Confianza científica: Muy alto

Te encuentras suspendido en el interior de una red infinita y bulliciosa de moléculas de agua a 300 K, rodeado en todas las direcciones por esferas traslúcidas de un azul pálido —los núcleos de oxígeno— separadas entre sí por apenas 2,75 Å, una proximidad tan extrema que la noción de espacio vacío pierde todo sentido. De cada esfera emergen dos protuberancias blancas y densas formando un ángulo de 104,5°, y entre moléculas vecinas parpadean puentes cianóticos de densidad electrónica compartida —los enlaces de hidrógeno— que se forman y se rompen cada picosegundo con un temblor demasiado veloz para percibirse como movimiento, registrado en cambio como un destello suave y continuo. Todo el entramado vibra con ruido térmico: cada esfera oscila con una agitación fina y rápida que se propaga a través de la red como un latido molecular viviente, una resonancia simpática que se siente más que se ve. Más allá de tres o cuatro diámetros moleculares, las esferas individuales se disuelven en una neblina volumétrica de índigo profundo, comprimiendo el campo visual hasta un puñado de moléculas nítidas antes de que el mundo entero se funda en una profundidad translúcida. No hay suelo, ni cielo, ni gravedad que oriente la mirada: solo esta red isótropa, respirante y parpadeante, extendiéndose de manera idéntica en todas las direcciones —la sustancia más común del mundo revelada como un cosmos cristalino y alienígena allí donde su arquitectura verdadera se convierte en la única realidad.

El suelo se extiende en todas direcciones como un pavimento infinito de hexágonos translúcidos, cada disco de benceno inclinado en ángulos alternos de 55 grados que crean un tejido de espiga hipnótico y perfectamente periódico, como baldosas de ámbar ahumado dispuestas por una inteligencia geométrica sin fisuras. Cada molécula reposa sostenida en su lugar no por gravedad sino por la suma silenciosa de interacciones CH–π, esos abrazos débiles pero precisos entre el borde de hidrógeno de un anillo y la nube π de su vecino, una arquitectura de fuerzas fantasmas que mantiene el cristal monoclínico en su exactitud a 175 K. Sobre y bajo cada disco flota un halo bilobulado de luz violeta-ámbar —la nube de electrones π delocalizados, extendida a apenas un ångström de cada cara molecular— que se funde suavemente con los halos vecinos en los estrechos corredores de 3,5 Å entre planos aromáticos, llenando la perspectiva con una neblina luminosa y cálida que se pierde en lo que parece el equivalente de varias manzanas de ciudad. Aquí y allá, una oscuridad abrupta interrumpe el tapiz: una vacancia cristalográfica, un disco ausente cuyo hueco hace que los vecinos inclinen sus halos levemente hacia el vacío, como si la perfección periódica reconociera, sin poder remediar, su única grieta.

De pie en el plano medio absoluto de una bicapa lipídica de DPPC a 37 °C, el observador se encuentra suspendido en un interior ceroso y penumbroso —un vacío carbón surcado por estrías gris-plateadas que ascienden en ambas direcciones como columnas de una catedral subterránea: son las largas cadenas de ácidos grasos saturados, dieciséis a dieciocho carbonos de longitud, empaquetadas en rangos paralelos y trémulas con el calor térmico de cada picosegundo que pasa. De vez en cuando, un codo de luminiscencia amarillo-verdosa interrumpe la monotonía gris —el doble enlace insaturado de una cadena torcida, cuya geometría sp² desplaza la linealidad y dispersa la escasa luz interior de un modo que las cadenas rectas no pueden hacer. Ascendiendo a través de este bosque hidrofóbico, el espacio se transforma en estratos geológicos sucesivos: primero la zona ámbar translúcida de los gliceroles y sus ésteres, nódulos vítreos donde los oxígenos carboxilo acumulan densidad electrónica como resina en piedra; luego la capa de los grupos fosfato, esferas de un naranja profundo y electroestáticamente cargadas, bordeadas por las colinas azul-cielo que flotan en un halo acuoso centelleante donde cada red de puentes de hidrógeno nace y se rompe en decenas de picosegundos. El grosor total de este universo —apenas cuatro nanómetros de extremo a extremo— se experimenta desde dentro como la profundidad abismal de un océano, una escala íntima y aplastante donde la gravedad no existe y la distancia entre vecinos moleculares es tan pequeña como el diámetro de un solo átomo de carbono.

