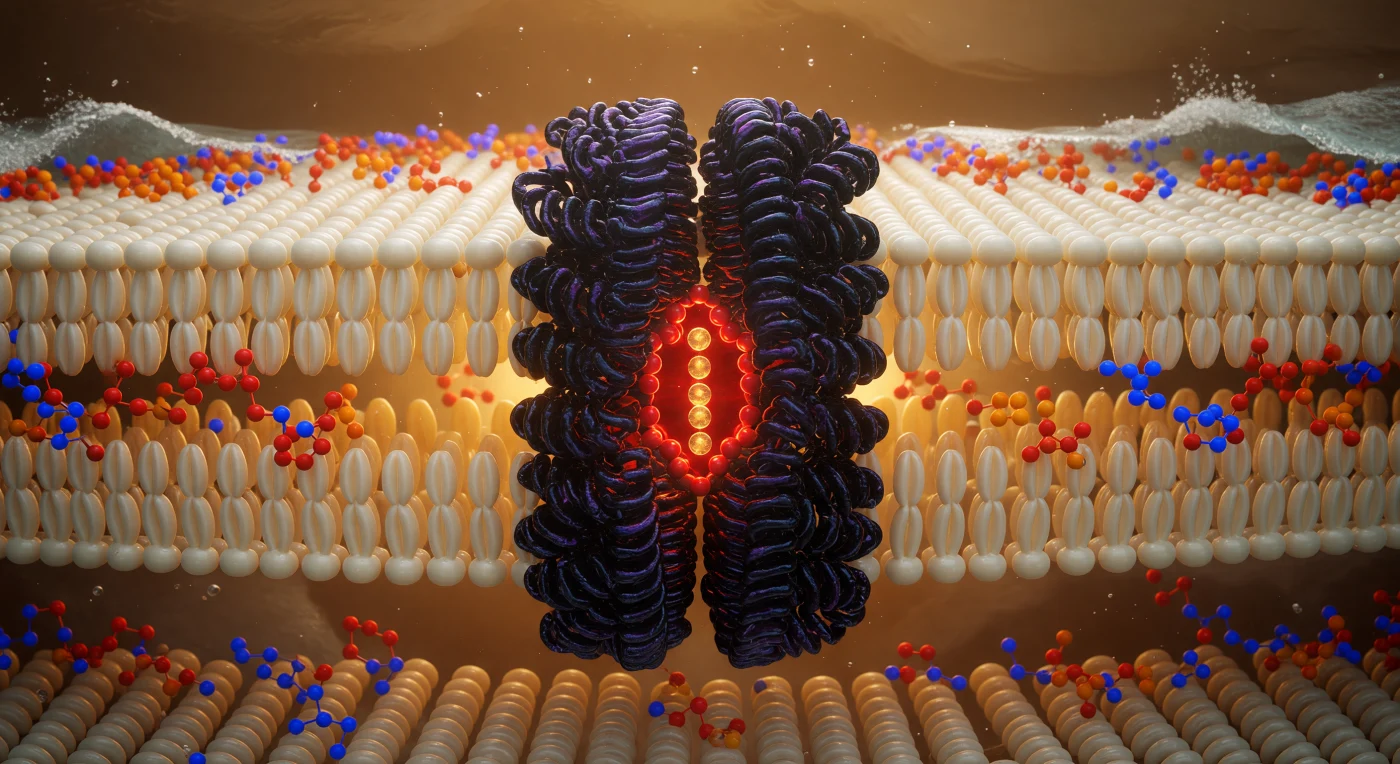
Je bevindt je in het geometrische hart van een fosfolipidendubbellaag, ondergedompeld in het hydrofobe binnenste op slechts twee nanometer afstand van beide kopgroep-kustlijnen, omringd door een dicht, golvend woud van ivoorkleurige en bleekgouden vetzuurketens die zich als een thermische nevel in alle richtingen uitstrekken en waarvan de van-der-Waalsoppervlakken zo dicht op elkaar gepakt zijn dat de tussenliggende ruimte nauwelijks als schaduw zichtbaar is. Het omgevingslicht heeft hier geen enkele bron: het ontstaat uit de flauw elektrostatische gloed van elk elektronenwolk, een diffuus, warm schijnsel dat het gehele hydrofobe interieur doet oplichten als een diepgouden amber-ivoren mist, licht bevend door thermische beweging alsof alles aan de rand van smelten trilt. Dominerend in het gezichtsveld rijst het kaliumkanaal op als een massief basalten monoliet van viervoudige symmetrie — een transmembraankolom van zo'n 3,5 nanometer doorsnede waarvan de diepaarse helices zijn omstrengeld met hydrofobe zijketens die naadloos verstrengeld zijn met de omliggende lipidestaarten, terwijl de selectiviteitsfilter in het poriecentrum brandt met een geconcentreerde rood-oranje gloed van carbonylzuurstofatomen die kaliumionen in een enkelvoudige rij omhullen als smeulende amber kolen. Ver boven en onder je pulseren de glycerofosforycholinekopgroep-kustlijnen met verzadigde atomaire kleur — cadmiumoranje fosforatomen, elektrisch kobaltblauwe stikstofatomen en zachtkarmozen esterzuurstofbruggen — terwijl het water daarbuiten slechts zichtbaar is als een turbulent zilverwitschuim dat tegen het geladen grensvlak drukt als branding tegen een rif.

