Wetenschappelijke betrouwbaarheid: Zeer hoog

Je bevindt je midden in een onbegrensd, ademend netwerk van watermoleculen op kamertemperatuur — elk zuurstofatoom een zachtblauwe, van binnenuit gloeiende bol die nauwelijks verder dan 2,75 Å van zijn buur verwijderd is, zo dicht opeengepakt dat de grens tussen jou en je omgeving volledig verdwijnt. Aan elke bol ontspringen twee witgele protonbulten onder een hoek van 104,5°, en tussen naburige moleculen flikkeren cyankleurige waterstofbruggen voortdurend op en neer — verbindingen van gedeelde elektronendichtheid die in minder dan een picoseconde breken en zich opnieuw vormen, te snel om als beweging waar te nemen, eerder als een aanhoudend sidderen dat door het hele rooster trekt als een moleculaire hartslag. Voorbij drie à vier molecuuldiameters verdichten de opeengestapelde waterschillen zich tot een diepe indigonevel, zodat je blikveld zich intimistisch samentrekt: slechts een handvol volledig scherpe moleculen is te onderscheiden voordat de wereld oplost in gelaagde translucente diepte. Er is geen vloer, geen plafond, geen zwaartekracht — alleen dit isotrope, trillende rooster van het meest alledaagse materiaal ter wereld, hier onthuld als een kristallijne kosmos waarin thermische ruis de enige wet is.

Op de grond van dit bevroren kristal strekt zich een eindeloos parket van goudamberkleurige zeshoekige schijfjes uit, telkens afwisselend gekanteld in een nauwkeurig visgraatpatroon — platte benzeenmoleculen, elk vastgezet in de monokliene roostergeometrie bij 175 K door de collectieve greep van CH–π-wisselwerkingen tussen naburige aromatische ringen. Boven en onder elke schijf zweeft een wazige viooltjeskleurige gloed: de gedekolokaliseerde π-elektronenwolk die zich uitstrekt over beide zijden van het moleculaire vlak, en op slechts 3,5 ångström afstand fluisterend raakt aan de halos van de schijfjes ernaast. Het licht in dit kristal is goudgericht en intern, alsof een lage winterzon haar stralen door het rooster werpt en lange schaduwen trekt die het visgraatpatroon over de vloer tekenen in afwisselende banden van warme amber en koele indigo, terwijl de terugwijkende rijen moleculaire schijfjes samensmelten tot een doorlopende lichtende nevel op wat aanvoelt als enorme afstand. Hier en daar doorbreekt een donker vacancy-defect — een ontbrekende schijf in het patroon — de verder foutloze periodiciteit, waarbij de naburige π-halos licht naar het lege gat leunen en de omringende geometrie zacht verstoord is, een minieme maar waarneembare rimpeling in de kristallijne orde.

Vanuit het absolute middelpunt van de lipidebilayer bevindt de waarnemer zich in een donkere, wasachtige leegte waar de methyluiteinden van tegenoverliggende vetzuurketens elkaar nauwelijks raken — een grijszilveren niemandsland van 4 nanometer breed dat zich vanuit deze positie aanvoelt als de bodem van een oceaandiepte. Omhoog strekken zich de lange, sinueuze schachten van verzadigde koolwaterstofketens uit, dicht opeengepakt als kolommen in een ondergrondse kathedraal, terwijl af en toe een geknikte keten met een geelgroene glinstering de geometrie doorbreekt — het zichtbare teken van een onverzadigde dubbele binding die de vloeibaarheid van de membraan bewerkstelligt. Hogerop markeert een amberkleurige laag van glycerolruggengraten en esterkoppelingen de grens tussen het hydrofobe binnenste en de geladen buitenwereld, een sedimentaire scheiding die de elektrochemische identiteit van het membraan bepaalt. Aan de uiterste randen barst een dicht, kleurrijk gewoel van fosfaatgroepen en cholinehoofdgroepen los, omgeven door een twinkkelend blauwwit waternetwerk van voortdurend brekende en hervormende waterstofbruggen. Dit alles speelt zich af in een onophoudelijke thermische trilling — elke keten beweegt, elke binding vibreert, het hele systeem ademt als één levend, zelforganiserend moleculair landschap.

