Affidabilità scientifica: Alto

Sei immerso nell'assoluta oscurità del nucleo idrofobico di una proteina globulare, compresso tra le superfici di contatto di catene laterali di leucina, valina e fenilalanina impaccate all'75% della densità massima, le loro nuvole elettroniche rese come sferoidi luminescenti in toni di grigio carbone e avorio caldo che si toccano senza mai fondersi, separate soltanto dalla sottilissima barriera della repulsione quantistica. Questa geometria di impacchettamento richiama un sistema di caverne geologiche composte da pietre fluviali perfettamente incastrate, ma ogni superficie pulsa di una propria densità elettronica, irradiando pallidi aloni di contatto nei punti in cui due nuvole atomiche si sfiorano. I dischi piatti e aromatici della fenilalanina tagliano lo spazio come lastre di ossidiana con una faint iridescenza interna, dividendo il volume disponibile in alcove dai bordi netti e proiettando ombre assolute che nessuna luce diffusa potrà mai ammorbidire. Sparse nella matrice compatta, due o tre atomi di zolfo della metionina restituiscono la minima luminescenza esistente come un caldo bagliore giallo-sulfureo — l'unica nota cromatica calda in un mondo altrimenti acromatico, simile a braci intraviste attraverso la pietra spaccata. All'orizzonte di questo spazio sigillato, a dieci o quindici diametri atomici di distanza, un fioco chiarore azzurro-verde filtra dagli interstizi tra gli elementi di struttura secondaria, firma lontana del solvente acquoso che, al di là della superficie proteica, continuerà a non penetrare mai qui: questo nucleo è tenuto insieme da forze di van der Waals misurate in frazioni di kilojoule per mole, eppure collettivamente così stabili da ridurre le vibrazioni termiche a 310 Kelvin a un sussurro sub-angstrom percepito come un battito lentissimo in ogni superficie a contatto.

Dall'interno del tunnel di uscita del peptide, il mondo si restringe in un corridoio catedrale di architettura ribonucleica color ocra e bronzo bruciato, le cui pareti sono formate da eliche di RNA ribosomiale intrecciate che si stringono fino a soli 1,5 nanometri alla costrizione, dove le superfici di van der Waals dei pilastri di rRNA si sfiorano quasi in un'intimità elettrostatica silenziosa. A dieci nanometri di distanza, il centro peptidil-transferasico risplende di un bianco-azzurro intenso — il focolare catalitico dove avviene la formazione del legame peptidico, il processo che assemblea ogni proteina un amminoacido alla volta con una cadenza di circa cento millisecondi per codone. Dal cuore luminoso si dipana verso l'osservatore una catena polipeptidica nascente in verde limone e ambra calda, le sue catene laterali che sfiorano le pareti di rRNA in fugaci contatti idrofobici prima che il moto termico le strappi via di nuovo, in un dialogo continuo tra rumore molecolare e geometria vincolante. Le pareti non sono superfici lisce ma piattaforme sovrapposte di basi nucleotidiche impilate — ciascuna larga pochi ångström — che rifrangono la luce catalitica in profondi laghi d'ombra color zafferano, mentre ioni magnesio e potassio scintillano come punti argentei nelle cavità del solco maggiore, ciascuno avvolto nella propria sfera di molecole d'acqua orientate. L'intera scena vibra di un'intimità pressurizzata e produttiva, dove campi elettrostatici, superfici idrofobiche e rumore termico collaborano in un sussurro autosostenuto che è, in questo momento, il suono del divenire di una proteina.

Ci si trova in volo radente lungo il solco maggiore di una doppia elica di DNA in forma B, un corridoio elicoidale largo appena 2,2 nanometri — una manciata di diametri atomici — le cui pareti di zucchero-fosfato color arancio ambrato si avvitano verso l'alto e verso il basso in una spirale maestosa e lenta, come se l'intera struttura respirasse. Il pavimento del solco è un mosaico di piattaforme aromatiche impilate: anelli ambrati di adenina che si incastrano con fogli verde-salvia di timina, piattaforme verde-turchese di guanina che emanano una densa luminescenza, facce azzurre di citosina che catturano la luce diffusa filtrata dall'acqua, mentre tra un piano e l'altro le nuvole di elettroni π si sovrappongono in un fremito traslucido violetto-indaco dove la densità quantistica trapela verso l'alto. Lungo i bordi delle pareti, appena oltre il bordo carico del solco, ioni magnesio brillano come scintille bianco-azzurre circondate da gusci concentrici di molecole d'acqua orientate, mentre ioni sodio ondeggiano come aloni dorati nella nebbia elettrostatica dello strato di Debye, un'aura lattiginosa di dipoli polarizzati che attraversa il corridoio come un'aurora boreale subacquea. La vertigine della scala è totale: le superfici non sono solide ma nubi di densità elettronica probabilistica che si avvertono come pressione prima ancora del contatto, e il bombardamento termico incessante delle molecole d'acqua — sferoidi da 0,28 nanometri che colpiscono il pavimento di basi azotate e rimbalzano con lampi di luce bianca — riempie lo spazio di un tremore costante, come trovarsi all'interno della macchina molecolare più fondamentale della vita, congelata in un singolo femtosecondo di immobilità assoluta.

