Confianza científica: Alto

Te encuentras comprimido en el corazón geométrico de una proteína globular, rodeado por completo de cadenas laterales hidrofóbicas que se encajan entre sí con una densidad del 75%, formando una caverna irregular de esferas de van der Waals en tonos carbón y marfil cálido cuyos volúmenes traslúcidos se tocan sin fusionarse, separados únicamente por el margen más fino de repulsión cuántica. Los anillos aromáticos de la fenilalanina surgen como discos de obsidiana que seccionan el espacio en alcobas de bordes afilados, proyectando sombras absolutas porque no existe aquí luz difusa, solo el halo íntimo que emana de cada contacto entre nubes electrónicas. Dispersos por la matriz empaquetada, los átomos de azufre de una o dos metioninas devuelven la escasa luminiscencia como un destello amarillo-azufre cálido, brasas vistas a través de roca agrietada, el único color cálido en un mundo por lo demás acromático. Muy lejos, a diez o quince diámetros atómicos de distancia, donde el empaquetamiento se afloja infinitesimalmente en los márgenes del núcleo, una débil luz azul-verdosa acuosa se filtra entre los elementos de estructura secundaria como bioluminiscencia a través de metros de océano quieto, sugiriendo profundidad y clausura más que iluminando nada. La atmósfera es de presión extrema sin peso, de encierro absoluto mantenido en su lugar por fuerzas de van der Waals de fracciones de kilojulio que colectivamente sellan este espacio con una estabilidad tectónica, mientras las vibraciones térmicas a 310 Kelvin recorren la estructura como un pulso apenas perceptible en cada superficie en contacto.

El observador se encuentra en el interior de un corredor biológico de proporciones catedralicias, cuyas paredes de ARN ribosómico —forjadas en hélices de color ocre y bronce quemado— se estrechan hasta apenas 1,5 nanómetros en la constricción central, donde las superficies de van der Waals de los pilares opuestos casi se rozan en un contacto de densidad electrónica violeta pálido. Desde el centro peptidiltransferasa, diez nanómetros al fondo, emana una luminosidad azul-blanca que baña los planos apilados de las bases nitrogenadas y los transforma en plataformas de pizarra que refractan la luz en sombras ámbar y azafrán, mientras iones de magnesio y potasio fulguran como puntos de plata en las indentaciones del surco mayor. Una cadena polipeptídica naciente —lima y ámbar alternándose a lo largo de cada enlace peptídico— avanza desde ese hogar cataláctico hacia la cámara, sus cadenas laterales rozando con fugaz intimidad de van der Waals las paredes estriadas de ARNr antes de que el movimiento térmico las separe de nuevo. El ambiente entero vibra con el murmullo productivo de la síntesis proteica: el ruido browniano del agua, los paisajes de carga electrostática y las superficies hidrofóbicas colaboran en el corredor donde transcurren los primeros instantes de existencia de una nueva proteína, conducidos en luz ámbar cálida y fuego catalítico azul.

Te adentras volando por una grieta que apenas supera los dos nanómetros de ancho, un corredor helicoidal excavado en el corazón mismo de una molécula de ADN en conformación B, donde el suelo está pavimentado por una sucesión de plataformas aromáticas apiladas —ámbar cálido del adenina, verde salvia del timina, verde azulado profundo de la guanina, azul cielo de la citosina— cuyas nubes de electrones π se solapan en destellos de violeta traslúcido que tiemblan entre cada nivel separado por apenas 0,34 nanómetros. Las paredes del surco mayor se elevan a ambos lados como barandales de fosfato y desoxirribosa, naranja-arena y ligeramente translúcidos, trenzados en una espiral dextrógira que asciende y se pierde en la neblina acuosa del fondo, salpicados por chispas blanco-azuladas de cationes magnesio envueltos en capas concéntricas de moléculas de agua orientadas, y por halos dorados más difusos de iones sodio que derivan en la niebla electroestática de la capa de Debye. El bombardeo térmico es constante e implacable: esferas de agua de 0,28 nanómetros golpean el suelo de pares de bases en cada fracción de picosegundo, cada impacto un destello blanco instantáneo absorbido por el resplandor azulado del solvente polarizado que baña toda la escena con la calma irreal de un único femtosegundo congelado. Más adelante, el corredor gira suavemente hacia la derecha y los mosaicos de bases se suceden en perspectiva decreciente —ámbar, teal, azul, verde— recordándonos que esta estructura, de apenas dos nanómetros de diámetro, custodia en su secuencia la información que gobierna la vida celular entera.