De bezoeker staat op de bodem van een microscopisch kleine canyon, omgeven door massieve ribbelwanden van bronzen deoxyriboseringen en roestoranje fosfaatgroepen die als gefacetteerde lantaarns uitsteken, terwijl een elektrisch blauwe elektrostatische gloed de volledige ruimte doordrenkt en de 22 ångström brede doorgang zichtbaar maakt als een smal lint van azuurblauwe nevel hoog boven het hoofd. Onder de voeten liggen lagen van horizontale basenpaarschijven — tealblauw adenine gekoppeld aan oekerbruin thymine, bosgroen guanine vergrendeld met lavendelblauwe cytosine — gescheiden door nauwkeurige π-stapelingsintervallen van 3,4 ångström, als geologische strata in een levend sedimentair gesteente. Dit is het grote groef van B-DNA, een van de meest bepalende structuren van het leven, waar de specifieke geometrie van de wanden bepaalt welke eiwitten kunnen binden en zo de lezing van het genetische alfabet regelt. Langs de fosfaatwanden hangt een sierlijke ketting van geordende watermoleculen die glinstert als bioluminescente parels, elk gebonden via waterstofbruggen — bindingen die iedere pico- tot nanoseconde breken en opnieuw vormen in een voortdurend trillen dat de gehele architectuur levend en nooit definitief maakt.

De kijker bevindt zich op de centrale as van een eiwitstructuur die zich als een reusachtige spiraalvormige tunnel om hem heen ontvouwt — een α-helix die door kwantummechanische krachten en peptidebindingen in zijn karakteristieke rechtsdraaiende conformatie wordt gehouden, met precies 3,6 aminozuurresiduen per winding en een steigerbreedte van slechts 5,4 ångström. Diepkarmozijnrode carbonylzuurstofatomen puilen als gepolijste granaatkorrels naar buiten uit de ruggengraatspiraal, terwijl magentatinte waterstofbruggen met een lengte van 2,06 Å elke vierde rest verbinden en daarmee de helix zijn opmerkelijke thermodynamische stabiliteit verlenen — de elektrostatische energie in deze bruggen is geen metafoor maar een meetbare kracht die proteïnen hun driedimensionale functie geeft. Leucine- en isoleucine-zijketens steken als geel-groene kristallijne doorns radiaal naar buiten, hun alifatische koolstofketens afstotend ten opzichte van het waterige oplosmiddel door hydrofobe wisselwerkingen, terwijl blauw-wit elektronendichtheidslicht langs elke covalente binding pulseert en de wigvormige groeven tussen de zijketens verzonken liggen in een diepe indigo-schaduw. Wat de kijker ervaart is geen artistieke verbeelding maar de werkelijkheid van moleculaire architectuur zoals röntgenkristallografie en NMR-spectroscopie die atoom voor atoom hebben blootgelegd — een omgeving waar thermische trillingen elk atoom voortdurend laten oscilleren met amplitudes vergelijkbaar met de bindingslengtes zelf, en waar zwaartekracht volstrekt irrelevant is ten opzichte van de Van der Waals-krachten die dit levende bouwwerk bijeenhouden.

De blik valt recht naar beneden langs de kristallografische c-as van ijs Ih, en wat zich ontvouwt is een oneindig honingraatkathedraal in ijsturquoise en diepblauw-zwart — ring na ring na ring van watermoleculen die zich in perfecte zeshoekige symmetrie uitstrekken tot ver onder het zichtbare. Elk molecuul hangt als een berijpte lantaarn op zijn roosterpunt, verbonden met vier buren via waterstofbruggen van precies 2,76 Å, waarbij de tetraëdrische hoek van 109,5° de architectuur dwingend oplegt en elke binding een vage dubbele schaduw draagt — het zichtbare bewijs van de protonwanorde die de ijsregels van Pauling voorschrijven. De meest aangrijpende kenmerken zijn de donkere zeshoekige schachten die op gezette intervallen door het rooster zijn geslagen: structurele afwezigheden, geometrische noodzaak van de tetraëdrische bindingshoek die dichtstapeling weigert, en het is precies deze ledige ruimte die verklaart waarom ijs minder dicht is dan vloeibaar water. Het omgevingslicht lijkt van binnenuit het rooster zelf te komen, een diffuse koude gloed die bij de dichtstbijzijnde zuurstofkernen bleekblauw-wit is en met toenemende diepte overgaat in een verzadigd teal-indigo, terwijl de thermische vibraties bij −10°C de contouren van de verste molecuullagen oplossen in een lichtgevende turquoise waas. Men staat hier niet buiten de materie maar er middenin, omsloten door tienduizenden moleculen die in kristallografisch unisono trillen.

