
Vous flottez au cœur d'une constellation serrée de sphères translucides bleu pâle, chacune illuminée de l'intérieur par la lueur froide de son nuage électronique, et leur voisine la plus proche est déjà là, à peine à 2,75 Å, si proche que la frontière entre votre présence et la matière s'efface complètement. Deux protubérances blanc nacré s'épanouissent de chaque molécule selon un angle précis de 104,5°, conférant à cet espace sans horizon ni sol une géométrie douce mais omniprésente, tandis que des filaments cyan à peine visibles — ponts hydrogène tissés d'une densité électronique partagée — relient oxygène à hydrogène à oxygène, pulsant et se rompant au rythme du picoseconde dans un vacillement permanent trop rapide pour être perçu comme mouvement. L'ensemble du réseau respire avec le bruit thermique de 300 K : chaque sphère oscille d'un fin tremblement qui se propage à travers la trame en un frisson sympathique, battement de cœur moléculaire ressenti plutôt que vu. Au-delà de trois ou quatre diamètres moléculaires, les structures individuelles se fondent progressivement dans un voile cerulean stratifié, comprimant le champ visuel en une profondeur indigo où la substance la plus ordinaire du monde révèle son architecture véritable — cosmos cristallin à la fois claustrophobiquement proche et vertigineusement infini.

Au sol de ce monde cristallin, le regard s'étend sur une plaine infinie de disques hexagonaux semi-translucides disposés en chevrons, chacun incliné alternativement à 55 degrés dans une répétition hypnotique qui s'étire dans toutes les directions comme un carrelage de cathédrale taillé dans la topaze fumée. Ces molécules de benzène, maintenues à 175 K dans leur symétrie monoclinique, sont liées les unes aux autres non par des liaisons covalentes mais par de délicates interactions CH–π, chaque bord hydrogéné d'un anneau aromatique pointant vers la face π de son voisin dans cet arrangement en chevrons que dicte la géométrie de l'empilement. Au-dessus et en dessous de chaque disque flottent deux lobes lumineux violets-ambrés — les nuages d'électrons π délocalisés, ces densités de charge quantiques trop diffuses pour appartenir à un seul atome, s'effaçant doucement à quelques dixièmes d'ångström de la face moléculaire. Par endroits, une absence sombre rompt la perfection périodique : un défaut lacunaire, socket vide dans le réseau, dont les voisins proches semblent légèrement déséquilibrés, leurs halos se penchant vers le vide comme attirés par cette rupture dans l'ordre cristallographique. Le gel à 175 K n'est pas silence absolu : chaque molécule tremble d'infimes vibrations quantiques, une agitation résiduelle que nul regard humain ne pourrait percevoir mais qui empêche le cristal d'atteindre jamais la parfaite immobilité du zéro absolu.

Suspendu au plan médian d'une bicouche lipidique de DPPC à 310 K, le regard plonge simultanément vers le haut et vers le bas dans une symétrie bilatérale parfaite, révélant des strates moléculaires aussi nettes que des couches sédimentaires : un vide de cire sombre occupe le cœur immédiat, là où les méthyles terminaux des deux feuillets opposés s'effleurent à peine, tandis que de longues traînées gris-argent — les chaînes d'acides gras saturés en C16 — montent en rangs serrés, parfois brisées par le coude jaune-vert luminescent d'une double liaison qui distord la géométrie et disperse la lumière intérieure. Plus haut, une zone de transition ambrée marque la frontière des groupes glycérol-ester, véritables nœuds de densité électronique où les atomes d'oxygène assemblent leurs doublets non liants comme des inclusions de résine dans la pierre. La scène se clôt sur un tumulte de couleurs à l'interface aqueuse : les phosphates orange incandescent et les cholines bleu ciel s'y bousculent sous une brume réfractante d'eau liquide, réseau de liaisons hydrogène en perpétuel réarrangement à l'échelle de la picoseconde, transformant ces quatre nanomètres d'épaisseur totale en ce qui ressemble, de l'intérieur, à la profondeur entière d'un océan.