Ante tus ojos se despliega un cañón vivo tallado en la misma sustancia de la herencia biológica: las paredes que te rodean son columnas de anillos de desoxirribosa —pentagonos de bronce ambarino que se repiten en arcos ascendentes— interrumpidas por grupos fosfato que sobresalen como linternas tetraédricas de óxido translúcido, su superficie tapizada por moléculas de agua ordenadas en cadenas luminiscentes de color azul perla, cada una anclada por puentes de hidrógeno a los oxígenos del esqueleto. El campo electrostático negativo del esqueleto azúcar-fosfato se hace visible como una niebla cobalto que espesa el aire entre ambas paredes, comprimiendo el cielo a una franja de apenas veintidós ångströms de anchura donde la atmósfera acuosa vibra con una carga densa y húmeda. Bajo tus pies, el suelo del surco mayor es una estratigrafía de discos de pares de bases apilados con una precisión de 3,4 Å entre capa y capa: adenina azul-verdosa frente a timina ocre-sienna, guanina verde bosque frente a citosina lavanda, sus interfaces de puentes de hidrógeno brillando como hilos dorados de densidad electrónica compartida. Todo vibra con oscilaciones térmicas de femtosegundos que hacen que la geometría sólida parezca provisional, como si la molécula entera pudiera reconfigurarse antes de que tu percepción alcance a registrarlo —porque aquí, a esta escala, el movimiento y la estructura son la misma cosa.

El observador flota en el eje central de una hélice alfa proteica, envuelto por una espiral de átomos del esqueleto peptídico que asciende en sentido horario como una escalera de caracol de proporciones catedralicias, cada vuelta con un diámetro equivalente a tres alturas humanas y el corredor extendiéndose treinta giros hacia adelante antes de perderse en una neblina áurea de moléculas de agua. Los átomos de oxígeno carbonílico —rojos como granates pulidos— sobresalen a intervalos regulares captando la luminiscencia azul-blanca que emana de la propia densidad electrónica concentrada en cada enlace covalente, mientras que los grupos NH de nitrógeno miran hacia el interior del túnel en una alternancia estructural que define la geometría exacta de 3,6 residuos por vuelta y un paso helicoidal de 5,4 ångströms. Cada cuarto residuo, un puente de hidrógeno magenta de apenas 2,06 Å de longitud conecta un NH con el carbonilo cuatro posiciones atrás, estos arcos rosados que desprenden una tenue luminescencia como si la energía electrostática almacenada en ellos se disipara lentamente como luz, manteniendo la arquitectura secundaria rígida frente al bombardeo térmico constante del solvente. Las cadenas laterales hidrofóbicas de leucina e isoleucina se proyectan radialmente como espinas cristalinas amarillo-verdes de geometría tetraédrica perfecta, repeliendo el medio acuoso y dejando un halo seco en torno a cada masa no polar, mientras en los pliegues entre ellas la densidad electrónica cae casi a cero y las sombras se vuelven índigo profundo, un recordatorio de que en esta escala la materia es en gran medida vacío iluminado por probabilidad cuántica.

El observador flota suspendido en el corazón de una catedral hexagonal infinita, mirando directamente hacia abajo a lo largo del eje cristalográfico c del hielo Ih a −10 °C, donde anillo tras anillo de moléculas de agua se extienden en una perspectiva perfecta hasta disolverse en una neblina turquesa luminosa. Cada molécula ocupa su punto de red como un nodo opalescente y esmerilado, anclada por cuatro puentes de hidrógeno de 2,76 Å que se irradian hacia sus vecinas más cercanas con una geometría tetraédrica impecable de 109,5°, mientras una bifurcación fantasmal en cada enlace delata el desorden protónico descrito por las reglas de Pauling: el protón no reside en un único lugar sino en una superposición de posiciones levemente desplazadas, una indeterminación cuántica hecha visible. Los rasgos más dominantes de este paisaje son los canales hexagonales que se abren a lo largo del eje c como gargantas de vacío absoluto, pozos de oscuridad aterciopelada bordeados por los nodos moleculares resplandecientes —una consecuencia directa de que la geometría tetraédrica del enlace se niega a empaquetar de manera compacta, y es precisamente ese vacío estructural lo que explica la anomalía de la baja densidad del hielo frente al agua líquida. La luz parece emanar del propio retículo, más brillante en los núcleos de oxígeno donde se concentra la densidad electrónica y progresivamente más profunda en tonos teal-índigo a medida que la distancia acumula incertidumbre térmica, traduciendo en imagen la energía vibracional que a −10 °C aún sacude cada átomo con amplitudes del orden de décimas de ángström.