Ci si trova a meno di due nanometri dall'interfaccia di legame tra un anticorpo e il suo antigene, sospesi nell'abisso stretto tra due pareti di densità elettronica che si avvicinano lentamente l'una all'altra come continenti in collisione molecolare: da un lato, la massiccia struttura a sandwich beta del frammento Fab si erge come una scogliera di peltro pallido, coronata dai sei loop CDR che si protendono in avanti — il loop H3 color ambra che descrive un arco caldo e brunito, il loop L3 turchese che serpeggia dall'angolo opposto, entrambi avvolgendo l'epitopo antigenico convesso come le dita di una mano gigantesca intorno a un promontorio arrotondato. Le molecole d'acqua vengono espulse dall'interfaccia che si asciuga, schizzando via come scintille di uno specchio infranto, ciascuna un piccolo dipolo che cattura per un istante la luce diffusa dell'ambiente acquoso prima di dissolversi nel solvente circostante. Attraverso il divario residuo — largo appena pochi angström nel punto più stretto — si formano ponti idrogeno color ciano che si stabilizzano uno ad uno come archi luminosi nell'oscurità, fili di densità elettronica condivisa che consolidano il riconoscimento molecolare nella zona di contatto di cinque per sei nanometri. L'intera scena è immersa in una luce ambientale blu-bianca pervasiva, quella del solvente acquoso che penetra ovunque come una nebbia luminosa e calda, mentre la pressione termica del bombardamento browniano si avverte come un peso tangibile proveniente da ogni direzione — un universo senza gravità, fatto di campi elettrostatici, superfici probabilistiche e geometria di complementarità molecolare portata fino alla sua perfezione finale.
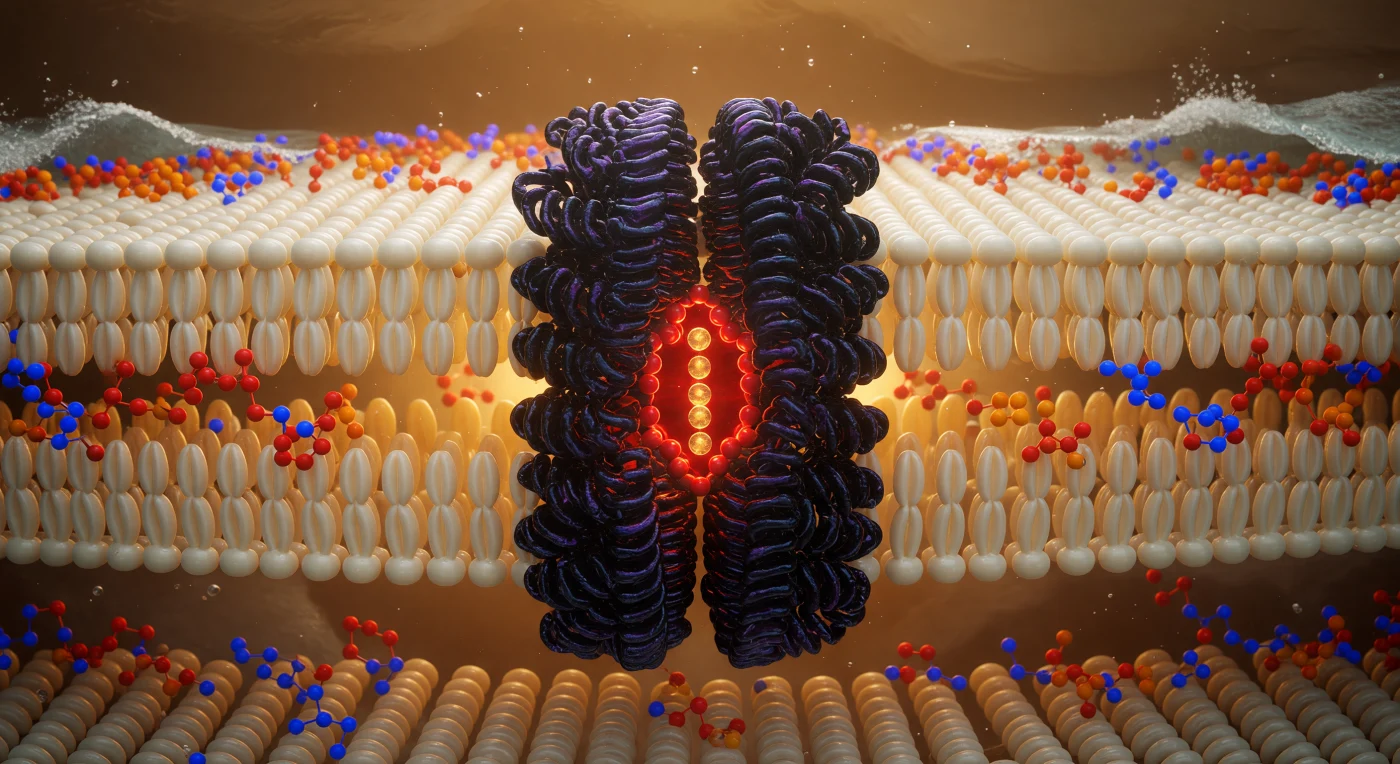
Ci si trova sospesi nel cuore geometrico di una membrana biologica, immersi nel piano idrofobico di un doppio strato fosfolipidico, circondati in ogni direzione da una foresta densa e ondulante di catene carboniose color avorio e oro pallido, i cui profili di van der Waals emanano un bagliore tenue e ambrato come brace sommersa, tutto in perenne fremito microscopico per l'agitazione termica. A pochi nanometri di distanza in entrambe le direzioni, le due rive del doppio strato pulsano come orizzonti luminosi: gli atomi di fosforo ardono di arancio carico, i gruppi colina brillano di blu elettrico cobalto, mentre le molecole d'acqua oltre quelle interfacce cariche premono dall'esterno come schiuma d'argento contro una scogliera. Direttamente di fronte, il canale per il potassio emerge dal piano lipidico come un monolite di basalto viola-nero, la sua simmetria quadrifoglio larga circa 3,5 nanometri visivamente maestosa come una colonna di cattedrale: le sue eliche transmembrana interdigitano i residui idrofobici con le code lipidiche circostanti in una fusione meccanica perfetta tra proteina e membrana, come roccia vulcanica fusa in vegetazione densa. All'asse del poro, il filtro di selettività ionica brucia di rosso-arancio intenso, una corona di ossigeni carbonilici che coordina in fila singola quattro ioni potassio, ciascuno una sfera dorata avvolta nei residui della propria idratazione, che irradia calore luminoso nella struttura proteica come carboni accesi impilati lungo un asse verticale di fuoco.