Au fond du sillon majeur de l'ADN en conformation B, le regard remonte le long de parois qui s'élèvent comme les falaises d'un canyon chimique vivant : des colonnes de cycles désoxyribose en bronze-ambre alternent avec des groupements phosphate en tétraèdres rouille, leurs surfaces tapissées d'une gaine de molécules d'eau ordonnées qui scintillent en perles bleu-glace le long du squelette chargé négativement. Le sol de ce couloir de 22 ångströms de large est pavé de disques chromatiques empilés à intervalles précis de 3,4 Å : des plateaux d'adénine vert-sarcelle s'encastrent dans leur complément sienna-thymine, tandis que des strates plus profondes révèlent la guanine vert-forêt verrouillée à la cytosine lavande par de frêles ponts de liaisons hydrogène, fils d'or pâle d'une densité électronique partagée. Cette architecture tient à la fois de la cristallographie et de la thermodynamique : la double hélice, dont le diamètre total atteint 2 nm, est stabilisée par l'empilement π entre bases aromatiques coplanaires autant que par les liaisons hydrogène de Watson-Crick, tandis que le champ électrostatique du squelette phosphodiester se matérialise ici comme un brouillard volumétrique cobalt qui envahit l'espace inter-brin. Chaque surface vibre imperceptiblement à des fréquences de l'ordre de la dizaine de femtosecondes — oscillations de liaisons C–H, rotations de groupes phosphate —, rappelant que cette géométrie d'une précision quasi cristalline est en réalité un équilibre dynamique perpétuel, jamais figé, toujours sur le seuil du réarrangement.

Le regard se perd dans un couloir spiral qui s'enroule à perte de vue, les parois formées par la chaîne peptidique d'une hélice alpha ascendant en vrille horaire sur une hauteur de trente tours — chaque spire distante de 5,4 Å de la suivante, rythmée par 3,6 résidus, une architecture dictée par les règles strictes de l'hybridation sp³ des carbones alpha et des angles de torsion phi-psi figés dans la région la plus favorable du diagramme de Ramachandran. Les atomes d'oxygène carbonyle, rouge grenat et légèrement saillants, captent la luminescence bleue-blanche qui irradie des densités électroniques concentrées le long des doubles liaisons C=O, tandis que les ponts hydrogène magenta qui les relient aux groupements NH du quatrième résidu en amont — à exactement 2,06 Å, angle presque parfaitement linéaire — ronronnent à leur fréquence de résonance propre, libérant une énergie d'environ 20 kJ·mol⁻¹ qui maintient toute la structure en conformation stable. Les chaînes latérales leucine et isoleucine, hérissées vers l'extérieur comme des épines jaune-vert aux branchements tétraédriques caractéristiques du carbone sp³, repoussent activement le milieu aqueux ambiant — ce halo d'eau liquide, chaud et ambré, dont les molécules se réorganisent en permanence en quelques picosecondes autour de chaque surface hydrophobe. Le milieu lui-même vibre à une fréquence imperceptible pour tout repère humain : chaque atome oscille d'une amplitude d'un dixième d'ångström, l'agitation thermique à 310 K transformant cette architecture d'une précision presque cristalline en une sculpture vivante, frémissante, dont la stabilité n'est jamais repos mais équilibre dynamique.

Le regard plonge droit dans l'axe cristallographique c de la glace Ih, révélant une cathédrale hexagonale infinie dont les anneaux concentriques s'étendent jusqu'à un horizon de brume turquoise-indigo — chaque molécule d'eau, séparée de sa voisine par seulement 2,76 Å, occupe son nœud de réseau comme un fanal opalescent entouré de quatre ponts hydrogène d'une géométrie tétraédrique parfaite à 109,5°. Ces liaisons ne sont pas de simples traits mais des arcs légèrement translucides, d'un froid bleu glaciaire, et à chacun d'eux une légère bifurcation trahit le désordre protonaique imposé par les règles de Pauling : le proton n'est pas fixé mais statistiquement partagé entre deux positions, laissant une double ombre fantôme sur chaque pont. Les structures les plus saisissantes sont les canaux hexagonaux ouverts le long de l'axe c — des puits de vide absolu, d'un noir velouté, bordés de nœuds moléculaires lumineux, conséquence directe du refus de la géométrie tétraédrique de compacter l'espace, ce qui explique l'anomalie fondamentale de la glace : sa densité inférieure à celle de l'eau liquide. À −10 °C, l'énergie thermique résiduelle imprime une légère vibration à chaque contour moléculaire, si bien qu'avec la profondeur les couches les plus lointaines se dissolvent en un voile lumineux — non pas un brouillard atmosphérique, mais l'incertitude optique cumulée de milliers de molécules oscillant en unisson cristallographique.