In een organisch kristal sta je aan de voet van een oneindig woud van opstijgende kolommen, elk opgebouwd uit coroneen-schijven van negen ångström doorsnede die met slechts 3,4 ångström tussenruimte op elkaar gestapeld zijn — een afstand niet groter dan de breedte van een enkele koolstofbinding. De aromatische kernen, elk gevormd door twaalf aaneengesmede benzeenringen, gloeien diep barnsteenkleurig doordat hun volledig gedelokaliseerde π-elektronenwolken zich uitstrekken over het volledige vlak van de schijf en bij elke stapelinterval samensmelten tot een gedeeld elektronisch membraan tussen twee buren. Dit π-stapelen is geen toevalligheid maar een kwantummechanische gedrevenheid: de overlappende π-orbitalen verlagen de totale energie van het systeem en maken tegelijk coherente ladingsoverdracht mogelijk langs de kolomas, zichtbaar als kortdurende elektrisch-oranje polaronflitsen die van schijf tot schijf springen als een stille bliksem in een amberen kathedraal. De ruimte tussen de kolommen is gevuld met een koele grijsblauwe waas van van-der-Waalswisselwerking — geen leegte, maar een zacht opalescent volume van zwakke, afstandsafhankelijke dipoolkrachten die het hexagonale kristalrooster bijeen houden zonder enige covalente binding.

Je bevindt je diep in het binnenste van een ribosomale uittunnnel, omsloten door gewelfd ribosomaal RNA dat als de wanden van een levende kathedraal om je heen kromt — de ribbelende dubbele helixen van rRNA-stelen vormen een nauwsluitend kanaal van nauwelijks tien ångström op de smalste punten, hun fosfaat-suikerruggengraten glanzend nat als gepolijst kobaltblauw steen. Op regelmatige intervallen vangen magnesiumionen het zwakke bioluminescente licht en kaatsen het terug als scherpe, geelwitte vonken, als klinknagels in een scheepsromp die de negatieve ladingen van het RNA neutraliseren en de architectuur van de tunnel in stand houden. Door het hart van dit beknellende kanaal kronkelt de nieuw gesynthetiseerde polypeptideketen — warm amber en gesiena, residue voor residue