En el corazón de un cristal orgánico, el observador se encuentra al pie de una columnata infinita de discos de coroneno apilados, cada uno un anillo aromático de doce hexágonos fusionados que emite una luz ámbar profunda como resina calentada desde dentro, con apenas 3,4 ångströms separando cada disco del siguiente, una distancia casi idéntica al espaciado entre capas del grafito. Esta arquitectura no es arbitraria: la superposición de las nubes π de electrones deslocalizados entre discos adyacentes crea una membrana electrónica continua y luminosa, visible como un halo dorado que sangra suavemente en cada intersticio, manifestación directa de las fuerzas de apilamiento π-π que estabilizan el cristal y le confieren sus propiedades semiconductoras. A lo largo de toda la matriz hexagonal, el espacio entre columnas no está vacío sino habitado por una niebla gris-azulada y traslúcida, el rastro visual de las interacciones de van der Waals, fuerzas débiles pero omnipresentes que gobiernan a esta escala donde la gravedad no tiene ningún papel. Y en ese silencio cristalino irrumpen destellos anaranjados eléctricos, fugaces como el chasquido de un arco voltaico: polarones en tránsito, cargas que saltan de columna en columna a través del sólido orgánico en los mismos procesos que hacen de estos materiales candidatos a la electrónica molecular del futuro.

Te encuentras dentro de un túnel tallado no en piedra sino en ARN vivo, donde las paredes están formadas por hélices dobles de ARN ribosómico que se curvan sobre ti como columnas retorcidas de una catedral sumergida, sus surcos profundos brillando en azul cobalto y verde azulado bajo una luminiscencia que no viene de ninguna fuente externa sino del potencial químico mismo. Iones de magnesio salpican la superficie a intervalos regulares como remaches de luz amarilla intensa, cada uno anclado en el surco de una vuelta de ARN, neutralizando la carga negativa del esqueleto fosfato y dando a las paredes una regularidad arquitectónica casi industrial. Por el centro del canal, el pasaje se estrecha hasta apenas diez ångströms — una constricción tan absoluta que obliga a la cadena polipeptídica naciente