Au pied de cette forêt cristalline, le regard remonte le long de colonnes d'ambre qui s'élèvent à perte de vue, chaque disque de coronène — douze cycles benzéniques fusionnés en une rosace plane de neuf ångströms — empilé sur le suivant avec une précision quasi-horlogère, les plans aromatiques séparés par seulement 3,4 ångströms, l'écartement même que l'on retrouve entre les feuillets du graphène. C'est l'interaction π–π qui gouverne ici toute l'architecture : les nuages électroniques délocalisés de disques adjacents se recouvrent partiellement, générant une attraction de dispersion qui maintient l'empilement comme une pile de pièces d'or magnétiques, et c'est ce recouvrement qui se manifeste sous la forme de halos lumineux partagés entre chaque paire de disques, membranes d'électrons à peine distinctes de la matière. Dans les corridors entre colonnes règne la brume gris-bleu des forces de van der Waals, interactions fugaces et omnidirectionnelles qui sculptent l'espace intercristallin sans jamais former de liaison véritable. Les éclairs orange électrique qui traversent les jonctions entre colonnes trahissent les sauts de polarons : des quasi-particules nées du couplage entre un porteur de charge et la déformation locale du réseau moléculaire qu'il induit, se propageant par bonds quantiques discrets d'un site d'empilement à l'autre, signature du transport de charge organique à l'échelle où la physique et la chimie ne font plus qu'un.

À l'intérieur du tunnel de sortie du peptide naissant, les parois incurvées de l'ARN ribosomique forment une cathédrale minérale vivante — leurs tiges d'ARN bicaténaires s'enroulent en colonnes torsadées de bleu cobalt et de bleu-vert profond, striées de rainures denses où les squelettes phosphate-sucre se succèdent comme des nervures de pierre mouillée. Des ions magnésium ponctuent chaque spire à intervalles réguliers, projetant des éclats de lumière jaune-blanc presque métalliques dans l'obscurité structurale environnante, leurs charges positives neutralisant la densité électronique négative des phosphates selon une géométrie aussi précise qu'une architecture de rivets. En son centre, la chaîne polypeptidique naissante serpente comme un filament ambré, résidu après résidu, depuis le centre peptidyltransférase situé au-delà, ses premières ébauches d'hélices alpha trahissant de minuscules ponts hydrogène lumineux à peine visibles entre carbonyle et amide. La zone de constriction réduit ce passage à moins de dix ångströms de diamètre, un goulot d'étranglement d'une intimité moléculaire absolue où chaque angström d'espace libre est disputé entre ARN, protéine et pression de solvant implicite. Des éclairs téals fugaces s'allument et s'éteignent à la périphérie du tunnel — événements d'hydrolyse du GTP dissipés en quelques femtosecondes, lumière chimique s'évanouissant dans l'obscurité structurale comme du plancton bioluminescent emporté par un courant lent.