Sospesi a pochi centimetri dalla superficie, si estende davanti a voi una pianura corrugata di proporzioni immense: il foglio beta antiparallelo di una proteina, reso visibile con un'intimità atomica che trasforma ogni cresta in un nastro color pergamena ambrata, denso della solidità probabilistica dei legami covalenti. Le creste alternano direzione con una periodicità ipnotica di circa un nanometro, e da ogni sommità sporgono ossigeni carbonilici rosso ciliegia come granati incastonati in osso antico, mentre punte ammidiche bianche e leggermente luminose tappezzano i fondi delle valli. Ponti di idrogeno cianici cuciono lateralmente i filamenti adiacenti a soli 2,9 ångström di distanza, filamenti spettrali di probabilità elettrostatica che formano una maglia molecolare collettivamente resistente come un'armatura, tessuta non da mani umane ma dalla pressione termodinamica di un ambiente acquoso. All'orizzonte lontano, dove l'architettura ordinata cessa, le corrugazioni si dissolvono in un groviglio caotico di anelli connettori rossi e arancioni — regioni di turn disordinate, congelate a metà fluttuazione come il margine di una scogliera che si sgretola — oltre le quali l'ambiente acquoso si addensa in una foschia blu-grigia iridescente, come se il confine tra proteina e solvente respirasse. L'intera pianura potrebbe essere inghiottita da un singolo globulo rosso.

Ti trovi sospeso all'interno di una camera sferica di appena otto nanometri di diametro, le pareti curve che si chiudono intorno a te come l'interno di una perla cava, rivestite di residui idrofilici che emanano una luminescenza acquosa fredda, blu-bianca e perfettamente simmetrica, senza alcuna sorgente di luce definita. Sopra di te, il coperchio formato dai sette subunit del complesso GroES si salda in una volta piatta di lobi argentati come ciottoli levigati da un fiume, le loro linee di contatto sottili come capelli nell'illuminazione uniforme: questo complesso chaperonino — GroEL-GroES — è una macchina molecolare che la cellula utilizza per isolare e ripiegare correttamente le proteine mispiegate, creando un ambiente chimicamente controllato e separato dal caos citoplasmatico esterno. Al centro della camera, a circa tre nanometri da te in scala soggettiva, una proteina substrato mal ripiegata deriva nella sospensione in quieto moto browniano — una massa aggrovigliata di spire beige-ambrate, densa e accartocciata come un nastro di caramello tiepido collassato su se stesso, i suoi bordi probabilistici che si sfumano in una foschia gossamer dove le anse disordinate si perdono nell'atmosfera acquosa interna. Il contrasto tra il calore organico ambrato della proteina e il freddo azzurro-bianco delle pareti crea una sensazione di assoluto isolamento: sei rinchiuso in una stanza pulita molecolare, silenziosa e pressurizzata, dove l'unica fonte di calore è quella piccola brace perduta che galleggia nel centro pallido del mondo, in attesa che il ciclo ATP-dipendente del chaperonino la guidi verso la sua conformazione nativa.