geboren uit het peptidyltransferasecentrum daarboven, met hier en daar de eerste glimpen van een alfa-helix die al begint te ontvouwen terwijl de keten voortglijdt. Aan de periferie, achter de innerlijke RNA-wand, flakkeren onregelmatig teal-blauwe lichtpulsen op en doven weer uit, GTP-hydrolyse-gebeurtenissen die zich manifesteren als vluchtige chemoluminescente flitsen voordat ze binnen femtoseconden oplossen in de dichte, solventgevulde duisternis. Alles hier beweegt — thermische trilling doordringt elke binding, elke knik van de keten, elk ångström van een ruimte die nooit stil is.
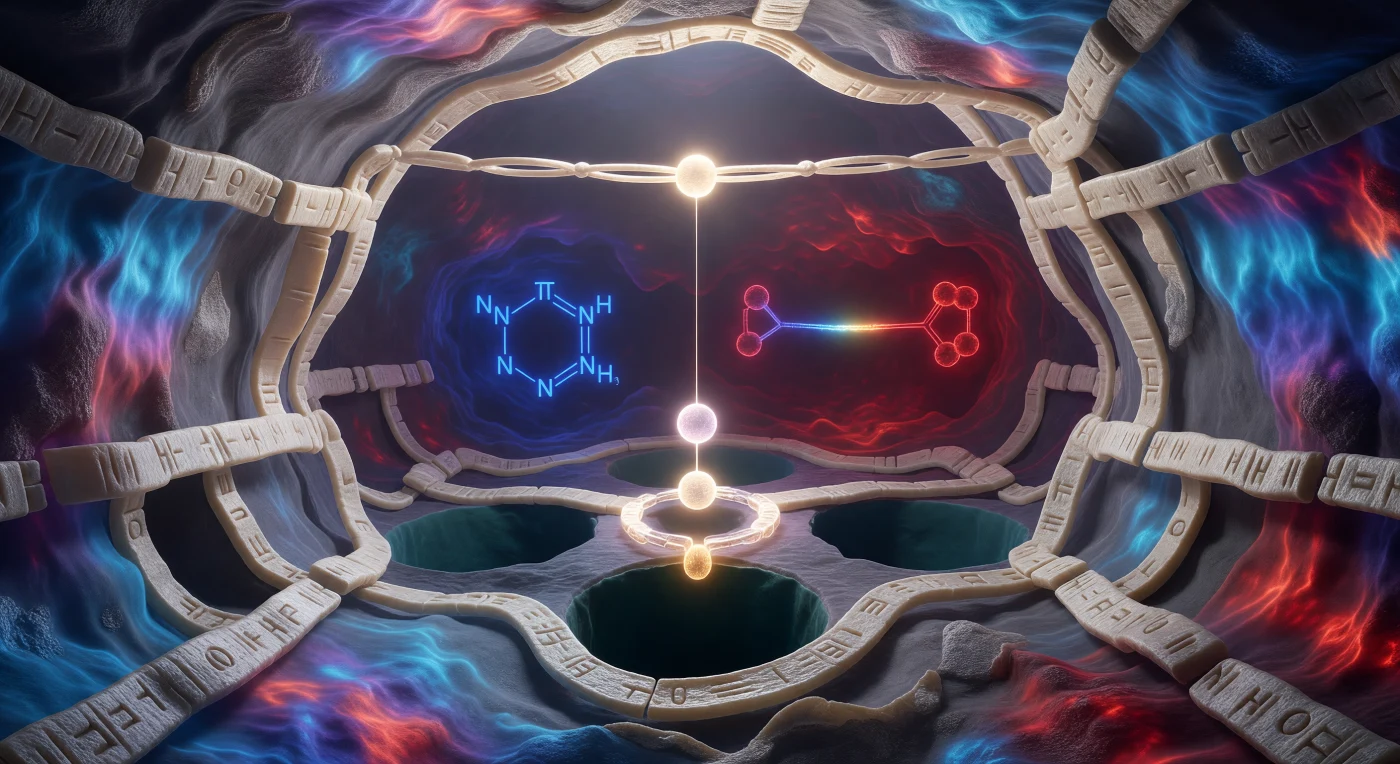
Je staat op de drempel van een moleculaire grot van slechts vijftien ångström breed, diep verscholen in het actieve centrum van trypsine, omgeven door wanden van beta-plaatststructuur die in ivoor en beenderenwit golfden als steen gehouwen uit een levend bouwwerk van eiwitketens. De elektrostatische potentiekaart die de holte verlicht, trekt een ononderbroken chromatische golf van koel kobaltblauw via violet naar diep scharlaken, want hier concentreren zich ladingen met een intensiteit die elk hoekje en elke atomaire uitstulping zichtbaar maakt — de ruwheid van van-der-Waalsradii is geen abstractie maar een tastbaar reliëf dat je van alle kanten omsluit. Midden in de holte zweeft de imidazolring van histidine als een elektrisch blauwe schijf, zijn π-elektronenwolk trillend boven en onder het vlak terwijl hij klaarstaat als protonshuttle tussen twee toestanden, en verderop ankert het donkerkarmozijnen carboxylaat van aspartaat de achterwand als het zwaartepunt van een katalytische driehoek die verbonden is door licht alleen. Recht boven je hoofd hangt de te knippen peptidebinding van het substraat over de holte als een lumineuze brug, precies drie ångström verwijderd van het wit-gouden zuurstofatoom van serine 195 dat zijn vrije elektronenparen uitstrekt in een spanning die onvermijdelijk aanvoelt — reactant en product gescheiden door niets dan elektrostatische adem en thermische trilling.