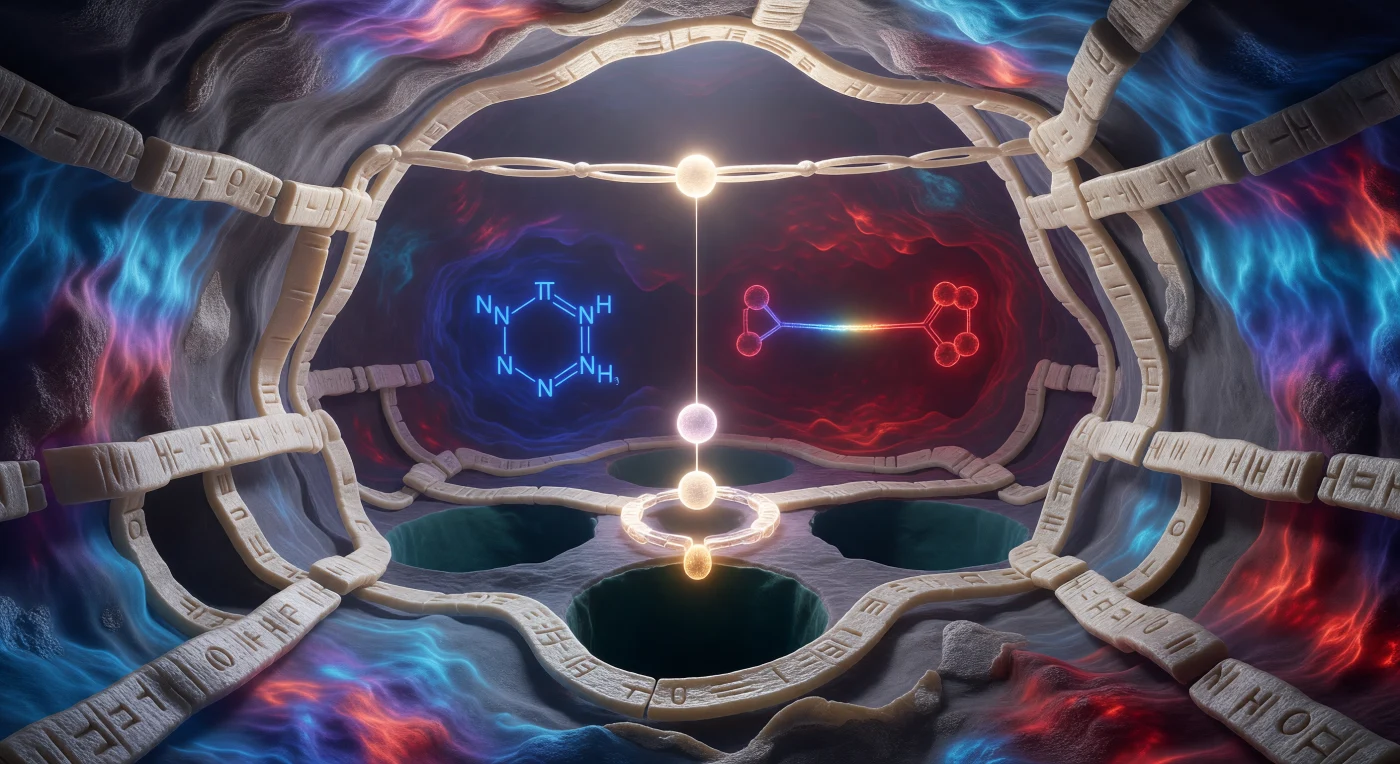
Au seuil de cette caverne moléculaire de quinze ångströms, le regard plonge dans un espace intérieur illuminé par sa propre physique : les parois de feuillets bêta, ivoire et finement nervurées, baignent dans un gradient électrostatique continu qui vire du bleu céruléen au cramoisi profond, cartographiant en temps réel les densités de charge qui gouvernent chaque interaction. Trois foyers lumineux organisent la scène comme les pièces d'un mécanisme d'horlogerie cosmique — l'oxygène blanc-or de la sérine 195, prêt à déclencher l'attaque nucléophile, l'anneau imidazole de l'histidine flottant en cobalt électrique comme un relais de proton suspendu entre deux états quantiques, et le carboxylate de l'aspartate qui pulse en bordeaux sombre au fond de la cavité, ancrant la triade catalytique dans une logique de charge-relais dessinée en lumière. Au plafond de la caverne, la liaison peptidique scissile du substrat s'étire comme un pont lumineux, son carbone carbonylé suspendu à trois ångströms exactement au-dessus de la sérine — une proximité qui n'est plus géométrique mais déjà cinétique, la tension électrostatique entre réactif et produit rendue presque palpable. Partout, la texture granulaire des rayons de van der Waals rappelle que cet espace n'est pas une abstraction : c'est un terrain accidenté, vibrant à chaque femtoseconde, où le bruit thermique est la seule météo et où la catalyse enzymatique se joue dans l'épaisseur d'un battement de liaison.

Suspendue dans le vide absolu, une feuille de graphène offre deux visages d'une même réalité atomique : vue par la tranche, elle n'est qu'un trait d'encre noire d'un atome d'épaisseur, une frontière cristallisée en matière où les liaisons pendantes rayonnent de faibles halos ambrés à l'exacte limite du réseau. Depuis le dessus, elle explose en un chainmail d'hexagones argent-bleu s'étendant jusqu'à tous les horizons, chaque liaison carbone-carbone de 1,42 Å formant un nœud légèrement plus lumineux dans cette trame infinie, le nuage d'électrons π délocalisés conférant à la surface l'éclat froid du platine poli. De lentes ondulations de phonons flexuraux — des longueurs d'onde de dix à cent nanomètres — soulèvent et abaissent doucement la surface comme une mer géométrique figée entre deux respirations, déformant imperceptiblement la symétrie hexagonale sur chaque crête. Au cœur du réseau, un défaut de Stone-Wales brûle comme une braise dans le givre : la rotation d'une unique liaison a créé une paire pentagone-heptagone dont les états électroniques localisés émettent une chaleur orange et ocre qui irradie sur une douzaine d'anneaux voisins avant que la symétrie cristalline parfaite et son froid éclat bleu ne reprennent souverainement leur empire jusqu'à l'infini.