Je zweeft op ademhoogte boven een uitgestrekte, geribbelde vlakte die zich naar elke horizon uitstrekt — een antiparallel bèta-sheet, de ruggengraatstranden oprijzend als amberkleurige perkamentribben die in afwisselende richtingen lopen met de geometrische regelmaat van oud steenwerk. Cyaanblauwe waterstofbruggen naaien de aangrenzende strengen aan elkaar op slechts 2,9 ångström afstand, als lichtgevende draden van elektrostatische waarschijnlijkheid die de vlakte samenvoegen tot moleculair kettingpantser; kersrode carbonylzuurstofatomen puilen uit elke richel als geslepen granaten, terwijl witte amidetoppen de vloer van elke dal afgrenzen. Dit zelfsamengestelde bouwwerk — gevormd onder thermodynamische druk in een waterig milieu — is niet gefabriceerd maar gegroeid, zijn strenge schoonheid het resultaat van duizenden gelijktijdige zwakke wisselwerkingen die samen een structuur stabiliseren sterker dan de som der delen. Aan de verre rand van de vlakte valt alle orde uiteen in een chaotisch kluwen van rode verbindingslussen, bevroren in schijnbare stilstand maar trillend op nanosecondeschaal, waar het geordende eiwit zich overgeeft aan het omringende waterige blauwgrijze milieu.

Je zweeft in het binnenste van een moleculaire kluis van nauwelijks acht nanometer doorsnede, omsloten door de gladde, grijsivoren wanden van het GroEL-eiwit, waarvan de binnenbekleding van hydrofiele zijketens een koel blauwwit schijnsel verspreidt alsof het materiaal zelf oplicht vanuit het water eromheen. Boven je sluit de GroES-kap de ruimte af als een gewelf van zeven zilverkleurige lobben, strak ineengevlochten als rivierstenen die precies in elkaar passen. In het midden van de kamer drijft op drie meter afstand — op menselijke maat — een verkeerd gevouwen eiwit, een verwarde klomp van amberkleurige lusjes en krommingen die langzaam beweegt onder de onophoudelijke druk van thermische botsingen, zijn oppervlak niet scherp maar waarschijnlijk, als de rand van een zachte vlam. Dit is een van de meest verfijnde moleculaire machines die de evolutie heeft voortgebracht: de chaperonine GroEL-GroES isoleert het misgevouwen eiwit van de cellulaire omgeving, biedt het een beschermde, hydrofiele omgeving aan en dwingt het via ATP-gestuurde conformatieveranderingen tot een nieuwe vouwpoging — een proces dat zich afspeelt in microseconden tot milliseconden, onzichtbaar voor elk menselijk zintuig maar fundamenteel voor het voortbestaan van elke cel.