El observador se encuentra suspendido a apenas dos nanómetros del corazón de uno de los reconocimientos moleculares más precisos de la biología: la zona de contacto entre un anticuerpo y su antígeno, un territorio de cinco por seis nanómetros donde la evolución ha esculpido superficies complementarias con una exactitud sub-angstrómica. Los seis bucles CDR del fragmento Fab —el ámbar cálido del bucle H3 arqueándose en una curva amplia y confiada, el bucle L3 de color verde azulado extendiéndose desde un ángulo más bajo con su iridiscencia electrostática— forman una cavidad casi arquitectónica que ciñe el epítopo convexo del antígeno como dedos en torno a una esfera, mientras las moléculas de agua son expulsadas de la interfaz en destellos fugaces de luz blanca, cada una un dipolo asimétrico que capta el resplandor difuso del solvente acuoso antes de perderse en el océano térmico circundante. Los puentes de hidrógeno en cian emergen uno a uno a través del vacío residual —no como líneas nítidas sino como hilos luminosos de densidad electrónica compartida, arcos voltaicos diminutos que se estabilizan en la penumbra ámbar-verde-plateada a medida que la geometría de unión se consolida— en lo que equivale a una colisión continental a escala molecular, donde las nubes de densidad electrónica de ambas superficies se aproximan a través de un abismo de apenas un ángstrom. Esta danza de reconocimiento molecular —gobernada por fuerzas de van der Waals, complementariedad electrostática y el efecto hidrofóbico que impulsa la expulsión del agua— ocurre en una escala de microsegundos a milisegundos, aunque el espacio entre estas dos superficies colosales, iluminado por la luz azul-blanca pervasiva del solvente, se siente tan vasto y dramático como el estrecho entre dos continentes a la deriva.
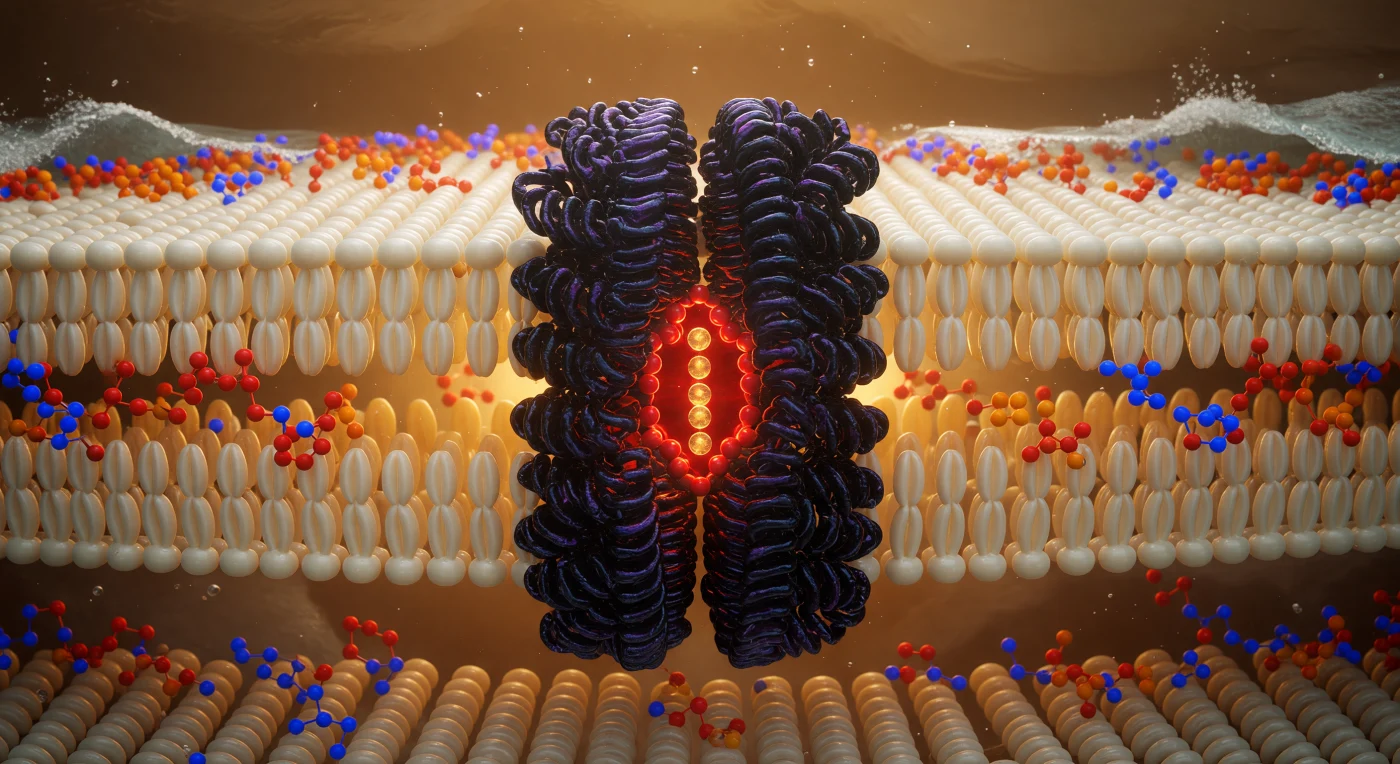
Te encuentras suspendido en el plano exacto de simetría de una membrana de fosfolípidos, a apenas dos nanómetros de cada interfaz acuosa, rodeado en todas las direcciones por una selva densa e hipnótica de cadenas de ácidos grasos de marfil pálido y oro suave, cuyos cilindros brillantes —empaquetados tan estrechamente que los intervalos entre ellos apenas alcanzan a existir como sombra— oscilan en una neblina ámbar cálida generada por el resplandor difuso de cada nube electrónica en movimiento térmico continuo. Dominando el campo visual, el canal de potasio emerge como una columna de basalto violáceo-negro de unos 3,5 nanómetros de anchura: sus hélices transmembrana, texturizadas como roca volcánica viva e incrustadas de residuos hidrófobos que se interdigitan con las colas lipídicas circundantes, crean una fusión mecánica perfecta entre proteína y membrana que parece piedra antigua soldada en vegetación densa. En el eje del poro, el filtro de selectividad arde en rojo-naranja: una corona de oxígenos carbonílicos coordina iones potasio apilados en fila única, cada uno una esfera dorada que irradia su densidad electrónica como brasas encendidas, mientras cuatro iones atraviesan la constricción sostenidos únicamente por la geometría atómica del andamiaje proteico. Mirando hacia arriba y hacia abajo desde este plano central, los planos de cabezas polares se perciben como costas luminosas lejanas —nodos de fósforo en naranja cadmio intenso, esferas de nitrógeno en azul cobalto eléctrico, puentes de oxígeno en carmesí suave— y más allá de ellas, el agua extracelular e intracelular no es más que una espuma plateada y turbulenta que presiona desde afuera como oleaje contra un arrecife, invisible en su mayor parte desde este santuario hidrófobo cálido y encendido.