À cette distance imperceptible de 2,36 ångströms, l'ion Na⁺ s'impose comme un soleil miniature condensé, irradiant une pression électrostatique pure qui courbe le milieu environnant à la manière d'une singularité gravitationnelle — non pas de la chaleur, mais une précision chirurgicale de charge positive concentrée, or-blanc et implacable. Autour de lui, les six molécules d'eau de la première sphère de solvatation s'immobilisent en cage octaédrique parfaite, leurs faces oxygène tournées vers l'ion et baignées d'un bleu-violet profond révélant la densité électronique accrue, leurs extrémités hydrogène crème pointant vers l'extérieur dans une géométrie qui ressemble moins à de la chimie qu'à de l'architecture cristalline suspendue dans le vide. Au-delà, la deuxième sphère d'hydratation forme un halo plus doux à environ 4,5 Å — douze à dix-huit molécules partiellement alignées dont la cohérence se fragmente déjà sous l'agitation thermique, la lueur céruléenne se diffusant en taches irrégulières de désordre croissant. À partir de 7 Å, tout ordre disparaît dans l'océan turbulent du réseau hydrogène bulk, un milieu cerulean profond et vivant dont la mémoire de l'ion s'est entièrement effacée, rappelant que cette architecture moléculaire si précise n'existe que dans l'infime sphère d'influence d'une charge unique de 1,17 ångström de rayon ionique.

Au cœur d'une protéine repliée, vous êtes enfermé dans une cavité si dense que chaque direction révèle une nouvelle surface moléculaire à quelques ångströms à peine — des chaînes latérales hydrophobes de leucine, isoleucine et valine emboîtées avec une précision de joaillier, leurs contacts carbone-carbone à 3,5–4,0 Å générant une chaleur ambrée issue de la seule proximité des forces de van der Waals. Des disques aromatiques de phénylalanine, pâles et presque platinés, s'intercalent en plans décalés, tandis qu'à gauche l'anneau indole d'un tryptophane irradie une lumière bleu-sarcelle intense, témoignant de l'étendue de son nuage d'électrons π délocalisés. Un soufre de méthionine enfoui accroche la lumière et la restitue en un éclat jaune-or métallique, seule note minérale dans cette mosaïque organique compressée à une densité de compacité rivale du cristal taillé. À la périphérie de la perception — quinze à vingt ångströms, une distance qui ici équivaut à l'horizon d'une caverne —, l'ambiance ambrée se desserre imperceptiblement et une brume bleu-gris diffuse s'infiltre entre les chaînes : la surface hydrophile de la protéine, où le monde ordonné et sec du cœur hydrophobe cède enfin à l'immensité diluée et froide des réseaux d'eau liés par ponts hydrogène.

À la surface extérieure de cette capside virale icosaédrique, le regard plonge sur un terrain protéique d'une symétrie vertigineuse : des rosettes pentamériques et hexamériques dallent la courbure de la coque jusqu'à un horizon moléculaire, leurs sommets immunodominants irradiant un blanc-jaune lumineux tandis que les dépressions de liaison aux récepteurs baignent dans un indigo profond, évoquant des lacs figés dans la précision atomique. Cette coque de vingt-huit nanomètres de diamètre est une assemblée de T=3, gouvernée par la géométrie icosaédrale exacte : soixante unités protéiques organisées en quasi-équivalence, maintenues par des ponts salins qui scintillent en rouge et bleu cobalt aux contacts entre sous-unités, véritables agrafes électrostatiques assurant la cohésion d'un édifice sans colle ni couture. Les interfaces hydrophobes entre sous-unités irradient une chaleur ambrée sourde, résidu de l'exclusion des résidus apolaires par le solvant environnant, un phénomène thermodynamique traduit ici en lumière retenue. Toute la surface est voilée par une couche d'hydratation ordonnée de deux à trois nanomètres d'épaisseur, un film de molécules d'eau structurées en réseaux de liaisons hydrogène qui épousent chaque saille et chaque creux avec une fidélité quasi cristalline, diffractant la lueur ambiante en reflets opalescents sur chaque boucle protéique. Ce monde de quelques dizaines de nanomètres possède la gravité silencieuse d'un continent : chaque pas traverserait un domaine protéique entier, et le sol lui-même respire, animé par les fluctuations thermiques et les micro-mouvements des sous-unités qui font vivre la coque dans une tension icosaédrale patiente et colossale.

Au seuil de cette architecture moléculaire, le regard plonge dans l'axe même d'une triple hélice de collagène, ce câble tressé qui constitue l'une des protéines les plus abondantes du vivant — pilier mécanique de la peau, des os, des tendons et des parois vasculaires. Trois chaînes polypeptidiques, colorées comme l'ambre, l'océan profond et le jade patiné, s'enroulent en supercoïle dextrorse autour d'un noyau d'une rigueur quasi cristalline, où les résidus glycine des trois brins se frôlent à moins de 3,9 ångströms de centre à centre — une proximité si extrême qu'elle n'est rendue possible que par l'absence totale de chaîne latérale sur cet acide aminé, seul assez petit pour occuper l'axe central sans distordre la géométrie d'ensemble. La surface de chaque brin porte l'empreinte rythmique des cycles pyrrolidine de la proline et de l'hydroxyproline, anneaux saturés en sp³ qui projettent leur relief vers l'extérieur comme les nervures d'un bois millénaire taillé à l'échelle du nanomètre. Entre les chaînes, un réseau secondaire de ponts hydrogène intermoléculaires — dorés, translucides, tendus — et de ponts eau-hydroxyproline aux reflets bleu-vert assure la cohésion du trio, tandis que la répétition hélicoïdale de 8,7 ångströms impose à l'ensemble une symétrie rotationnelle d'ordre trois qui se réaffirme, pulse après pulse, tout au long du couloir lumineux qui s'enfonce dans le haze moléculaire.