Je zweeft op vijf nanometer boven een van de meest onwaarschijnlijke machines die de natuur heeft voortgebracht: het replicatievork-complex, een moleculaire fabriek die met verbluffende precisie de volledige genetische blauwdruk van een cel kopieert. Recht onder je torент het CMG-helicase als een monumentale stenen ring van vijftien nanometer doorsnede, opgebouwd uit zes verweven eiwitsubunits waarvan de oppervlakken gepolijst zijn door hydratatieschillen en bezaaid met de ruggen van alfa-helices — het is dit complex dat de inkomende dubbelstrengige DNA-helix, donker als gepolijst jet, onverbiddelijk uit elkaar trekt in twee slingerende enkelstrengige sjablonen, het ene diep tirquoise, het andere goud-amberkleurig, beide golvend in de thermische Browniaanse stroom van het waterige milieu dat hier dikker en weerstandsgebiedend aanvoelt dan lucht ooit zou kunnen zijn. Iets verderop klauwt het DNA-polymerase — een massief grijsocherkleurig moleculair bouwwerk — zich vast aan de leidende streng, en bij elke inbouw van een nieuw nucleotide ontsnapt er een felle oranje-witte fakkel van pyrofosfaat die in een femtoseconde uitwasemt in het aquatische blauw, terwijl magnesiumionen als zilverwitte vonken de actieve kern in en uit schieten langs onzichtbare elektrostatische kanalen. Dit is replicatie op zijn meest elementair: geen passief kopiëren, maar een continue, energieverslindende mechanische prestatie waarbij enkele tientallen eiwitcomplexen in perfecte coördinatie werken om elke milliseconde honderden basenparen foutloos te dubbelen.

Je staat midden in een woud van spiraalvormige eiwitkolommen die zich als gedraaide kathedraalzuilen boven je verheffen — elke alfa-helix amper 1,2 nanometer breed maar met een architectonische precisie die geen menselijk bouwwerk kan evenaren, hun oppervlak gewonden in een strakke rechtsdraaiende groef van warm barnsteen en diep goud waar leucineresten als bronzen spaken naar binnen steken om zich via van der Waals-krachten te verweven met de naburige helixen langs de coiled-coil-naad. Aan de buitenkant steken lysine-zijketens in karmozijnrood en arginine-groepen in elektrisch blauw de waterige omgeving in, hun elektrostatische halo's opgelost in een lichtgevend cerulean medium dat geen lucht is maar een voortdurend trillende massa watermoleculen van 0,28 nanometer die in picosecondes van richting veranderen en vluchtige waterstofbrugnetwerken weven die als gossamer-draden in het licht flikkeren. De corridors tussen de helixparen zijn nauwelijks breed genoeg voor een rij watermoleculen, en langs de hydrofobe contactvlakken heerst een donkerder, drogere schaduw waar water volledig wordt buitengesloten — een zone van dicht, olieachtig contact die zwaar aanvoelt ten opzichte van de heldere solventgangen. Op twintig nanometer afstand verdwijnt dit moleculaire woud in een zacht blauwgrijs waas, de coiled-coil-naden traceend als elegante diagonale lijnen die in een herringbonepatroon oplossen, terwijl de dichtstbijzijnde kolommen in hyperreëel detail fonkelen — elk peptidemotief zichtbaar als afwisselende knooppunten van licht en schaduw langs de gedraaide, fossiel-houten structuur van het eiwitruggengraat.

Je zweeft op de as van een amyloïde fibril, omsloten door vier reusachtige protofilamenten die zich als de vleugels van een kathedraal in perfecte radiale symmetrie om je heen uitstrekken — elke boog opgebouwd uit bèta-strengen die zich in een onverbiddelijk ritme van 4,7 ångström op elkaar stapelen, zo nauwkeurig als de pijpen van een kristallijn orgel. De wanden zijn koud en gepolijst, een diep staalsblauw met iriserende ondertonen waar aromatische elektronenwolken het licht weerkaatsen, terwijl de lichte ribbeling van individuele aminozuurresiduen het oppervlak corrugeerd als in steen gehouwen reliëfs. In het hart van de structuur gloeit een amberkleurige steric zipper — een droge, watervrije binnenste heiligdom waar de zijketens van tegenoverliggende bèta-vellen zo precies in elkaar grijpen als ineinandergestrengelde vingers, bijeengehouden door van-der-Waalskrachten en een bijna geologische stilte. Aan de buitenrand van elk protofilament scheurt dit zwijgen open: glutamaatresiduen steken karmozijnrood naar buiten als vulkanische uitsteeksels geladen met negatieve lading, terwijl lysineresiduen in elektrisch kobaltblauw hun positief geladen toppen uitstrekken in de diëlektrische nevel van gebonden watermoleculen. Dit is wat er overblijft wanneer eens oplosbare eiwitten ineenstorten tot een structuur van beangstigende permanentie — een moleculaire orde die duurzamer is dan steen.