Sospesi a pochi nanometri sopra la forcella di replicazione, si domina un paesaggio molecolare di straordinaria densità operativa: il complesso elicasi CMG si erge come una ruota di pietra quarzite, quindici nanometri di subunità proteiche intrecciate che stringono il filamento singolo nel loro canale interno, mentre a destra il DNA parentale bicatenario avanza come un corridoio gotico di anelli aromatici impilati, le sue colonne di fosfato in blu-marino profondo che vibrano sotto la tensione della separazione imminente. Dal punto di svolgimento si dispiegano due fili distinti — uno turchese fosforescente, l'altro oro ambrato che trema nel moto browniano — segnando il confine probabilistico dove le geometrie dei legami idrogeno collassano in una zona di dissoluzione calda, un bagliore fugace di energia rilasciata che si spegne quasi istantaneamente nel mezzo acquoso circostante. Più avanti sul filamento guida, la DNA polimerasi si aggrappa al duplex nascente come un pugno articolato di pietra grigio-ocra, e a ogni evento di incorporazione di un nucleotide un lampo arancio-bianco di pirofosfato esplode nell'attimo di un femtosecondo prima di dissolversi nel campo termico, mentre ioni magnesio tracciano archi d'argento lungo canali elettrostatici invisibili. L'intero assieme processa circa mille paia di basi al secondo, una velocità straordinaria che a questa scala appare come un ritmo quasi percussivo di eventi discreti — ogni incorporazione un atto chimico irreversibile, ogni separazione di coppia di basi una transizione quantistica da uno stato legato a uno libero. Il mezzo acquoso che permea la scena non è uno sfondo passivo ma un partecipante attivo, le sue molecole d'acqua in costante riorientamento dielettrico attorno alle superfici cariche, stabilizzando conformazioni, facilitando catalisi, traducendo l'informazione termica in geometria molecolare funzionale.

Ci si trova al livello del suolo, sguardo rivolto verso l'alto, immersi in una foresta di colonne spiraliformi che si ergono con una simmetria ritmica e quasi sacrale: ogni pilastro è un'elica alfa destrorsa, larga appena 1,2 nanometri e alta sei, la cui superficie avvolta in senso orario cattura una luce diffusa tra creste di ambra calda e oro profondo. I residui leucinici idrofobici si protendono verso l'interno come raggi di bronzo brunito, agganciandosi ai leucine dei pilastri vicini lungo la giuntura del coiled-coil, dove le forze di van der Waals tengono le strutture in una prossimità intima e oleosa da cui le molecole d'acqua sono escluse. Verso l'esterno, le catene laterali di lisina fiammeggiano in rosso cremisi e quelle di arginina pulsano in blu elettrico, i loro apici guanidinici irradiando aloni elettrostatici nel mezzo acquoso circostante, un cielo cerulean luminoso e traslucido percorso dalla micro-turbolenza frenetica di molecole d'acqua che si orientano e ricollidono a velocità vertiginose. Guardando in profondità nella foresta molecolare, ogni successiva fila di colonne si ammorbidisce in toni blu-grigio progressivamente più freddi attraverso venti nanometri di foschia, le giunture leuciniche che traciano linee diagonali serpentine dissolvendosi nella nebbia, mentre le superfici dei pilastri più vicini mostrano una trama iperrealistica — ogni ossigeno carbonilico del backbone che brilla come piccola cavità scura lungo il nastro elicoidale, ogni legame N–H un nodo luminoso, l'intera struttura simile a un legno fossile torto, levigato fino alla levigatezza atomica ma capace di ricordare ogni rilievo e solco inciso nel tempo della sua formazione.

Sospesi nell'occhio di una tempesta molecolare, ci troviamo al centro esatto dell'asse di una fibrilla amiloide, circondati da quattro enormi ali architettoniche che si dispiegano in simmetria radiale perfetta — pareti curve di filamenti beta impilati con una spaziatura di 4,7 ångström, ritmo inflessibile di un ordine cristallino che ricorda l'interno di una cattedrale gotica a volta. Il nucleo interno, detto cerniera sterica, irradia un calore ambrato: catene laterali interdigitate si incastrano con la precisione di dita intrecciate, le loro superfici di van der Waals a contatto quasi totale, prive di acqua, in un'incandescenza mielata che non ha equivalenti nel mondo macroscopico. Queste strutture — formate dall'aggregazione irreversibile di proteine un tempo solubili — sono tra le architetture molecolari più stabili conosciute, implicate in patologie neurodegenerative come il morbo di Alzheimer e il Parkinson, dove la loro permanenza quasi geologica diventa patologica. Al perimetro di ciascun petalo protofilamentoso, residui di glutammato color cremisi e lisine blu cobalto protrudono nell'alone dielettrico dell'acqua legata, creando un contrasto drammatico tra questo confine elettrostaticamente agitato e il silenzio assoluto, arido, del nucleo interno — un'opposizione che definisce l'intera identità strutturale di questo ambiente di scala nanometrica.