a atravesarla residuo a residuo, serpenteando en ámbar cálido como un filamento de hueso fundido que apenas empieza a plegar sus primeras espirales alfa en la oscuridad molecular. En la periferia más remota del túnel, destellos de color cerceta parpadean y se apagan en fracciones de femtosegundo — eventos de hidrólisis de GTP que se expresan como pulsos de luminiscencia acuosa antes de disolverse en la presión implícita del solvente que ocupa cada ángstrom no reclamado por proteína o ARN.
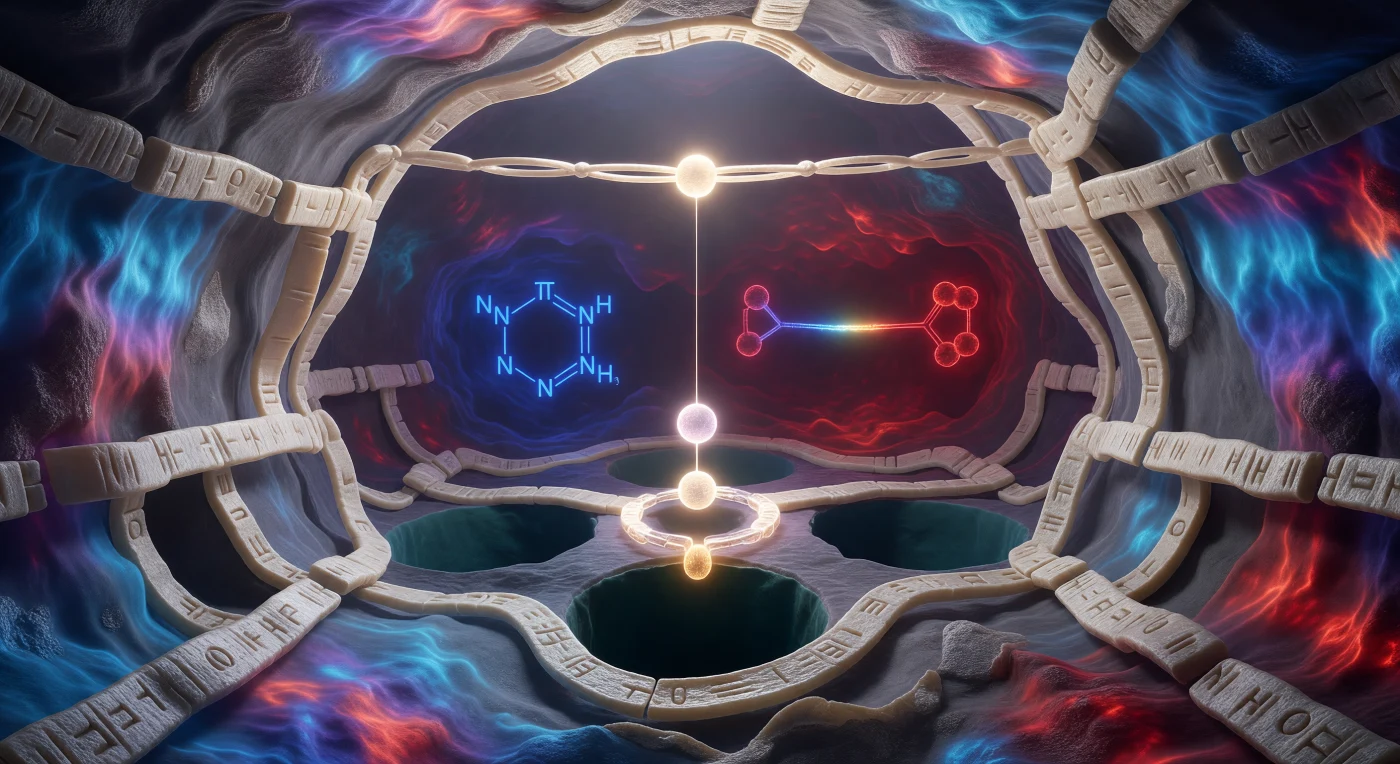
Desde el umbral de esta caverna de quince ångströms, la mirada se pierde en un interior que late con su propia luminosidad: las paredes de lámina beta se despliegan como mármol vivo surcado de relieve atómico, mientras un mapa de potencial electrostático las baña en una marea cromática que va del azul cerúleo hasta el carmesí profundo, revelando cada concentración de carga como si la roca misma fuera la fuente de luz. En el centro del escenario, la tríada catalítica se articula con la precisión de un mecanismo de relojería cuántica: el oxígeno hidroxilo de la Serina 195 arde en blanco dorado con sus pares de electrones libres orientados hacia el enlace peptídico del sustrato que cruza el techo de la caverna como un puente tenso a apenas tres ångströms de distancia, mientras el anillo imidazol de la Histidina flotante en azul cobalto aguarda su papel como lanzadera de protón, y el carboxilato del Aspartato ancla la pared posterior en borgoña pulsante, los tres nodos conectados por un gradiente de color que es, en sí mismo, un circuito molecular dibujado en luz. El aire —si puede llamarse así a este espacio de donde el agua ha sido desplazada— vibra con una tensión electrostática palpable: no hay gravedad aquí, sino fuerzas de van der Waals, puentes de hidrógeno y una agitación térmica que sacude cada residuo con amplitudes comparables a la longitud del propio enlace que está a punto de romperse. Uno percibe vísceramente que la catálisis no es un acontecimiento sino un estado de inminencia perpetua, una geometría tan perfectamente tensa que la frontera entre reactivo y producto parece una cuestión de femtosegundos.