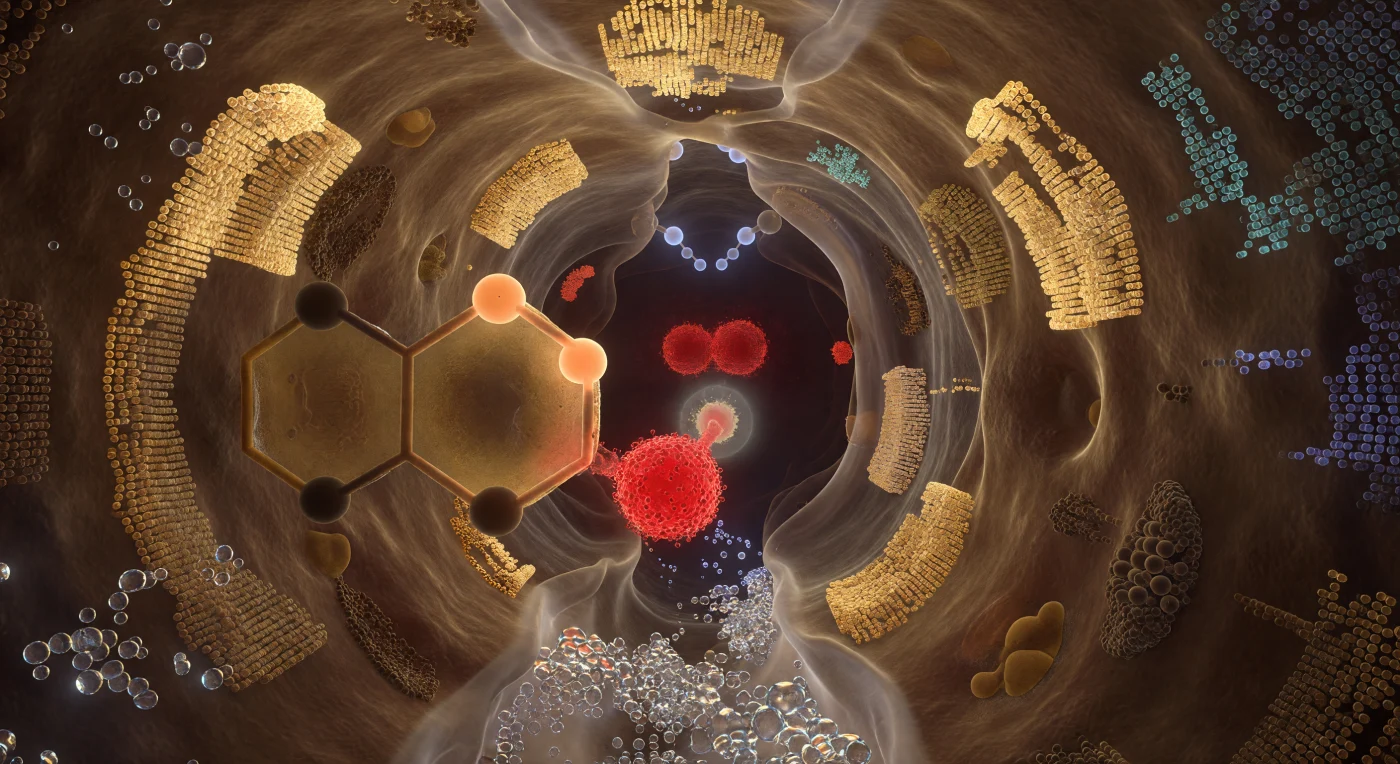
Je staat letterlijk binnenin het werkende hart van een enzym — een holte van nauwelijks twee nanometer breed, omsloten door proteïnewanden die zich als massieve, half-doorschijnende gelaagde rotswanden om je heen welven en een warme, goudachtige gloed uitstralen vanuit hun gepolariseerde elektronenwolken. De katalytische triade domineert de ruimte als drie geologische monumenten: het zuurstofatoom van Ser195 gloeit als een gloeiende kool op slechts 1,5 ångström van de te klieven binding van het substraat, terwijl de imidazoolring van His57 zich als een enorm barnsteen-getint venster aan je linkerflank verheft, een gesommeerde proton middenin zijn overdracht verbergend in apricotkleurig licht, en het carboxylaat van Asp102 diep achter alles als een karmozijnrood anker pulseert met negatieve ladingsdichtheid. Boven het carbonylkoolstof van het substraat vormen twee ruggraat-NH-donoren een soort gewelfde nis — het oxyanion-gat — hun waterstofatomen zichtbaar als kleine, koud-witte knooppunten die electrostatisch licht werpen op de zich vormende tetraëdrische overgangstoestand. Alles hier ademt niet in de tijd maar in kansverdelingen: elke oppervlakte lost aan zijn buitenste ångström op in een kwantummist van elektronenwaarschijnlijkheid, zodat de hele ruimte tegelijk vast en vervaagd aanvoelt, bevroren op het exacte moment waarop chemie werkelijkheid wordt.