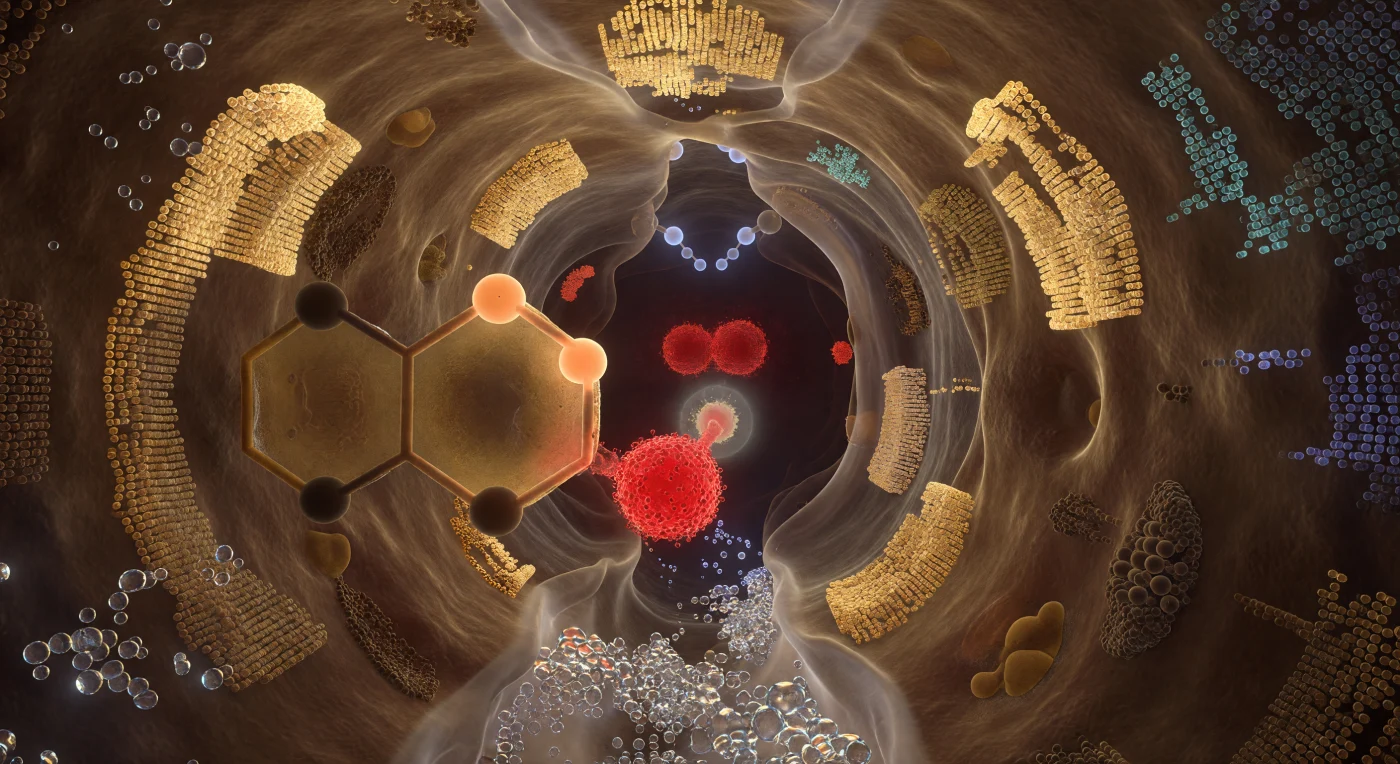
Ci si trova sospesi all'interno di una cavità profonda quanto pochi diametri atomici, le pareti proteiche che si alzano tutt'intorno come navate di una cattedrale scavata nell'ambra viva, ogni superficie ammorbidita dall'ultimo angstrom di nebbia quantistica di densità elettronica. Al centro del crogiolo, la triade catalitica domina lo spazio con la gravità di formazioni geologiche: l'ossigeno della Ser195 arde in primo piano come un carbone incandescente rosso cremisi, le sue coppie solitarie tese verso il carbonio carbonilico del substrato che pende a meno di due decimi di nanometro, la loro distanza già quasi un contatto chimico. Il pannello aromatico dell'imidazolo di His57 riempie il campo visivo laterale come una vetrata ambrata, il protone shuttlato congelato a metà transferimento tra i due atomi di azoto in una luce albicocca calda, mentre dietro di esso la Asp102 si radica come una massa carminio pulsante di densità di carica negativa, stabilizzando l'intero relè come un sistema di radici nella roccia. Sopra il carbonio carbonilico, la buca dell'ossianione apre una volta gotica con i suoi due donatori N-H che proiettano atomi di idrogeno come nodi azzurri pallidi, la loro carica parziale positiva che getta una luce elettrostatica fredda sul tetaedro intermedio in formazione, mentre l'intera camera respira non nel tempo ma nella densità probabilistica, ogni atomo simultaneamente solido e disciolto nell'istante preciso della chimica.