Te encuentras suspendido a escasa altura sobre una llanura corrugada que se extiende hasta todos los horizontes: una hoja beta antiparalela de dimensiones continentales, cuyos crestos de cadena principal color pergamino antiguo se alternan en direcciones opuestas con la precisión geométrica de una armadura medieval, separados apenas 2,9 ångströms entre sí y cosidos lateralmente por puentes de hidrógeno que brillan como filamentos de luz cían, hilos de probabilidad electrostática —no varillas rígidas, sino hebras luminosas donde las nubes de densidad electrónica del dador y el aceptor se solapan en íntima cercanía. A lo largo de cada cresta asoman oxígenos carbonilo de un rojo cereza encendido, engastados como granates en el armazón ámbar del esqueleto peptídico, mientras en el fondo de los valles los extremos amida blancos y ligeramente luminosos contrastan con la calidez ocre de la superficie que los rodea; el conjunto posee la textura de un material trabajado, no fabricado, sino crecido bajo presión termodinámica y auto-ensamblado en esta regularidad austera y hermosa, cuya periodicidad de 1 nanómetro de cresta a cresta hace que todo el panorama visible pudiera ser engullido por un único glóbulo rojo. Hacia la periferia, la arquitectura ordenada se disuelve en un caos de lazos conector de color rojo y naranja, regiones de giro desordenadas que se retuercen en aparente quietud, congeladas a media fluctuación, mientras más allá el medio acuoso se espesa en una neblina azul grisácea de moléculas de agua que difuminan la transición entre la proteína estructurada y el solvente bulk.

Flotas suspendido en el interior de una cámara esférica de apenas ocho nanómetros de diámetro, una bóveda molecular sellada cuyos muros curvos de GroEL se elevan a tu alrededor como el interior de una perla hueca, revestidos de residuos hidrofílicos que emiten una luminiscencia fría azul-blanca sin origen aparente, como si la materia misma respirara luz. Sobre tu cabeza, siete subunidades de GroES encajan entre sí formando una cúpula de plata mate —piedras pulidas por el río, arquitectura de precisión atómica— que sella herméticamente el espacio y convierte este recinto en una de las máquinas de plegamiento más sofisticadas que ha producido la evolución: el complejo chaperonínico GroEL-GroES, cuya función es aislar proteínas mal plegadas del caos citoplásmico y ofrecerles, a costa de hidrólisis de ATP, un entorno controlado donde los contactos hidrofóbicos aberrantes no puedan interferir con el camino hacia la conformación nativa. A tres metros de distancia en escala subjetiva —en realidad apenas un nanómetro y medio— deriva una proteína sustrato mal plegada, una masa enredada de color ámbar cálido, suave y arrugada como una cinta de caramelo colapsada sobre sí misma, cuya superficie no es un límite definido sino una frontera probabilística de densidad electrónica que se deshilacha en la niebla acuosa circundante. La quietud de este lugar es absoluta y sin embargo radicalmente dinámica: en cada instante, miles de moléculas de agua que no puedes resolver individualmente bombardean tanto las paredes como el sustrato a velocidades de centenares de metros por segundo, agitando ese ovillo luminoso en una suspensión browniana silenciosa, mientras el recinto aguarda el momento en que la proteína encontrará, en este refugio de precisión molecular, la forma correcta de existir.