Je zweeft op drie nanometer van de collageenkabel, zo dichtbij dat haar oppervlak je volledig gezichtsveld vult van links naar rechts: drie polypeptideketens — warm ivoor, bleek goud en gebleekt zand — zijn ineengevlochten tot een trage, majestueuze rechtshandige superheli van amper 1,5 nanometer breed, als een touw gevlochten over geologische tijd. Op elke derde positie langs elke keten markeren de stijve pyrrolidineringen van prolineresiduen de structuur als grijsbleke knokkels, hun vlakke cyclische geometrie licht oplopend in het diffuse, richtingloze licht dat voortkomt uit de thermische stralingsbad van de moleculaire omgeving zelf, terwijl de amberkleurige hydroxylgroepen van hydroxyproline naar buiten steken als kleine geladen dauwdruppels. De kabel omhult zichzelf in een geestachtig bleekblauwe eerste hydratatieschil — een vrijwel perfecte tetraëdrische rangschikking van watermoleculen op 2,8 ångström afstand van elkaar, vergrendeld in een waterstofbrugrooster dat de triple heli als bevroren glas inkapselt en zacht blauw-wit polarisatielicht weerkaatst. Voorbij die geordende schil lost de structuur op in een warme, turbulente moleculaire nevel van aquamarijnen en zilverwittig licht, het collectieve adem van tienduizenden watermoleculen per kubieke nanometer, waartussen de kabel zich uitstrekt over honderden nanometers als een eindeloze corridor die verdwijnt in lichtgevende verte.