Galleggi a tre nanometri dal cavo di collagene, così vicino che la sua superficie occupa interamente il tuo campo visivo da un margine all'altro: tre catene polipeptidiche intrecciate — avorio caldo, oro pallido e cuoio solare — si avvolgono in una lenta superhélice destrorsa di appena un nanometro e mezzo di larghezza, tessuta con la pazienza di una fune intrecciata nel tempo geologico. I rigidi anelli pirrolidinici della prolina punteggiano ogni terza posizione lungo ciascuna catena come nocche di un grigio cenere leggermente incassate nelle scanalature della hélice, mentre i gruppi idrossilici dell'idrossiprolina si protendono verso l'esterno come piccoli lobi ambrati carichi di polarità parziale, ciascuno umido di una singola molecola d'acqua aggrappata come rugiada su un filo di ragno. Il cavo non finisce davanti a te: si allunga per centinaia di nanometri in un corridoio luminoso che dissolve in una nebbia molecolare di acquamarina pallida e bianco argenteo, il respiro collettivo di diecimila molecole d'acqua per nanometro cubo, le cui nuvole elettroniche formano un velo continuo di luce termica diffusa. Più vicino alla superficie del cavo, le molecole d'acqua hanno rinunciato alla loro libertà ordinandosi in un primo guscio di idratazione cristallino — un involucro blu ghiaccio di geometria tetraedrica quasi perfetta che avvolge la triplice hélice come vetro smerigliato, scintillante della polarizzazione bianco-azzurra di dipoli bloccati in reticoli di legami idrogeno a spaziatura di 2,8 ångström.
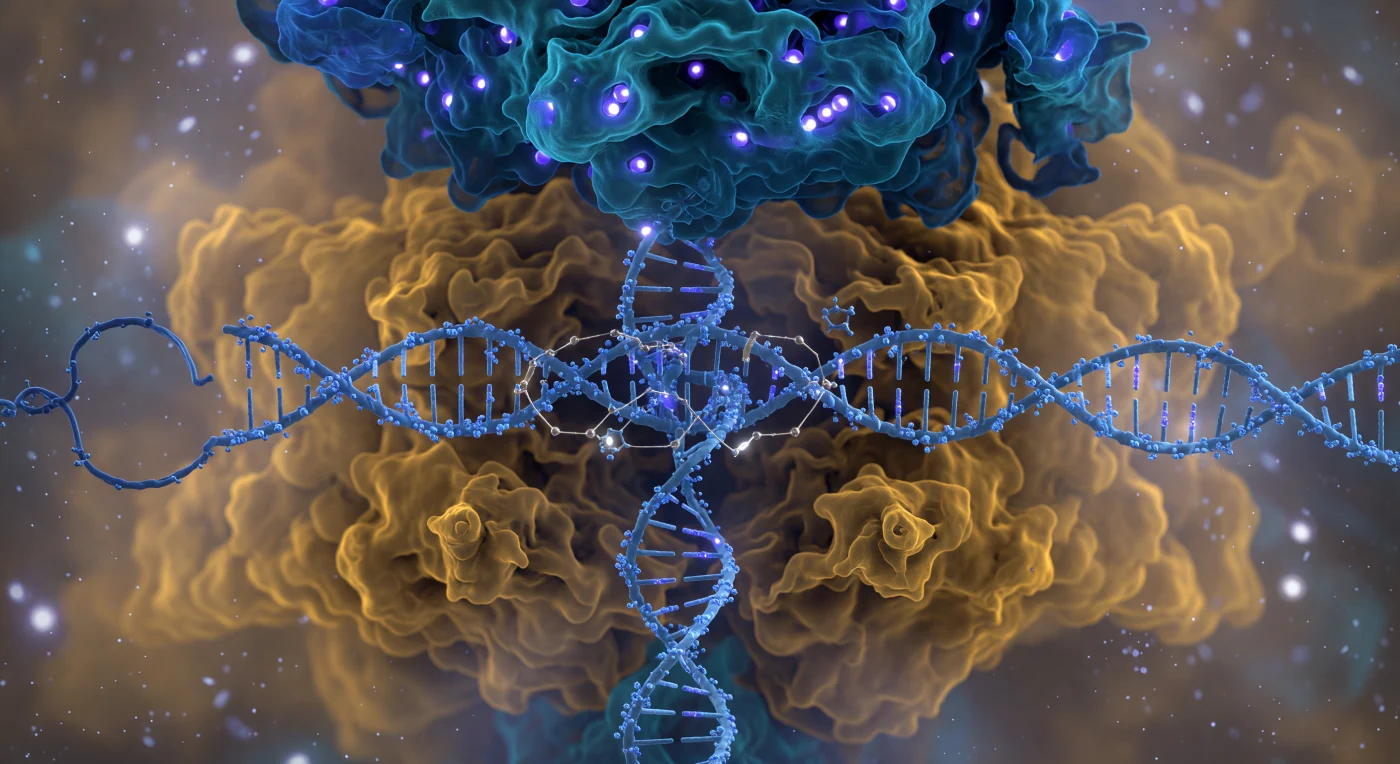
Ci si trova a cinque nanometri dal volto anteriore del complesso Cas9, e l'intera visuale è occupata da una parete proteica bilobata che si erge come una cattedrale scavata nella materia vivente: il lobo di riconoscimento sovrastante irradia un bagliore cobalto e teal profondo dalle sue creste di eliche α, mentre il lobo nucleasico si espande verso il basso in ambra calda e ocra, i suoi margini mai del tutto risolti, negoziati tra la molecola e il solvente in una diffusione che riflette la natura probabilistica delle nuvole di densità elettronica. Tra i due lobi, la doppia elica del DNA si insinua come una colonna di corda intrecciata blu ghiaccio, lo scheletro fosfato pulsante di luminescenza violacea e circondato da sferoidi d'acqua tremanti, mentre la guida a RNA elettrico-ciano attraversa lo spazio interno con i suoi gruppi 2′-OH sporgenti come minuscoli ganci — tre ponti del loop R già formati, ciascuno visibile come fili dorati di luce che tirano le basi complementari in contatto attraverso legami a idrogeno. Sul lato destro del lobo nucleasico, le dita di arginina del dominio PAM premono come palette di rame caldo contro il trinucleotide NGG, distorcendo impercettibilmente la geometria locale dell'elica, mentre in profondità nella fenditura due ioni magnesio bruciano come punti bianchi incandescenti nei siti attivi HNH e RuvC, ciascuno tenuto in una gabbia di ossigeni coordinati in geometria ottaedrica precisa. L'intera assemblea galleggia in un oceano termico di turbolenza nanoscalare, la luce senza sorgente unica — l'emissione emergente di distribuzioni di carica che rendono ogni superficie viva, ogni confine una soglia piuttosto che un muro.