Op de rand van een enkel grafeen-vel staand, snijdt de wereld zich in twee absolute realiteiten: de zijwaartse blik onthult een donkere, chirurgisch scherpe lijn van slechts één atoom dik, een wiskundige grens die op de een of andere manier in materie is gekristalliseerd, terwijl langs die rand de elektronenwolken uitwaaieren in zachte amber-gouden halo's waar de periodieke roosterstructuur eindigt en vrije bindingen in het vacuüm reiken. Vanuit het bovenaanzicht explodeert het grafeen-vlak in een oneindig zilveren-blauw zeshoekig kettenhemd dat zich naar alle horizonten uitstrekt, waarbij de gedekolaliseerde π-elektronen een koude metallieke glans verspreiden die doet denken aan gepolijst platina. Lange, trage golfbewegingen — flexurale fononen met golflengte van 10 tot 100 nanometer — wentelen traag door het anders wiskundig perfecte oppervlak als een bevroren ademhaling over een stalmeer. Diep in het midden van dit kristallijne landschap gloeit een Stone-Wales-defect warm oranje: twee koolstofringen die door een lokale bindingsrotatie zijn omgezet tot een versmolten vijfhoek-zevenhoek-paar, een smeulende kool begraven in vorst, waarvan de gelokaliseerde elektronische toestand zich als amber uitwaait door een dozijn omringende ringen voordat het perfecte hexagonale symmetrie de koele blauwe roosterglans hervat.

Op een afstand van slechts 2,36 ångström van het Na⁺-ion hangt de waarnemer in het hart van een van de meest fundamentele ordenende krachten in waterige systemen: de hydratatieschil die elektrostatische lading omsluit en tempert. Het ion brandt in het midden als een gecomprimeerde goudwitte ster, geen warmtebron maar een bron van pure elektrostatische druk die het omringende medium naar zich toe buigt en elk molecuul binnen bereik in een ijzeren geometrische greep houdt. De zes watermoleculen van de eerste hydratatieschil hangen in een octaëdrische kooi met een precisie die in vloeibaar water elders vrijwel nooit bestaat — hun zuurstofzijden naar het ion gericht, blauwverschoven door de versterkte dipoolordening en de toegenomen waterstofbindingsstructuur die de elektrostatische omgeving afdwingt, hun waterstofuiteinden naar buiten wijzend als bruggen naar de losser georganiseerde tweede schil op circa 4,5 ångström. Voorbij die overgangszone, waar thermische beweging de elektrostatische orde begint te overheersen, lost het blauw op in het donkere, roerige netwerk van vrij bulk water — een woelig, flickerend landschap van moleculen die voortdurend van waterstofbindingspartner wisselen, zonder geheugen van de orde die zo dichtbij nog absoluut heerst.