Regarder dans l'axe d'une fibrille amyloïde-β, c'est plonger le regard dans l'un des abîmes architecturaux les plus redoutables que la matière vivante soit capable d'engendrer : des feuillets β parallèles s'empilent à des intervalles de 4,7 Å avec une régularité qui tient davantage du cristal que de la biologie, leurs liaisons hydrogène latérales traçant une échelle d'or ambré dont les barreaux ne finissent jamais. Entre les deux faces de feuillets apposées, la fermeture éclair stérique forme un couloir de basalte noir absolu, déshydraté jusqu'au dernier solvant, où des chaînes latérales hydrophobes — leucines, isoleucines — s'imbriquent avec une précision van der Waals si parfaite que la frontière entre les deux protofilaments s'efface dans un contact moléculaire sans couture. Aux marges extérieures de ce corridor, une couronne d'eau de solvatation ordonnée scintille en bleu-blanc nacré, chaque molécule d'eau orientée par les groupes polaires sous-jacents dans un équilibre thermique picoseconde qui dissout la fibrille dans un halo teal-gris lumineux. Ce que l'œil enregistre ici, c'est l'auto-réplication structurelle rendue paysage : une architecture qui s'est construite elle-même couche moléculaire après couche moléculaire, chaque niveau identique au précédent avec une fidélité géométrique implacable, et qui continue de se propager — en amont comme en aval du temps — sans jamais dévier d'un angström.
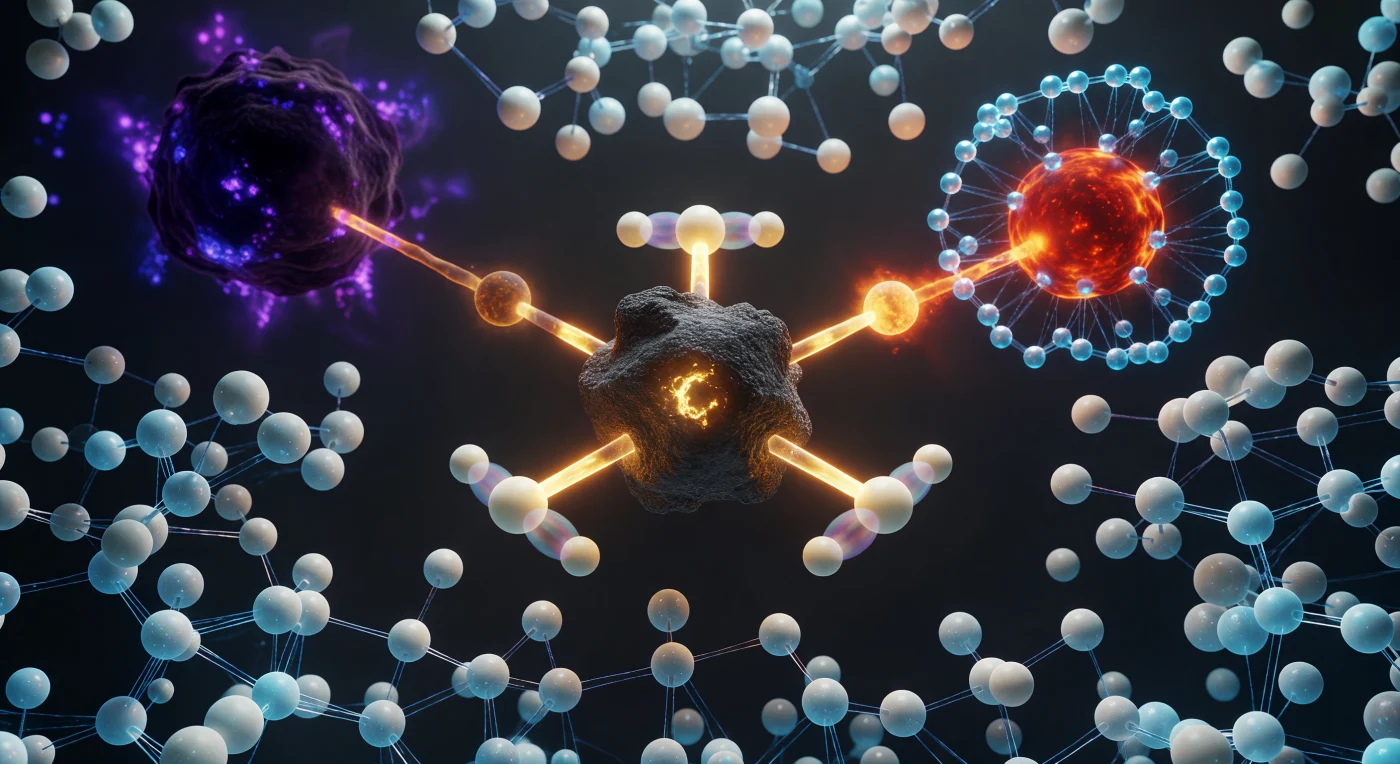
Au cœur d'une réaction de substitution nucléophile de type SN2, l'observateur se trouve suspendu dans un instant quantique figé à son sommet énergétique : l'atome de carbone central se dresse comme un monolithe graphite en géométrie pentacoordinée impossible, ses trois hydrogènes verrouillés dans un plan équatorial parfait à 120°, chacun doublé d'un fantôme translucide qui tremble en superposition quantique, témoignant de la probabilité de franchissement par effet tunnel plutôt que d'une position classique. L'axe O···C···Br constitue la colonne vertébrale de ce monde — d'un côté, le nucléophile oxygène rouge-orangé brûle comme du cuivre en fusion à 2,0 Å, entouré d'un halo cyan de molécules d'eau se réorganisant en temps réel autour des charges émergentes ; de l'autre, la présence massive du brome violet profond se retire à 