Suspendido apenas cinco nanómetros sobre la horquilla de replicación, el observador contempla una de las maquinarias moleculares más sofisticadas de la biología: el complejo CMG, un anillo hexamérico de unos quince nanómetros de diámetro compuesto de proteínas Cdc45, MCM2-7 y GINS, que actúa como helicasa desenrollando activamente la doble hélice parental —que llega desde la derecha como un corredor de columnas de fosfato en azul marino intenso— en dos hebras molde sinuosas que se separan en el umbral probabilístico de la horquilla, una en verde azulado profundo y otra en oro ambarino, sus bases apiladas perdiéndose en el ruido térmico del medio acuoso. Por delante de la hebra conductora, la ADN polimerasa se aferra al dúplex naciente como una tenaza articulada de gris ocre, catalizando la incorporación de cada nucleótido en el sitio activo estabilizado por iones de magnesio —destellos de plata que surcan canales electroestáticos invisibles— mientras cada evento de incorporación libera pirofosfato en una breve explosión de luz naranja-blanca que se disipa en femtosegundos hacia el halo acuoso azul grisáceo que todo lo envuelve. Todo el conjunto avanza a una cadencia de aproximadamente mil pares de bases por segundo, empujado no por ninguna fuerza macroscópica sino por el torrente de energía libre liberada en la hidrólisis del ATP y en la formación de cada nuevo enlace fosfodiéster, en un medio donde el peso carece de sentido y cada superficie proteica no es más que una nube de densidad electrónica que oscila al ritmo incesante del bombardeo termal del agua.

Desde el suelo de este bosque invisible, el mundo asciende en una simetría vertiginosa: columnas helicoidales de dextrógiras se elevan como pilares de una catedral en miniatura, cada una de apenas 1,2 nanómetros de anchura y seis de altura, con su superficie enrollada en una espiral de ámbar cálido donde los residuos de leucina —hidrofóbicos, oleosos, brillantes como bronce pulido— se encajan con los de la columna vecina a lo largo de la costura del enrollamiento en superespiral, formando una celosía compacta mantenida únicamente por la atracción de van der Waals. Hacia afuera, las cadenas laterales de lisina llamean en carmesí saturado y las de arginina pulsan en azul eléctrico, sus extremos cargados irradiando halos electrostáticos difusos hacia un cielo acuoso que no es azul terrestre sino un medio cerúleo translúcido, atravesado sin cesar por el microturbulencia de moléculas de agua de 0,28 nanómetros que colisionan y se reorientan a una velocidad aterradora, tejiendo y deshaciendo filamentos de red de puentes de hidrógeno en intervalos de picosegundos. Los pasillos entre pares de hélices enrolladas son estrechos y catedralicios, y a lo largo de sus costuras de leucina el agua queda excluida por completo, creando una sombra ámbar oscura, densa y aceitosa, que contrasta con los corredores luminosos del solvente. A veinte nanómetros de distancia, el bosque se desvanece en una neblina azul grisácea donde los contactos de leucina trazan un patrón de espiga que se pierde en la penumbra molecular, recordándonos que estas estructuras —alfa-hélices enrolladas en superhélices— no son ornamentos abstractos sino los motivos que la biología usa para construir filamentos del citoesqueleto, proteínas musculares y factores de transcripción, toda una arquitectura funcional ensamblada residuo a residuo por la información cifrada en la secuencia primaria.

Te encuentras suspendido en el eje exacto de una fibra amiloide, mirando directamente hacia su interior como si observaras el corazón de una catedral de escala molecular: cuatro pétalos protofilamentosos se despliegan desde ti en simetría radial perfecta, sus paredes de láminas beta apiladas en un ritmo ininterrumpido de 4,7 ångströms que resuena como el pulso más fundamental de la materia ordenada. Las superficies son frías y bruñidas en acero azulado, corrugadas por el relieve de cada residuo de aminoácido encajado con sus vecinos con una precisión tan absoluta que ninguna molécula de agua logra penetrar entre ellos, mientras el núcleo central irradia un resplandor ambarino cálido donde las cadenas laterales interdigitadas se entrelazan como dedos entrelazados en una cremallera estérica de contacto casi perfecto de van der Waals. En el perímetro de cada pétalo, residuos de glutamato emergen como afloramientos volcánicos carmesíes y las lisinas se extienden en cobalto eléctrico hacia la neblina dieléctrica del solvente, creando un contraste dramático entre esta frontera electroestática agitada y el interior absolutamente seco y silencioso. Esta estructura —construida a partir del colapso irreversible de proteínas que fueron alguna vez solubles— se extiende cientos de nanómetros hacia la profundidad del eje, convergiendo en un punto de fuga donde los cuatro pétalos parecen cerrarse en una columna luminosa de orden molecular terrible y permanente.