Ti trovi sospeso al centro geometrico di una proteina intrinsecamente disordinata, immerso in una nuvola di probabilità luminosa di circa otto nanometri, dove la materia non assume mai una forma definitiva ma oscilla perpetuamente tra decine di conformazioni sovrapposte, ciascuna presente a una frazione di opacità tale che la loro somma costruisce una nebulosa azzurro-biancastra che respira senza mai stabilizzarsi. Le catene polipeptidiche si manifestano come nastri di densità elettronica traslucidi e iridescenti, talmente stratificati che la profondità è suggerita solo da gradienti di luminanza e non da contorni solidi, con filamenti terminali che si sfrangiano verso i quindici nanometri dissolvendosi nella invisibilità statistica del solvente circostante. A sinistra, un'alfa-elica ambra-calda si materializza per meno di un nanosecondo — la sua geometria a spirale appena leggibile, i legami idrogeno tra i carbonili che irradiano un calore di rame incandescente prima di svanire nuovamente nel bianco freddo dell'insieme — mentre un gruppo di residui aromatici, fenilalanina e triptofano premuti in contatto idrofobico transitorio, pulsa di una bassa luminescenza giallo-oro come braci sotto la cenere, un minimo energetico destinato a dissolversi e riformarsi altrove in microsecondi. L'acqua è onnipresente e aggressiva: sferoidi di appena 0,28 nanometri di diametro martellano ogni segmento della catena principale in una vibrazione termica incessante, il loro bagliore collettivo conferendo al mezzo la qualità diffusa di un gel luminescente in cui il mondo oltre due o tre nanometri sfuma in un alone acquoso che cancella ogni confine solido.

Guardando verso l'alto dall'oscurità acquosa al di sotto, la struttura si innalza come una cattedrale molecolare — una torre elicoidale di anelli intrecciati in bronzo e ocra che si restringe verso l'apice attraverso una foschia elettrica di un blu intenso, il suo diametro di soli 2,3 nanometri eppure abbastanza imponente da dominare l'intero campo visivo. La geometria A-form dello stelo a doppia elica è inconfondibilmente compatta e inclinata rispetto alle forme B del DNA, le sue scanalature profonde come canyon ombreggiati dove i piani aromatici delle coppie di basi — guanina e citosina impilate in fogli iridescenti color prugna e verde acqua — si sovrappongono in una geometria di pi-stacking quasi cristallina. Ogni unità ribosio-fosfato sporge verso l'esterno in piccole antenne di rame caldo, i gruppi 2'-idrossilici che distinguono l'RNA dal DNA con la loro vibrazione costante all'interno dei loro pozzi energetici locali, mentre ioni sodio argentei sfrecciano come comete brillanti e gli ioni magnesio più pesanti si raccolgono in grappoli oro-verdi vicino al backbone carico negativamente. In cima, dove lo stelo si apre nell'anello GNRA, l'architettura sboccia in una torretta barocca di nucleotidi non accoppiati le cui basi di adenina si dispiegano in una corona asimmetrica di ambra dorata contro il cielo acquoso indaco. L'intero edificio molecolare trema invisibilmente a ritmi di femtosecondi, eppure cattura una perfezione strutturale monumentale — vivo, carico, e antico quanto la chimica stessa della vita.