Je bevindt je in het verstilde hart van een gevouwen eiwit, omsloten door een mozaïek van koolwaterstofketens en aromatische ringen die zo strak in elkaar grijpen dat elke richting binnen enkele ångströms op een gebogen moleculair oppervlak stuit. De ruimte gloeit in warme amber- en goudtinten: vertakte leucine- en isoleucinegroepen, hun methylgroepen elkander rakend via van-der-Waalskontakten van 3,5 tot 4,0 Å, vormen een driedimensionaal kristal van organische materie met een pakkingsdichtheid die die van geslepen edelsteen evenaart. Plotseling doorbreekt het diepblauwe schijnsel van een tryptofaan-indolring de warme amber-warmte — de π-elektronen van zijn bicyclisch aromatisch systeem stralen een turquoise aura uit over de naburige ketens — terwijl een begraven methioninesoufrgatom een scherpe goudgele glinstering terugkaatst als een metalen kraal in organisch barnsteen. Aan de verste rand van het waarneembare, op zo'n vijftien tot twintig ångströms afstand, begint de strakke pakking te verslappen en sijpelt een koele blauwgrijze nevel naar binnen: het water dat de hydrofiele buitenkant van het eiwit omspoelt, oneindig ver weg in deze wereld van moleculaire intimiteit.

Op het oppervlak van dit eiwitlandschap strekt de licht gebogen proteïnehuid van een icosaëdrisch plantvirus zich in alle richtingen uit als een kristallijn planetoïdeoppervlak, betegeld met vijfbladige pentamere rozetten en zeshoekige hexamere kronen waarvan elk bladblad een afzonderlijk manteleiwit-subunit is met zijn eigen reliëf van ruggen, zadelvormige valleien en golvende koepels. De concave receptor-bindingsholten tussen de rozetten kolen diepblauw en indigo op, geometrisch precies afgestemd om complementaire moleculaire partners op te vangen die elk moment uit het omringende oplosmiddel kunnen opdrijven, terwijl de immunodominante lusstructuren aan de pentamere hoekpunten oplichten in lumineus geelwit. In de smalle voegen tussen naburige subunits flikkeren zoutbruggen tussen tegengesteld geladen residuen in gepaard rood-blauw — aspartaat-zuurstofwolken in karmozijn, lysine-stikstofhalo's in koel kobalt — een elektrostatische spanning die het icosaëdrische rooster op zijn plaats houdt met een kracht die tastbaar maar onzichtbaar is. Over het gehele buitenoppervlak zweeft een geordende hydratatielaag van twee tot drie nanometer dik: watermolecule gebonden in quasi-kristallijne netwerken die elk contour van het eiwit getrouw volgen en het omgevingslicht verstrooien in opaalkleurige blauwwitte accenten langs elke kam en lus. Dit eiwitwereld ademt subtiel — thermische fluctuaties en micro-bewegingen van de subunits geven het landschap de geologische kalmte van een systeem dat enorme energie opslaat in perfecte, geduldige icosaëdrische symmetrie.

De kijker staat pal in de mond van een collageentrippelhelix en kijkt recht langs de centrale as naar binnen, waar drie ineen gevlochten polypeptideketens — barnsteengeel, oceaanblauw en verweerd jade — zich als reusachtige strengen in een kloksgewijze supercoil van de beschouwer weg kronkelen, hun oppervlakken glanzend als geslepen halfedelstenen belicht vanuit een bron die tegelijk van buiten en van binnenuit lijkt te komen. In het hart van deze moleculaire kathedraal raken de glycineresiduen van alle drie de ketens elkaar bijna aan met een onderlinge Cα–Cα-afstand van slechts 3,9 ångström, een tussenruimte nauwelijks breder dan één waterstofatoom, terwijl de pyrrolidinering van prolyl- en hydroxyprolineresiduen naar buiten steken en het oppervlak van elke streng een diep gecorrigeerde textuur geven die doet denken aan de schors van een eeuwenoude boom, teruggebracht tot de nanoschaal. Tussen de strengen weeft een secundair netwerk van goudomrande interketenwaterstofbruggen en koelblauwgroene hydroxyproline-waterbruggen — licht doorschijnend, richtingsgevoelig en taut — de drie helices zonder ooit te raken in een precise registratie bijeen, als een gebrandschilderd raam verlicht van achter. De helische herhaling van 8,7 ångström manifesteert zich als een visueel ritme: op regelmatige intervallen bevestigt de drievoudige rotatiesymmetrie zich opnieuw in een pols van uitgelijnde geometrie, als een kaleidoscoop bevroren midden in een draai. Dieper langs de as vervaagt de gevlochten structuur geleidelijk tot een warme, moleculaire nevel van barnsteenoranjeachtig gloed, een afstand die op deze schaal aanvoelt als het einde van een lange gang.