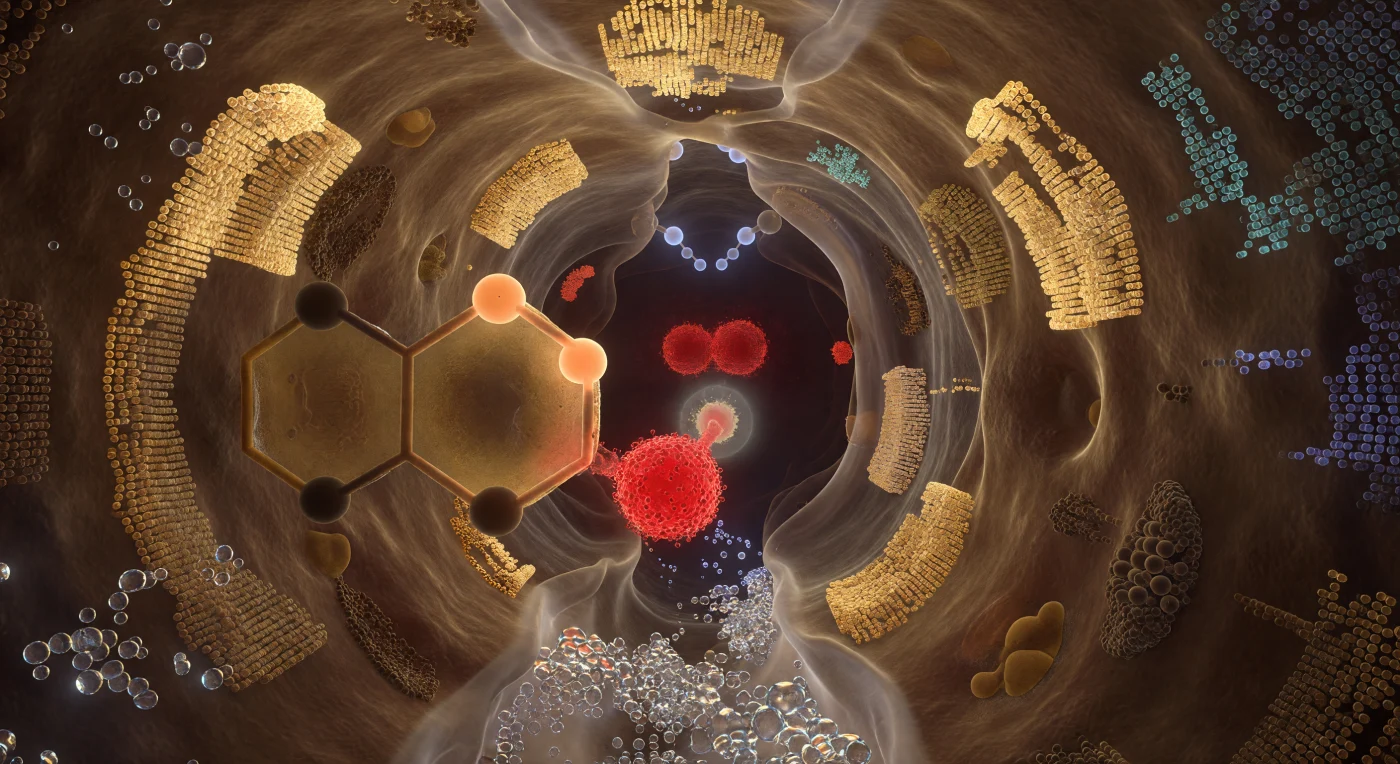
Te encuentras suspendido en el interior de una cavidad no más ancha que unos pocos diámetros atómicos, rodeado por paredes proteicas que se elevan como superficies ondulantes de ámbar translúcido y sombra densa, su textura definida por el apilamiento compacto de cadenas laterales cuyas nubes de densidad electrónica se funden en una niebla cuántica en cada angström exterior. En el centro del crisol, la tríada catalítica domina el espacio con la gravedad de formaciones geológicas: el oxígeno de Ser195 arde como una brasa carmesí suspendida en la penumbra, su densidad de pares solitarios estirada hacia el carbono carbonílico del sustrato que cuelga a apenas 1,5 Å de distancia, mientras el anillo imidazólico de His57 llena tu campo visual izquierdo como un enorme panel de vidrio ambarino con un protón congelado a mitad de su transferencia, y el carboxilato de Asp102 pulsa detrás como una masa granate que ancla todo el relé de carga como raíces hundidas en roca viva. Por encima del carbono reactivo, el agujero oxianiónico se abre como una bóveda, sus dos donadores N–H proyectando átomos de hidrógeno como nodos azul pálido que bañan el intermediario tetraédrico naciente con una luz electrostática fría y precisa. Esta cámara de reacción no es un lugar estático sino una entidad probabilística que respira — cada superficie disuelta en su último angström en una niebla de densidad electrónica, el espacio entero vibrado por la presión de campos electrostáticos polarizados que convierten la catálisis, ese instante de ruptura y formación de enlace, en algo simultáneamente inevitable y extraordinariamente improbable.