Ante ti se extiende el borde de una lámina de grafeno suspendida en el vacío, una frontera de grosor imposible que divide el espacio en dos realidades absolutas: del canto, la hoja aparece como una línea oscura y quirúrgica de un solo átomo de espesor, con nubes electrónicas desbordándose en halos ámbar-dorados donde los enlaces colgantes se pierden en la nada; desde arriba, un infinito camisote hexagonal de plata y azul metálico se despliega hasta todos los horizontes, cada enlace carbono-carbono de 1,42 Å pulsando como un nodo brillante en la red. Esta geometría perfecta es el resultado de la hibridación sp² del carbono: seis átomos por anillo comparten una nube π deslocalizada que flota sobre y bajo el plano atómico como una película luminosa de platino pulido, responsable tanto de la extraordinaria conductividad eléctrica del grafeno como de su iridiscencia azul-metálica. Ondas flexurales lentas —fonones de longitud de onda entre 10 y 100 nm— recorren la superficie en suaves montículos topográficos que distorsionan levemente la simetría hexagonal, recordando que incluso esta superficie matemáticamente plana tiembla con energía térmica. En la distancia media, un defecto Stone-Wales —un par pentágono-heptágono nacido de la rotación de un enlace covalente— arde en naranja cálido contra el frío azul circundante, un punto de carbón enterrado en escarcha, concentrando estados electrónicos localizados antes de que la simetría perfecta se reafirme y la red cristalina retome su extensión silenciosa hacia cada horizonte.

Suspendido a la distancia exacta de la primera capa de hidratación, el ión Na⁺ ocupa el centro del campo visual como un sol comprimido en miniatura, irradiando una luminiscencia blanco-dorada de carácter puramente electrostático que no se dispersa sino que presiona el medio circundante con una precisión casi quirúrgica. Las seis moléculas de agua de la primera esfera de coordinación se mantienen en geometría octaédrica perfecta a 2,36 Å, con sus átomos de oxígeno orientados hacia el ión y bañados en un azul cerúleo-violeta intenso que delata el aumento local del orden dipolar y el desplazamiento hacia el azul de sus propiedades ópticas, mientras los extremos de hidrógeno, pálidos y cremosos, apuntan hacia el exterior como dedos de carga parcialmente positiva. Más allá, una segunda corona de doce a dieciocho moléculas parcialmente alineadas a ~4,5 Å forma un halo nebuloso donde la ordenación electrostática ya compite con la agitación térmica, produciendo un azul más difuso y fluctuante. Pasados los 7 Å, toda estructura reconocible se disuelve en la red de puentes de hidrógeno del agua líquida en equilibrio: un océano cerúleo profundo e inquieto, denso de reordenamientos moleculares que suceden en picosegundos y que no conservan ninguna memoria del campo iónico que domina tan cerca.