De blik valt regelrecht in de schacht van een amyloïde eiwitlezel, en wat zich ontvouwt is geen organisch bouwwerk maar een architectuur van onheilspellende machinemaat: laag na laag van parallelle β-strengen stapelen zich op met een tussenruimte van nauwelijks 4,7 ångström, een cadans zo precies dat hij niet gegroeid lijkt maar gefreesd. Gouden waterstofbruggen spannen zich als dwarsbalkjes loodrecht op de diepte, de chemische lijm die elke peptiderug aan haar buur klinkt en die samen een ladder vormen waarvan de sporten nooit ophouden. In het absolute centrum gaapt de steric-zipper-kern als een zwartbasalten naad: hier sluiten hydrofobe zijketens van leucine en isoleucine zo naadloos op hun spiegelbeeldpartners aan dat geen enkel watermolecuul binnendringt, en het moleculaire oppervlak voelt aan als gepolijst obsidiaan, koud en ondoordringbaar. Aan de buitenrand van het fibril omhult een halfgeordende schil solvatatiewater de polaire ruggengraat in een parelmoeren glinstering van blauwwit licht, waar vloeistof en geordende fase in picoseconden-evenwicht coëxisteren. Wat de geest ten diepste treft is de herhaling zelf: deze structuur bouwde zichzelf moleculaire laag voor moleculaire laag, strekte zich in beide richtingen uit met dezelfde meedogenloze geometrische trouw, en confronteert de toeschouwer met zelftemplaterende replicatie als landschap — biologische onvermijdelijkheid versteend in atoomnauwkeurige diepte.
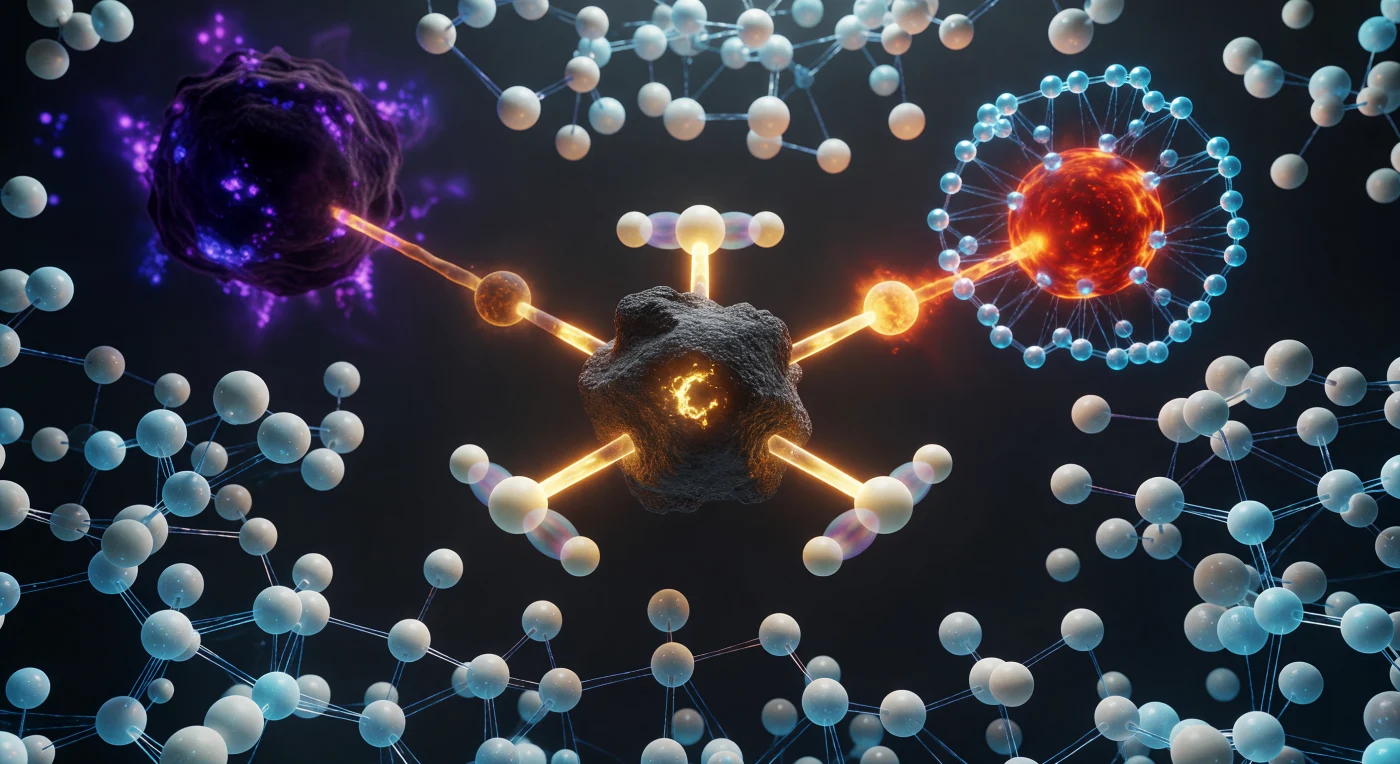
Op dit bevroren kwantummoment staart de kijker recht in het hart van een SN2-reactie: het centrale koolstofatoom hangt in onmogelijke pentacoördinatie, drie waterstofatomen vast in een perfect planair waaierpatroon van 120°, terwijl een rood-oranje zuurstofattoom aan één zijde nadert en een dieppaars broom aan de andere zijde langzaam loslaat — beide gedeeltelijke bindingen oplichtend in amber als half-gevormde beloften aan weerszijden van een lineaire O···C···Br-as. Dit is het overgangstoestandscomplex zoals de kwantumchemie het beschrijft: een energetisch maximum op de vrije-energieheuvel, een configuratie die nooit in rust bestaat maar slechts als vluchtig statistisch maximum in de faseruimte flakkert, met een levensduur van slechts enkele femtoseconden. De doorschijnende geestbeelden van de waterstofatomen — hun gespiegelde posities schemeren zichtbaar in kwantumsuperpositie — verbeelden de inversie van de valentiehoeken die kenmerkend is voor dit mechanisme, de moleculaire omkering die Walden al in 1896 beschreef. Rondom het reactiecentrum herordenen polaire watermoleculen zich voortdurend in concentrische cyaanblauwe hydratieschillen, hun waterstofbrugnetwerk zichtbaar als sidderende draden van licht die reageren op de herverdelende ladingen: een dichte, dynamische vloeistofwereld zonder horizon, waar elektrostatische krachten alles beheersen en thermische bewing van alle kanten indringt.