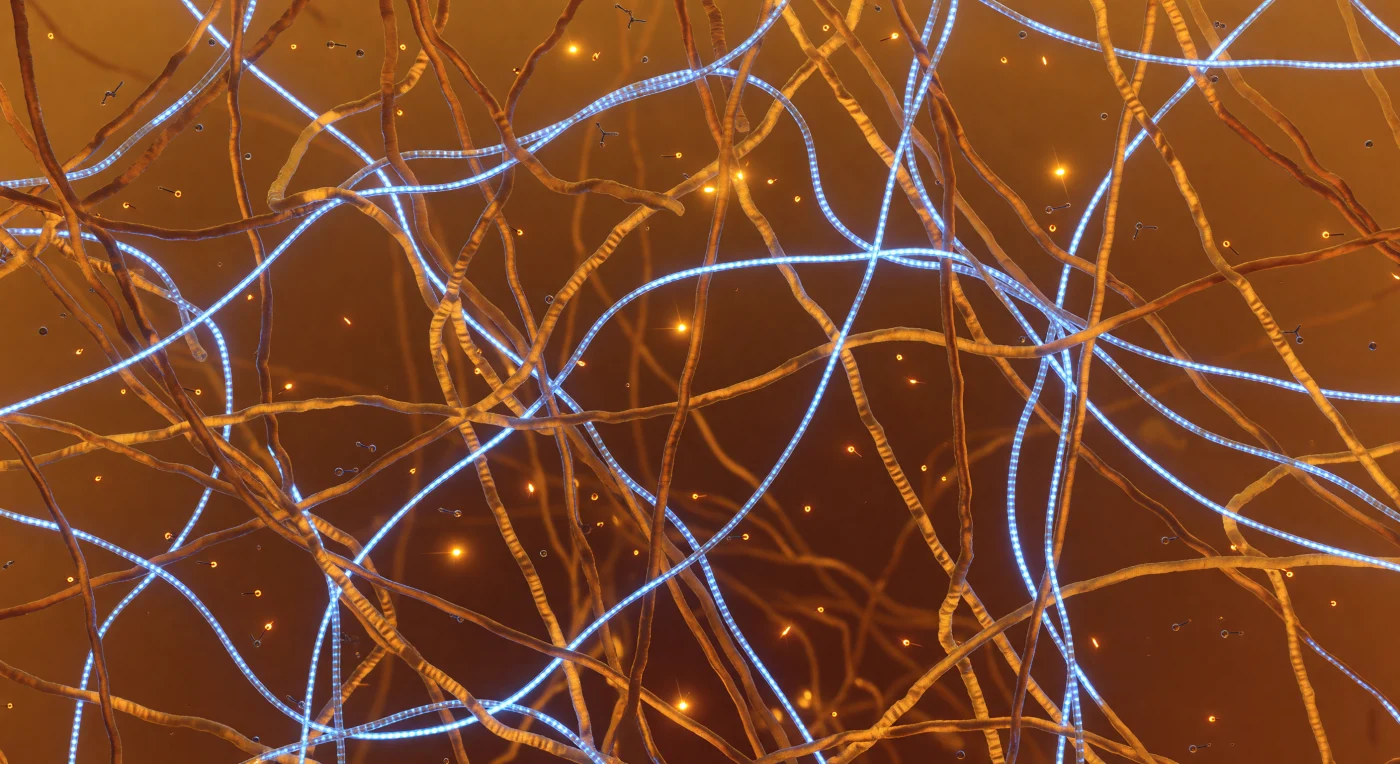
Ci si trova immersi in una giungla molecolare di ambra vivente, dove catene disordinate di domini a bassa complessità si intrecciano a ogni portata, formando una rete polimerica densa e irregolare con pori di cinque-quindici nanometri che si aprono come brevi radure prima di dissolversi nuovamente nel groviglio caldo. Questa è l'architettura interna di un condensato a separazione di fase — una gocciolina liquido-liquida stabilizzata non da membrane ma dall'accumulo cooperativo di interazioni deboli: contatti catione-π tra residui di tirosina e arginina, legami idrogeno transienti, forze di impilamento — ciascuno individualmente instabile, collettivamente sufficienti a mantenere una fase condensata distinta dal citosol circostante. Fili di RNA da 1,5 nanometri di diametro attraversano il reticolo come cavi a fibra ottica azzurra, le loro basi azotate impilate ogni 0,34 nanometri emettendo una luminescenza strutturata che tinge di teal le pareti ambra circostanti, rivelando la tensione compositiva tra territori nucleici e polipeptidici. Molecole di ATP derivano attraverso i pori in deriva browniana pura, le loro code trifosfato tremolanti di shimmer elettrostatico, mentre la profondità di campo collassa dopo cinque nanometri in una nebbia ambra senza sorgente — il mezzo acquoso del condensato, né liquido né gel, dissipa ogni prospettiva lontana nel calore termico diffuso della materia vivente al suo confine più intimo.