Te encuentras atrapado en el corazón ámbar de una proteína globular, rodeado en cada dirección por cadenas laterales no polares encajadas con una precisión que rivaliza con la de una gema tallada: leucinas y isoleucinas de tonos marfil y beige cálido se tocan a distancias de 3,5 a 4,0 Å en contactos C–C que no son vacío sino proximidad van der Waals pura, mientras discos aromáticos de fenilalanina —dorado pálido, casi platino— flotan en planos paralelos desplazados, emitiendo la fosforescencia fría de sus electrones π deslocalizados. De pronto, hacia la izquierda, el anillo indol de un triptófano irrumpe en azul verdoso intenso, su nube electrónica π irradiando un halo cerúleo que contrasta con el calor ambarino circundante, y un destello amarillo-dorado metálico delata el azufre enterrado de una metionina encajada entre texturas orgánicas más suaves. El empaquetamiento es tan denso —rozando 0,74 de fracción de ocupación— que no existe horizonte ni cielo abierto, solo superficies moleculares curvas cerrándose desde todos los ángulos, como el interior de una catedral excavada en cristal de caramelo translúcido. En el límite más lejano de la percepción, a quince o veinte ångströms de distancia que aquí equivalen a la boca de una caverna, el ámbar compacto comienza a abrirse en huecos ligeramente más anchos y una bruma azul-grisácea fría se filtra hacia adentro: la superficie hidrofílica de la proteína, donde las cadenas polares dan paso al mundo acuoso exterior, tan radicalmente distinto de esta intimidad molecular sellada como el aire lo es de un fósil de ámbar.

La superficie que se extiende bajo tus pies no es una simple membrana, sino un mundo proteico de geometría casi perfecta: un continente curvo de cápsides virales icosaédricas de 28 nanómetros de diámetro, cuyo horizonte molecular se curva con la suavidad majestuosa de un planetoide cristalino. El suelo está pavimentado con una arquitectura de rosetas pentaméricas y coronas hexaméricas, cada pétalo una subunidad proteica con sus propios valles en silla de montar y cúpulas suavemente ondulantes, mientras las depresiones de unión a receptores se hunden como lechos de lagos en añil y azul cobalto profundo, y los bucles inmunodominantes resplandecen en amarillo-blanco en sus cimas convexas. En las costuras entre subunidades adyacentes, los puentes salinos entre residuos cargados emiten destellos parpadeantes de carmesí y azul cobalto —nubes de oxígeno de aspartato en rojo vivo, halos de nitrógeno de lisina en frío añil—, chispas electrostáticas congeladas en un único fotograma de femtosegundo que mantienen unida la simetría icosaédrica con una tensión palpable aunque invisible. Por encima de la superficie, una capa de hidratación ordenada de dos a tres nanómetros de espesor cubre cada protuberancia y cada depresión con un velo translúcido de moléculas de agua dispuestas en redes de puentes de hidrógeno cuasi-cristalinas, refractando el resplandor electrónico ambiente en destellos azul-blancos a lo largo de cada cresta. El mundo se siente inmenso e íntimo a la vez: cada paso atravesaría un dominio proteico entero, y las microfluctuaciones térmicas de la cápside —su respiración silenciosa y constante— confieren al paisaje el carácter paciente y colosal de una energía geológica aprisionada en simetría perfecta.

Al asomarse a la boca de la triple hélice de colágeno y mirar a lo largo de su eje central, el ojo encuentra una arquitectura trenzada de escala ínfima pero de presencia monumental: tres cadenas polipeptídicas —ámbar, azul oceánico y verde jade— se enrollan entre sí con una simetría de tres ejes que se repite cada 8,7 ångströms, generando un ritmo geométrico tan preciso como el de un cristal y tan vivo como el de un tejido vivo. En el núcleo apretado de esta estructura, los residuos de glicina de las tres cadenas se aproximan hasta casi tocarse, separados apenas 3,9 Å entre sus carbonos alfa, un espacio tan estrecho que ningún aminoácido con cadena lateral podría caber sin desgarrar la hélice entera. La superficie interior del corredor helical está recorrida por los anillos pirrolidínicos de la prolina e hidroxiprolina, que sobresalen en sucesión rítmica como las costillas de una columna antigua, mientras una red secundaria de puentes de hidrógeno intercatenarios —dorados y translúcidos— y puentes agua-hidroxiprolina de tono aguamarina teje una malla luminosa entre las cadenas, visible como un vitral iluminado desde dentro. A esta escala, el calor molecular no es un concepto abstracto sino una vibración perpetua que difumina los bordes de los átomos más lejanos en una niebla cálida y fosforescente, recordando que cada enlace oscila con una frecuencia de femtosegundos y que la quietud, aquí, es simplemente imposible.