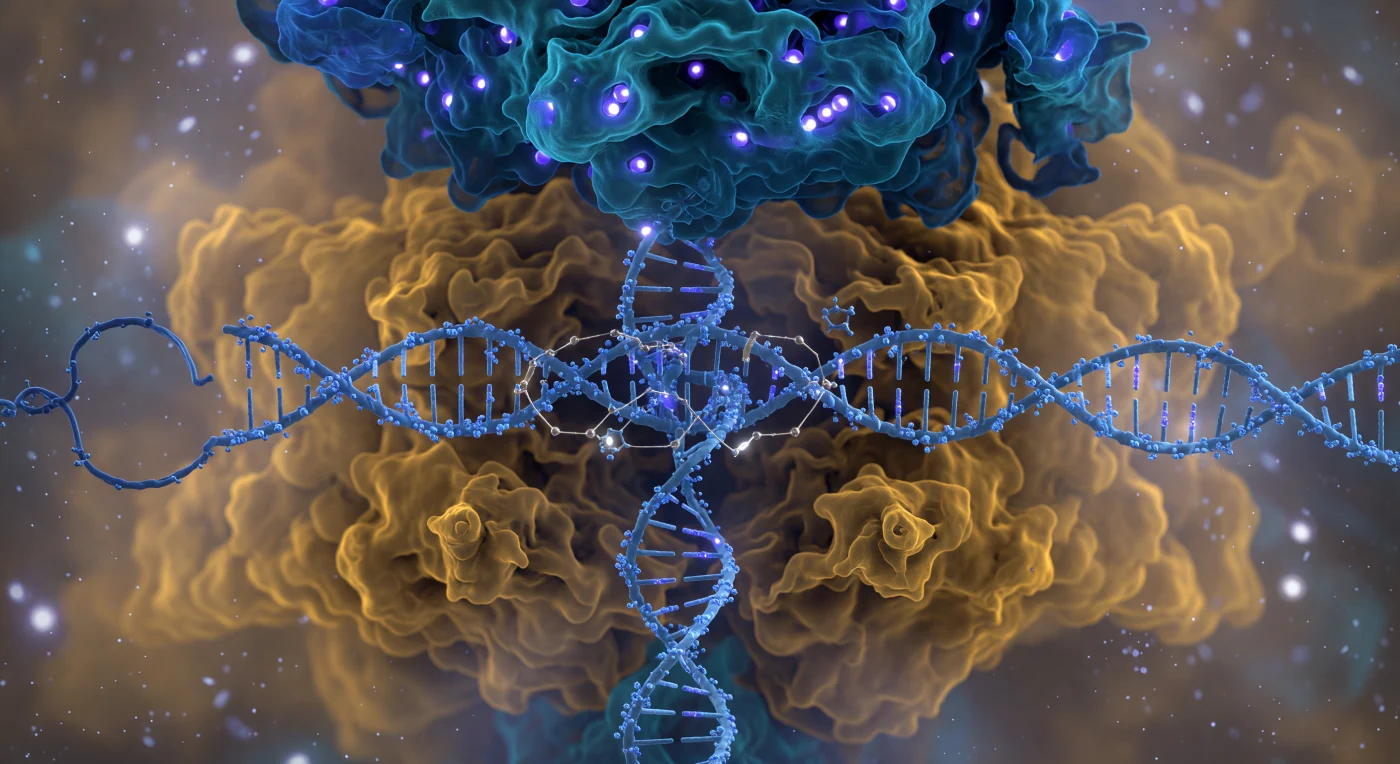
Vijf nanometer van het Cas9-complex zweef je voor een klifrots van gevouwen eiwit die je volledig gezichtsveld vult — een bilobaire reus van negen nanometer breed, waarvan de kobaltblauwe herkenningslob zich als een gewelf boven je verheft en de warme amber nucleaselob naar buiten bolt met oppervlakken die nooit scherp zijn, omdat de grens tussen molecuul en oplosmiddel eerder onderhandeld dan vastgesteld is. Door de kloof tussen de twee lobben snijdt een ijsblauwe B-vorm DNA-dubbelhelix als een gedraaid touw van bevroren fosfaatladingen, terwijl de elektrisch cyaan gekleurde gids-RNA-spacer er doorheen weeft met drie zichtbare R-lus-bruggen — elk een plek waar waterstofbindingen als gouden draden twee complementaire basen naar elkaar toe trekken in een geometrie die je eerder voelt dan ziet. Aan de rechterzijde drukt het PAM-interagerende domein zijn argininevingers als kopergloeiende spatelarmen in de kleine groef van het NGG-trinucleotide, waardoor de helixgeometrie plaatselijk subtiel vervormt, terwijl diep in de actieve kern twee Mg²⁺-ionen als witgloeiende puntlichten flikkeren, omringd door coördinerende zuurstofatomen in een perfect octaëdrische geometrie. Het omringende oplosmiddel is geen leegte maar een lichtgevende thermische oceaan van waterdipolen en tegenionen die alles in constante nanoschaalturbulentie baadt — een herinnering dat op deze schaal elke grens, elke binding, elke structuur slechts een tijdelijk evenwicht is in een wereld van onophoudelijke beweging.