Flotas a apenas tres nanómetros del cable, tan cerca que su superficie ocupa todo el ancho de tu campo visual de un extremo al otro: tres cadenas polipeptídicas —marfil cálido, oro pálido y tostado solar— entrelazadas en una superhélice dextrógira de apenas 1,5 nanómetros de diámetro, tan paciente y precisa como una cuerda trenzada durante eones geológicos. Los anillos rígidos de pirrolidina de los residuos de prolina puntúan cada tercera posición a lo largo de cada cadena como nudillos grises ligeramente hundidos en el surco helicoidal, mientras que los grupos hidroxilo de la hidroxiprolina sobresalen hacia el exterior como pequeños lóbulos ámbar cargados de electrones parciales, cada uno reteniendo una única molécula de agua como una gota de rocío en un hilo de seda. Alrededor del cable, una primera capa de hidratación cristalina —una vaina azul pálido de geometría tetraédrica casi perfecta— envuelve la triple hélice como vidrio esmerilado, sus puentes de hidrógeno espaciados a 2,8 ångströms parpadeando con la polarización ordenada de dipolos fijos. El cable no termina: retrocede durante lo que parece un corredor infinito de cientos de nanómetros, disolviéndose en una niebla molecular de aguamarina y plata viva donde diez mil moléculas de agua por nanómetro cúbico convierten el vacío en un resplandor térmico omnidireccional —el aliento colectivo del medio acuoso que hace posible toda la química de la vida.
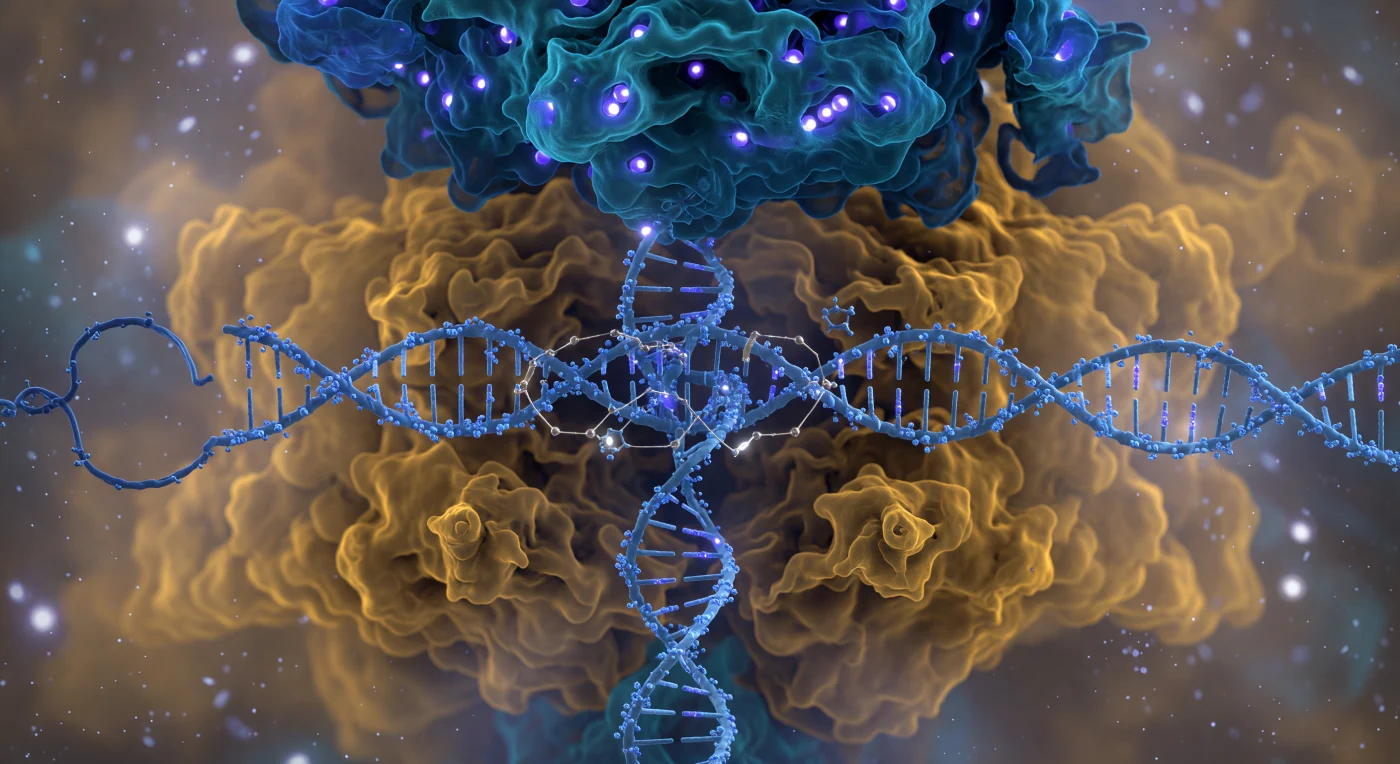
Te encuentras suspendido a apenas cinco nanómetros de la cara frontal del complejo Cas9, una maquinaria molecular de nueve nanómetros de anchura que llena absolutamente todo tu campo visual como una catedral de mineral vivo: el lóbulo de reconocimiento, azul cobalto y teal profundo, se arquea sobre ti como una bóveda de hélices-α que irradian una bioluminiscencia interior, mientras el lóbulo nucleasa se hincha por debajo en ámbar cálido y ocre, sus superficies difusas negociando su frontera con el solvente en lugar de trazarla con precisión, pues a esta escala la materia no tiene bordes sino nubes de densidad electrónica en perpetua negociación térmica. Atravesando el espacio entre los dos lóbulos, la doble hélice de ADN en conformación B pasa como una columna de cuerda retorcida en azul hielo, cada nodo del esqueleto fosfato titilando con carga negativa violácea mientras moléculas de agua se agrupan a su alrededor como satélites temblorosos que dispersan luz pálida en todas direcciones; el ARN guía la intersecta en cian eléctrico, sus grupos 2′-OH erizados hacia afuera como ganchos diminutos, y tres puentes del bucle R ya se han formado ante ti como hilos de luz dorada que sellan la complementariedad entre el ARN espaciador y la hebra molde del ADN. En el dominio de interacción con el PAM, dos dedos de arginina se presionan como paletas de cobre cargadas positivamente contra el trinucleótido NGG, distorsionando sutilmente la geometría local del surco menor, mientras en el núcleo catalítico los iones Mg²⁺ de los sitios activos HNH y RuvC arden como puntos blancos incandescentes, cada uno encerrado en una jaula octaédrica de oxígenos coordinados que tiemblan con precisión matemática, todo el conjunto bañado en la neblina fosforescente del solvente iónico que se extiende hasta una profundidad molecular infinita detrás de esta asombrosa máquina de edición genómica.