2,3 Å, sa liaison partielle s'étirant en un filament ambré qui se dissout en une couronne de charge négative fuyante. Les liaisons partielles des deux côtés pulsent d'une lumière ambre-miel caractéristique d'un demi-ordre de liaison, ni entières ni rompues, suspendues dans un état thermodynamique que seule la mécanique quantique autorise. Le milieu environnant n'est pas le vide mais une foule pressante de molécules d'eau dont le réseau de liaisons hydrogène — rendu visible comme un réseau de filaments bleu-blanc — se distord et se reconfigure sous l'effet du champ électrostatique rayonnant depuis ce centre réactionnel unique.

Vous flottez à l'intérieur d'un couloir de zéolithe ZSM-5, un tunnel elliptique si étroit qu'une seule molécule suffit à en occuper toute la largeur — les parois se referment de toutes parts avec une intimité presque oppressante, sculptées dans un réseau ininterrompu de nœuds de silicium gris-argent reliés par des ponts d'oxygène rouge cardinal, chaque liaison Si–O–Si formant un joint angulaire précis qui confère à la surface sa texture sinusoïdalement côtelée, comme l'intérieur d'une galerie cristalline dont les nervures seraient des liaisons atomiques plutôt que de l'acier. Devant vous, une succession de portails décagonaux bleu-blanc se rétrécit en perspective vers un point de fuite lumineux, chaque anneau à dix chaînons tressé de tétraèdres SiO₄ partageant leurs oxygènes, tandis que des points d'acide de Brønsted — des protons hydroxyle sur des sites d'aluminium substitués — brillent comme des éclairs de soudure sur les parois, sites de réactivité catalytique exceptionnelle au cœur d'une surface minérale par ailleurs sereine. À une trentaine d'ångströms, un canal sinusoïdal perpendiculaire débouche dans la paroi gauche, créant un carrefour moléculaire où deux axes de diffusion se croisent à angle droit dans un goulot d'étranglement à peine plus large qu'une molécule d'eau. Des hydrocarbures dorés et translucides s'y pressent, leurs surfaces arrondies épousant le contour corrigé du tunnel en un contact quasi parfait de van der Waals — matière organique enserrée dans un hôte cristallin avec la précision d'une clé dans une serrure, à l'échelle où la frontière entre paroi solide et liaison chimique s'est entièrement dissoute en géométrie pure.