Je bevindt je in het kloppende hart van een intrinsiek wanordelijk eiwit, omgeven door tientallen spookachtige polypeptideconformaties die gelijktijdig dezelfde ruimte bewonen — elk bijna transparant, maar samen opbouwend tot een diffuse blauwwitte nevel van ongeveer acht nanometer breed, met ijle uitlopers die tot vijftien nanometer reiken als cirruswolken die in het oplosmiddel vervagen. Dit eiwit heeft geen vaste structuur: het bestaat als een statistisch ensemble van mogelijkheden, een kansarchitectuur waarbij elke conformatie even reëel is als de volgende, gedreven door thermische energie en voortdurend geherstructureerd door de omringende waterlaag. Links van je materialiseert een amber glanzend alfa-helix — zijn waterstofgebonden carbonylzuurstofatomen oplichten als gloeiend koper — maar binnen nanoseconden lost het alweer op in de koele parelwitte wazigheid van het ensemble, terwijl een vluchtige cluster van aromatische residuen zoals fenylalanine en tryptofaan kortstondig goudgeel pulseert als gloeiende as. Het water is alomtegenwoordig en dominant: bolronde watermoleculen van nauwelijks 0,28 nanometer bombarderen elk blootgesteld ruggraatsegment in voortdurende thermische agitatie, hun georiënteerde dipolaire oppervlakken licht blauw tintelend, en het is juist deze agressieve solvatatie die elke voorbijgaande structuur vormt, destabiliseert en opnieuw ontketent.

Vanuit dit standpunt, diep onder de voet van de RNA-haarpeld, rijst de dubbele helix op als een bronzen kathedraal die zich door het elektrisch blauwe waterige waas heen boort — haar A-vorm geometrie merkbaar compacter en schuiner dan DNA, elke ribose-fosfaateenheid gehuld in warm barnsteenkleurig koper waarvan de elektronendichtheidsranden oplossen in een zwak elektrostatisch schijnsel. De 2'-hydroxylgroepen steken als kleine koperen antennes uit elke ribose naar buiten, wat RNA onmiskenbaar onderscheidt van haar genetische tegenhanger, terwijl de fosfaatgroepen langs beide zijden van de ruggengraat als rijen gepolijste obsidiaanknopen fonkelen, elk omgeven door een krakende halo van negatieve lading die het omringende water doet oplichten in violet-blauwe coronae. Natriumionen scheuren als harde zilverwitte punten door het medium met vluchtige hydratatieschillen in hun kielzog, terwijl zwaardere magnesiumionen dicht bij de ruggengraat zweven in goudgroene clusters, vastgehouden door onzichtbare elektrostatische banden aan het fosfaatskelet. Hoog boven, waar de steel overgaat in de GNRA-tetralus, opent de architectuur zich in een barokke torenspits van ongepaard gebleven nucleotiden waarvan de geconserveerde adeninebasen gestapeld en kruislings verstevigd zijn tot een verrassend rigide platform dat goudkleurig oplicht tegen het koelere blauw. De gehele kolom trilt onzichtbaar op femtosecondeschaal, en toch legt dit bevroren moment een onvergelijkbare structurele volmaaktheid vast — monumentaal, oud, en levend tot in elke elektronenwolk.