Te encuentras suspendido en el corazón geométrico de una proteína intrínsecamente desordenada, inmerso en una nube de probabilidad luminosa de unos ocho nanómetros de diámetro: no hay arquitectura sólida aquí, sino decenas de confórmeros polipeptídicos superpuestos a mínima opacidad, cuya presencia acumulada construye una nebulosa azul-blanca que respira y se desplaza sin jamás estabilizarse. A tu izquierda, una cinta de color ámbar cálido coalesce por apenas un nanosegundo —una alfa-hélice transitoria con sus oxígenos carbonílicos captando un brillo interior como cobre incandescente— antes de disolverse de nuevo en la niebla luminosa del ensamble conformacional, mientras un racimo de residuos aromáticos —fenilalanina, triptófano, tirosina— pulsa brevemente en amarillo dorado, un mínimo energético efímero que desaparecerá y reaparecerá en otro lugar de la nube en cuestión de microsegundos. El agua lo domina todo: moléculas esferoidales de apenas 0,28 nanómetros presionan contra cada segmento expuesto de la cadena principal en un mosaico denso y agitado, sus dipolos reorientándose sin cesar, su energía térmica colectiva el ruido de fondo contra el cual compite cada evento estructural transitorio de la proteína. Esta es la esencia del desorden funcional: no la ausencia de estructura, sino una superposición estadística de estados que el medio acuoso esculpe y demolece continuamente, manteniendo a la proteína perpetuamente entre el ser y el devenir.

Desde aquí abajo, mirando hacia arriba desde la base del tallo, la horquilla de ARN se alza como una catedral de bronce que gira sobre sí misma, su doble hélice en conformación A —compacta, inclinada, apenas 2,3 nanómetros de diámetro— llenando el campo visual hasta perderse en una niebla azul eléctrica donde el disolvente acuoso se convierte en algo cercano al infinito. La cadena azúcar-fosfato reluce en ámbar oscuro y cobre bruñido, y de cada unidad de ribosa brotan los grupos 2'-hidroxilo como pequeñas antenas de cobre cálido, señal inconfundible de que esto es ARN y no ADN, cada uno vibrando suavemente en su pozo de energía local a ritmos de femtosegundos. Las moléculas de agua se agolpan desde todos los ángulos como esferoides opalescentes y agitadísimos, mientras los iones de sodio trazan destellos plateados y los de magnesio se adhieren al esqueleto fosfato en cúmulos dorados, todos ellos formando la atmósfera electrostática que estabiliza esta estructura y la mantiene plegada. En lo alto, donde el tallo se abre en el tetraloop GNRA, la torre termina en una corona asimétrica de nucleótidos no apareados cuyas bases de adenina apiladas forman una plataforma sorprendentemente rígida bañada en un resplandor ámbar más suave, como el remate barroco de un campanario molecular. Todo tiembla de manera invisible a escalas de tiempo inimaginablemente cortas, y sin embargo este instante captura una perfección estructural absoluta: información codificada en geometría, mantenida viva por el mismo caos térmico que amenaza con deshacerla.