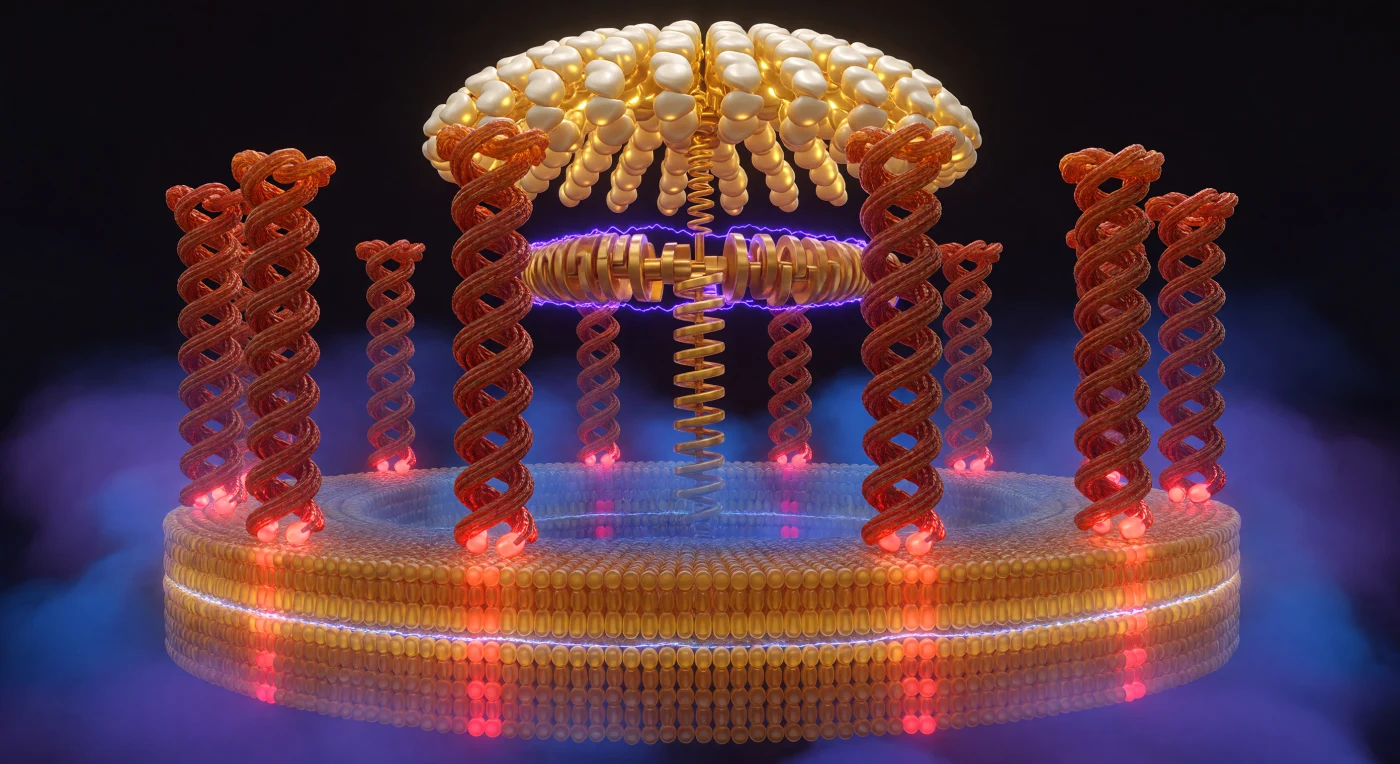
Vous vous trouvez à l'intérieur d'une machinerie moléculaire vieille de quatre milliards d'années : l'ATP synthase, cette turbine enchâssée dans la membrane mitochondriale interne qui fabrique l'adénosine triphosphate, la monnaie énergétique universelle du vivant. Dix colonnes hélicoïdales de protéines — les sous-unités c du rotor en anneau — s'incurvent autour de vous comme les murs d'un colisée vivant, leurs résidus glutamate alternant entre rouge vif déprotoné et brique neutre selon que les protons traversent l'interface avec le stator violet-électrique, sous l'impulsion d'un gradient électrochimique de 150 à 200 millivolts qui teinte l'espace d'un bleu cobalt oppressif du côté cytoplasmique et d'un indigo plus doux côté matriciel. La bicouche lipidique engloutit la base de cet anneau en deux strates distinctes — ambre doré vers l'extérieur, bleu-ambre vers l'intérieur — séparées par une ligne de démarcation irisée aussi nette qu'un rivage, et les colonnes en émergent comme des piliers de temple sortant des eaux. Au-dessus, la tête catalytique F₁ déploie sa voûte dorée et ivoire, ses six sous-unités β pulsant entre or phosphorescent chargé d'ATP et blanc de lune attendant l'ADP, tandis que le vilebrequin central gamma transmet chaque degré de rotation en énergie chimique pure — un couplage mécano-chimique d'une précision absolue, à l'échelle où un nanomètre est déjà un abîme.