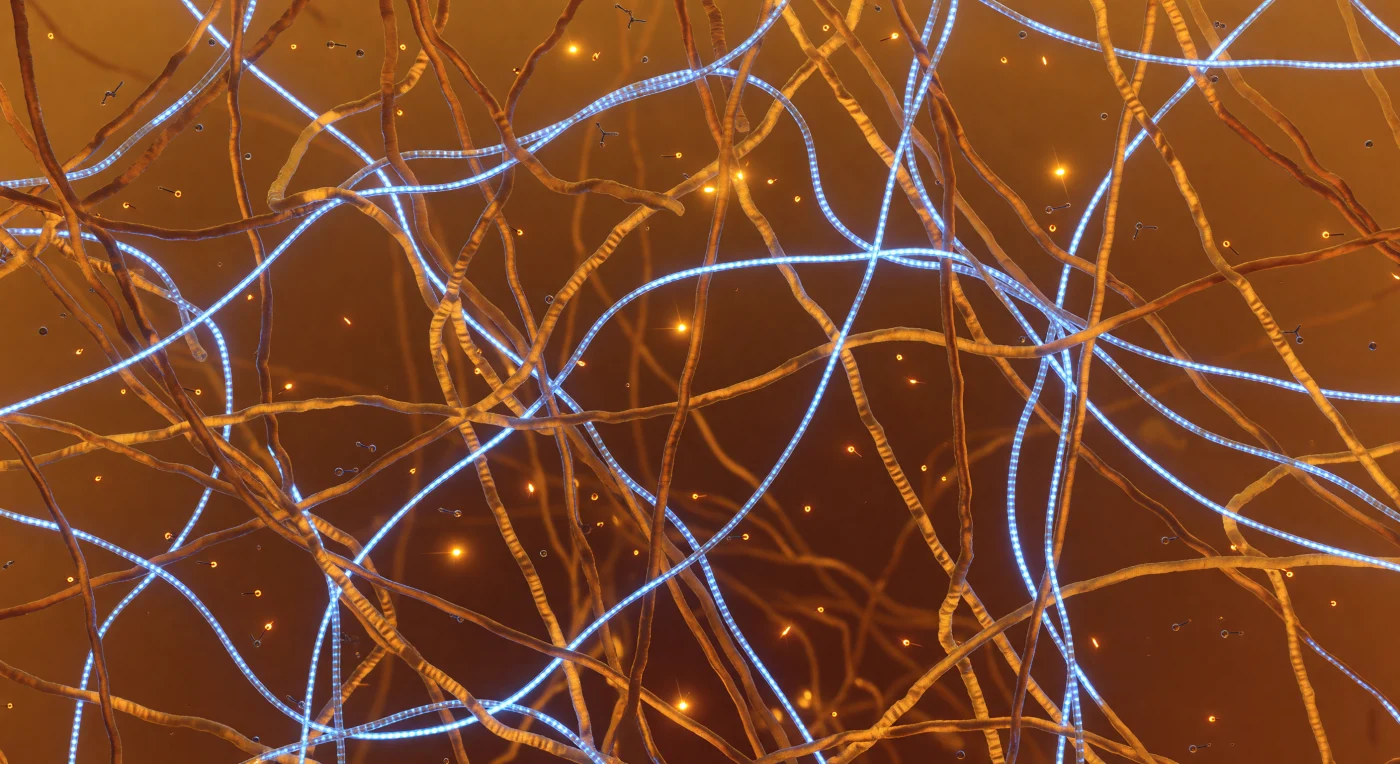
De wereld om je heen is één doorschijnend amberkleurig weefsel — een ondoordringbaar dicht netwerk van gedesorganiseerde eiwitketens dat zich in alle richtingen uitstrekt, elke twee tot drie nanometer gevuld met zachte, matte polymeerdraden die gloeien als oud hars voor een kaarsenvlam. Dit is het binnenste van een fase-gesepareerd condensaat, een biomoleculaire druppel die spontaan is ontstaan doordat eiwitten met lage-complexiteitsgebieden en RNA-strengen via zwakke, veelvuldige interacties een vloeibaar-achtige maar dense afzonderlijke fase vormen — een proces aangedreven door cation-π-contacten tussen tyrosine en arginineresiduen, die hier en daar als kortstondige oranje vonken oplichten en binnen fracties van een nanoseconde alweer doven. Door dit amber labyrint slingeren RNA-strengen van precies 1,5 nanometer breed als blauw-witte lichtkabels, hun fosforugruggengraat en gestapelde aromatische basen periodiek opgelicht met een koele neongloed die bleekblauwe reflecties werpt op de naburige eiwitwanden en kleine tealachtige halo's achterlaat waar nucleïnezuur en polypeptide elkaar raken. In de donkerdere holten van vijf tot vijftien nanometer — kortstondige openingen in het maaswerk — drijven ATP-moleculen in puur Browniaanse beweging, hun trifosfaatstaarten omgeven door een nauwelijks zichtbare elektrostatische trilling. Vijf nanometer verder lost alles op in een warme, gelijkmatige amberkleurige nevel: geen horizon, geen harde grenzen, alleen de zachte, zelfgegenereerde gloed van thermische energie die door dit levende, ademende moleculaire interieur pulseert.