La mirada cae en picado hacia el interior del eje fibrilar como si se contemplara el fondo de un pozo sin límite, y lo que se abre ante los ojos no es vacío sino arquitectura: capas apiladas de láminas beta que retroceden con una regularidad tan absoluta —separadas exactamente 4,7 ángstroms entre sí— que el cerebro las percibe como obra de ingeniería antes que como producto de la biología. Estas estructuras son fibrillas amiloides, ensamblajes proteicos en los que cadenas peptídicas se pliegan en conformación beta extendida y se apilan perpendicularmente al eje de crecimiento, unidas lateralmente por escalones dorados de puentes de hidrógeno que conectan cada hebra con la siguiente en una red infinita de vínculos no covalentes. En el corazón del corredor, donde las dos láminas apuestas se entrelazan cara a cara, los residuos hidrofóbicos encajan con una precisión estérica tan perfecta que ninguna molécula de agua penetra jamás, generando esa seam oscura y densa —textura de basalto pulido— que concentra las fuerzas de dispersión de London en un contacto casi continuo. El exterior de la fibrilla, en cambio, irradia una corona azul-blanca de agua de solvatación ordenada, moléculas orientadas por los grupos polares del esqueleto peptídico que crean una costra nacarada y temblorosa en el límite donde lo líquido y lo semicristalino coexisten en equilibrio de picosegundos. Lo más perturbador es la repetición misma: cada capa es clon exacto de la anterior, un molde que se propaga solo, estrato a estrato, hacia atrás y hacia adelante en el tiempo con una fidelidad geométrica que convierte la enfermedad —porque estas fibrillas protagonizan el Alzheimer y otras amiloidosis— en una catedral de precisión molecular.
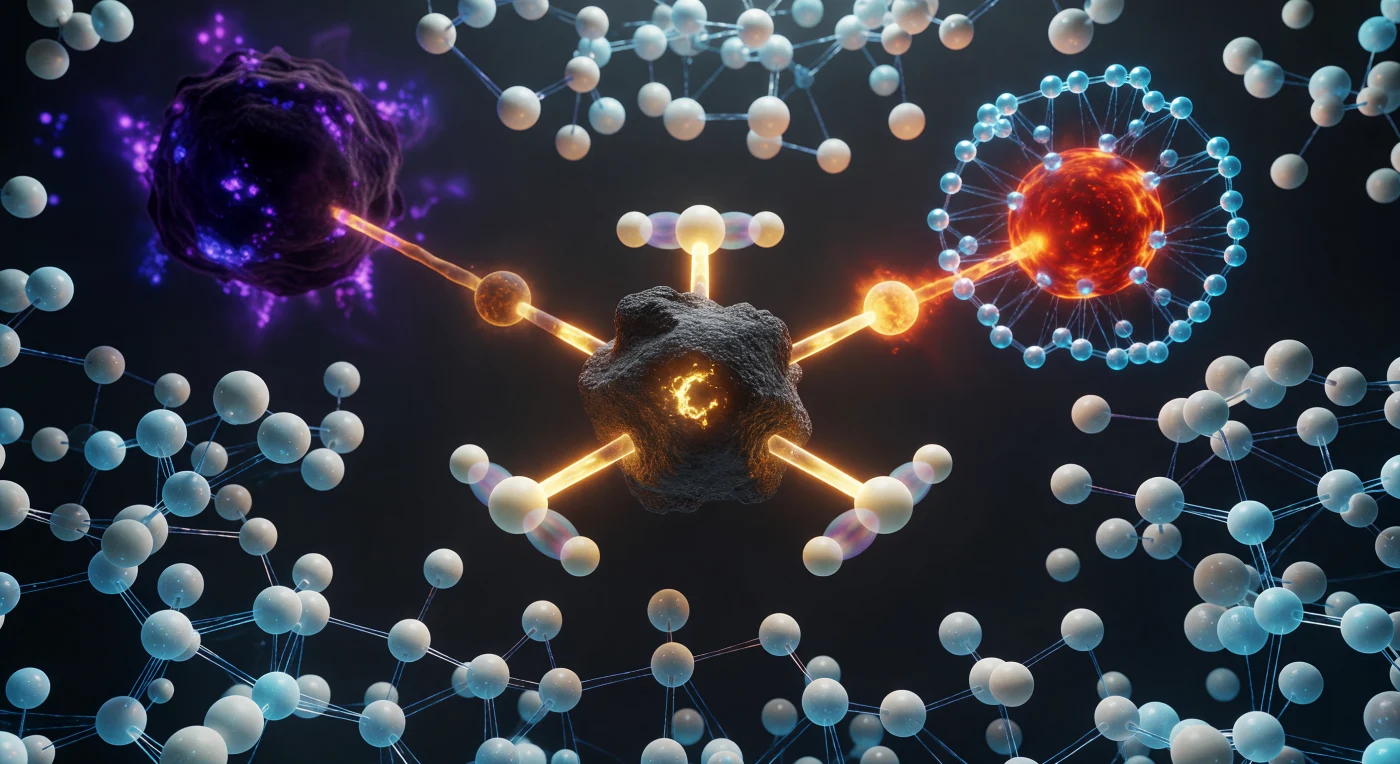
En este instante cuántico congelado, el átomo central de carbono emerge ante nosotros como un monolito oscuro atrapado en una geometría pentacoordinada imposible: tres hidrógenos bloqueados en un plano ecuatorial perfecto a 120° y dos enlaces parciales que se extienden en una línea recta, uno hacia el nucleófilo de oxígeno rojo-naranja que avanza a 2,0 Å y otro hacia el enorme bromo violeta que se aleja a 2,3 Å, ambos enlazados a medias y brillando con una luminiscencia ámbar-dorada que delata su estado de semiformación. Este es el estado de transición de una reacción de sustitución nucleofílica bimolecular, el punto más alto de la barrera energética donde la molécula existe ni como reactivo ni como producto, sino suspendida en el máximo del camino de reacción durante apenas decenas de femtosegundos antes de colapsar irreversiblemente hacia uno u otro lado. Las sombras translúcidas que acompañan a cada hidrógeno no son artefactos visuales sino la manifestación de la superposición cuántica y el efecto túnel, fenómenos que a esta escala de ángstroms son tan reales y determinantes como la gravedad lo es para los planetas. Todo el espacio circundante está colmado por moléculas de agua en reorganización dinámica, sus halos cian apretándose en tiempo real alrededor de las cargas parciales emergentes, porque en este mundo denso y sin gravedad son las fuerzas electrostáticas y los puentes de hidrógeno quienes dictan cada movimiento, y el universo térmico que aguarda fuera de este instante congelado empuja sin cesar desde todos los ángulos.