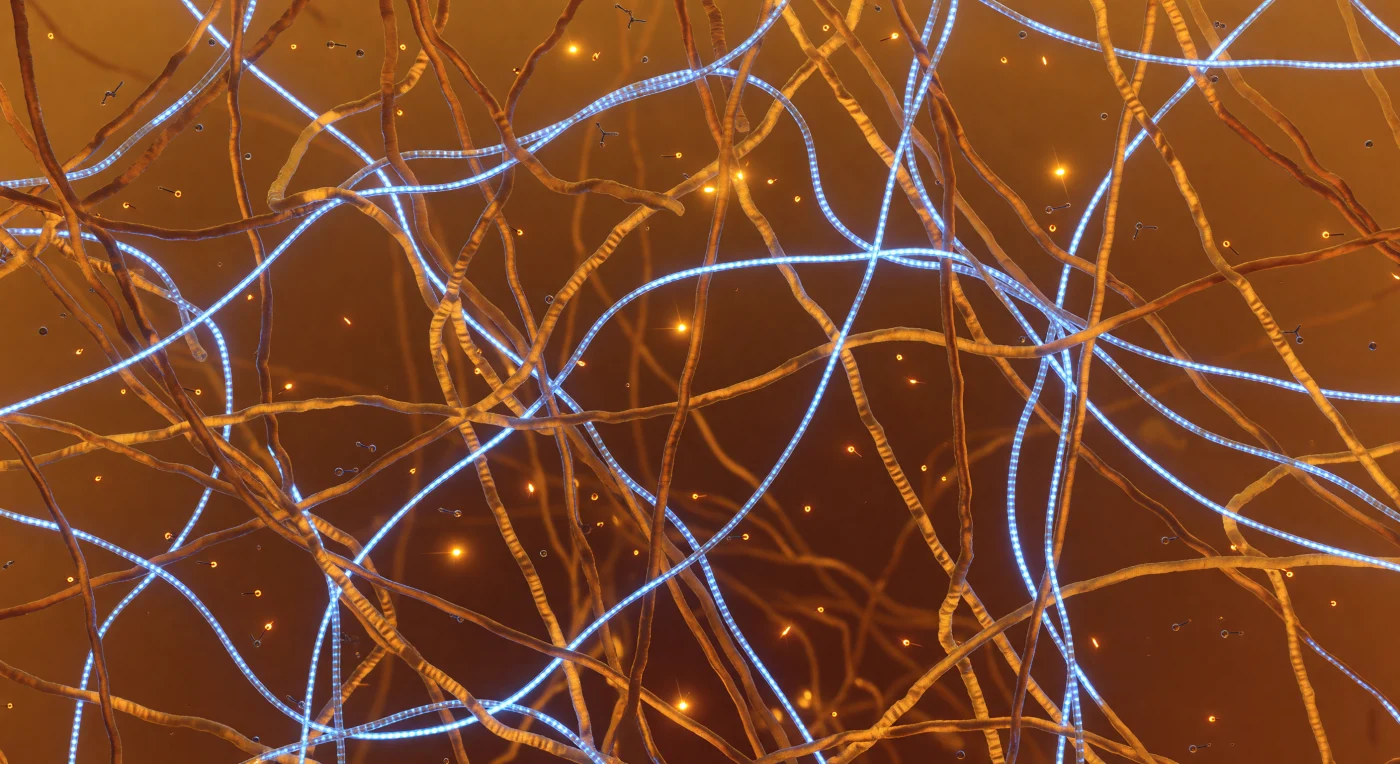
Te encuentras suspendido en el corazón líquido de un coacervado proteína-ARN, envuelto en una jungla ámbar de cadenas poliméricas desordenadas —dominios de baja complejidad que se entrelazan a dos o tres nanómetros de distancia en todas las direcciones, densos como cuerdas de seda envejecida impregnadas de luz de resina. Donde los anillos aromáticos de tirosina rozan los grupos guanidinio de arginina en contactos catión-π transitorios, destellan chispas de ámbar fundido que se apagan antes de que el ojo pueda seguirlas, efímeras como brasas avivadas por el aire, dispersas por todo el volumen medio de esta malla viviente. A través de ese tejido cálido serpentean hebras de ARN de 1,5 nanómetros de diámetro, cables de neón azul-blanco cuya luminiscencia estructurada revela cada 0,34 nanómetros los peldaños de las bases apiladas bajo la cadena principal fosfodiéster, proyectando reflejos azulados sobre las paredes ámbar más cercanas y creando halos de aguamarina allí donde el ácido nucleico y el polipéptido se disputan el mismo territorio. Moléculas de ATP derivan sin trayectoria aparente a través de los poros de cinco a quince nanómetros que se abren brevemente en la malla, arrastradas por el movimiento browniano en un medio acuoso cuya viscosidad se sitúa entre el agua libre y un gel blando, todo ello gobernado por el bombardeo térmico incesante de moléculas de agua que colisionan a centenares de metros por segundo. A cinco nanómetros la red ámbar se disuelve en bruma luminosa, y a ocho el mundo entero ha desaparecido en una niebla cálida y uniforme que parece generar su propia luz interior desde la energía térmica misma, transformando este espacio claustrofóbico y primordial en algo simultáneamente íntimo y eléctrico.