Je drijft door een elliptische tunnel die nauwelijks breder is dan één enkel molecuul, de wanden opgebouwd uit een eindeloos herhalend rooster van bleekgrijze siliciumknopen verbonden door felrode zuurstofattomen — Si-O-Si-bruggen die de buiswand een sinusoïdaal gegolfd reliëf geven, alsof de ribben van een kristallijne pijp uit atomaire bindingen zijn gesmeed in plaats van staal. Tien meter-ringen van onderling vergrendelde tetraëders verlichten zich vooruit als ijsblauw-witte decagonale bogen die in perfect perspectief samenkomen in een lichtend verdwijnpunt, terwijl felwitte punten op aluminiumsubstitutieplekken als microscopische lasvlammen gloeien — Brønsted-zuursites waar geconcentreerde reactiviteit in de verder stoichiometrisch serene mineraalwand is ingebrand. Dertig ångström verder opent een sinusoïdaal kanaal zich loodrecht in de linkerwand en vormt een moleculaire kruising van nog geen vijf ångström breed, een knooppunt waar gouden, ruimtevullende koolwaterstofmoleculen hun omhulsels in bijna perfecte van-der-Waals-contact tegen het zuurstofbeklede oppervlak drukken — elk molecuul past in zijn kristallijne gastheer als een sleutel in een slot. Dit is het hart van een ZSM-5-zeoliet, een alumino-silicaatrooster met een precisie die elke doorgang filtert op molecuulgrootte en -vorm, waardoor het wereldwijd wordt ingezet als katalysator bij de omzetting van methanol naar benzine en bij de kraakprocessen van de petrochemische industrie.
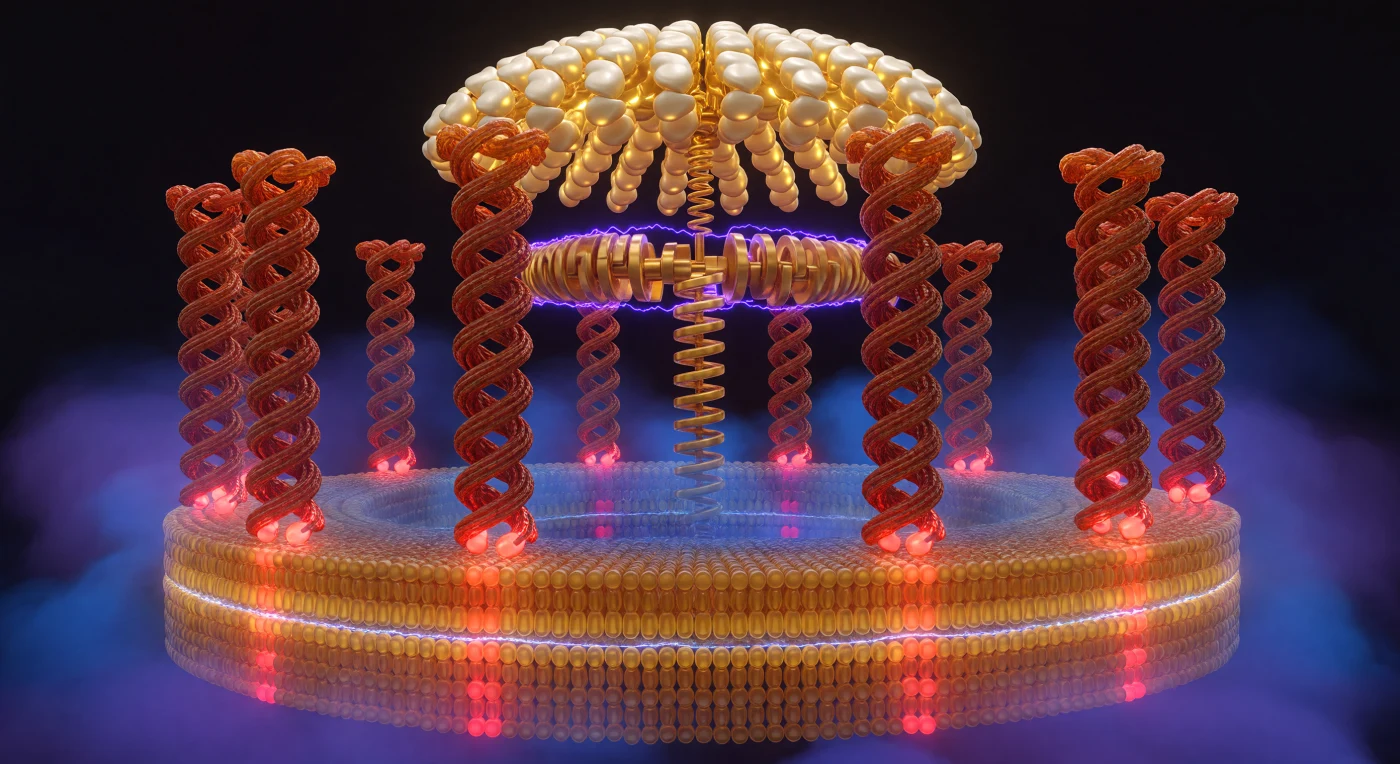
Je staat in het binnenste van een moleculaire machine die al vier miljard jaar zonder onderbreking draait: tien terracottakleurige α-helixkolommen welven zich rondom je als de muren van een levend colosseum, elk opgebouwd uit spiraalvormig gevlochten peptideketens waarvan de oppervlakken een zwak fosforescerende glans dragen, alsof nat zandsteen vuurlicht weerkaatst. Aan de voet van elke kolom brandt een enkel glutamaatresidu — felrood en elektrostatisch rauw waar protonen zich door het halvekanaaltje bewegen, gedempter baksteenkleurig waar de lading al geneutraliseerd is — terwijl dit pulserende contrast zich over een afstand van slechts enkele nanometer afspeelt, een ruimte die je met uitgestrekte armen zou kunnen overbruggen. De lipidenbilaag die deze cilindrische wereld omhult, tekent zich af als twee onderscheiden strata — warm barnsteengoud aan de buitenzijde, koeler blauwamber aan de binnenzijde — met op hun grens een iriserende naad zo scherp als een kustlijn, waar de rotorkolommen als tempelpilaren uit het membraan rijzen. Over deze grens heerst een electrochemisch spanningsveld van 150 tot 200 millivolt dat zich niet aandient via symbolen maar via het licht zelf: kobaltviolet en elektrisch blauw comprimeert zich aan de cytoplasmatische zijde met de gewelddadige dichtheid van opgehoopte protonen, terwijl de matrixzijde ademt in een rustiger middernachtsblauw dat langzaam oplost in de ruimte boven het membraan, waar de gouden F₁-katalytische koepel zich welft als een kathedraalgewelf en de gamma-subeenheid als een excentrische krukas de rotatiemomenten van dit protonenstroom omzet in de chemische energie die al het leven aandrijft.