Te encuentras suspendido en el interior de un pasadizo mineral tan estrecho que las paredes parecen rozarte, un túnel elíptico tallado átomo a átomo en la estructura cristalina de una zeolita ZSM-5, donde nódulos grises de silicio y puentes de oxígeno rojo carmesí se repiten con la regularidad implacable de un andamiaje arquitectónico a escala cuántica. Ante ti, una sucesión de portales decagonales retrocede en perfecta perspectiva, cada anillo de diez miembros formado por tetraedros SiO₄ entrelazados que emiten un resplandor frío azul-blanco, mientras que puntos brillantes marcan los sitios de Brønsted donde átomos de aluminio sustituyen al silicio y concentran protones reactivos capaces de catalizar la ruptura y formación de enlaces carbono-carbono. A unos treinta angstroms, el canal sinusoidal perpendicular abre su boca en la pared lateral, creando una encrucijada molecular donde dos corredores de apenas 5,3 por 5,6 angstroms se cruzan en ángulo recto, un cuello de botella que obliga a cada molécula viajera a negociar su paso con la precisión geométrica de una llave en su cerradura. Las moléculas de hidrocarburo que colman el canal brillan en tonos dorados y ámbar, presionando contra las paredes con un contacto casi perfecto de van der Waals, encajadas en su huésped cristalino con una intimidad que disuelve toda frontera entre sólido mineral y entidad química.
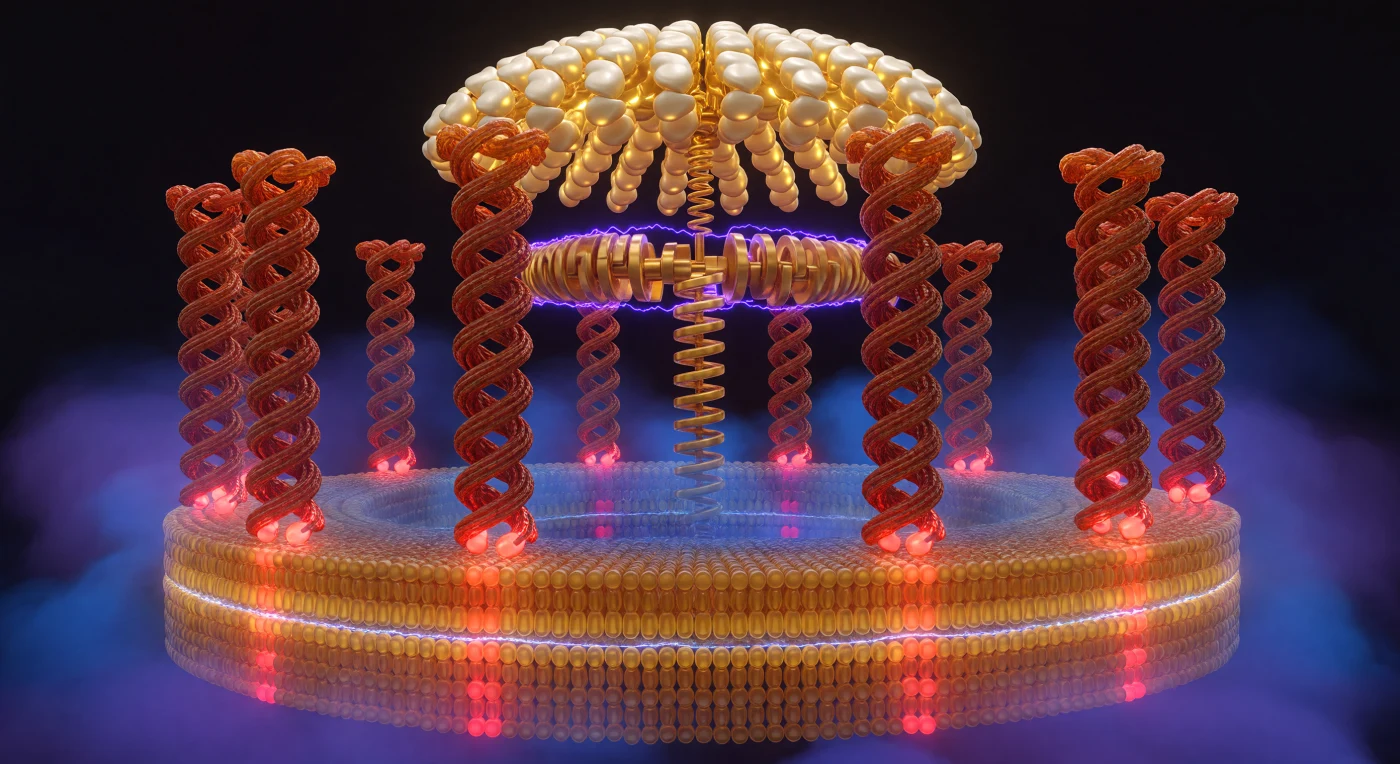
El observador se encuentra en el interior de un cilindro proteico de escala nanométrica, rodeado por diez columnas helicoidales de color terracota que forman el anillo c de la ATP sintasa, cada una separada de la siguiente por apenas unos pocos nanómetros de vacío cargado, una proximidad tan extrema que las superficies corrugadas de las hélices α parecen accesibles al mismo tiempo en ambos lados, como las paredes de un pasaje tallado en proteína viva. En la base de cada columna, los residuos de glutamato oscilan entre el rojo encendido de su estado desprotonado —expuesto al canal de protones y electrostáticamente desnudo— y el ladrillo apagado de su forma neutra enterrada en el núcleo hidrofóbico de la membrana, una alternancia que codifica el transporte de protones a través de la interfaz rotor-estátor y convierte el gradiente electroquímico en trabajo mecánico de rotación. La bicapa lipídica abraza la mitad inferior de este mundo cilíndrico en dos estratos diferenciados —ámbar dorado en el exterior, azul-ámbar más frío en el interior— y a través de ella se extiende un gradiente de voltaje de 150 a 200 milivoltios que no se anuncia con símbolos sino con la calidad de la luz misma: el violeta cobalto comprimido del lado citoplasmático se relaja en un índigo más cálido hacia la matriz mitocondrial, como si la atmósfera estuviera cargada hasta el umbral de la ruptura dieléctrica. Por encima de la membrana, la cúpula catalítica F₁ se arquea como una bóveda de proteína pálida en oro y marfil, sus seis subunidades β pulsando entre estados de ligación —oro fosforescente donde el ATP queda atrapado, blanco lunar donde el ADP espera— mientras el eje gamma desciende hasta el rotor con la inevitabilidad maquinada de un cigüeñal, transmitiendo el torque rotacional acumulado durante cuatro mil millones de años de evolución hacia la síntesis de la molécula energética fundamental de la vida.
