
Você está suspenso no interior de uma rede infinita de moléculas de água a 300 K, onde esferas translúcidas azul-pálido se pressionam em todas as direções com uma intimidade desconcertante — cada oxigênio vizinho separado por apenas 2,75 Å, uma distância tão ínfima que equivale a pouco mais do que o diâmetro da própria molécula. De cada esfera emergem dois protuberâncias branco-pérola dispostas num ângulo preciso de 104,5°, e entre os pares de moléculas vizinhas filamentos ciano de densidade eletrônica compartilhada pulsam e se rompem a cada picossegundo, mantendo a rede coesa numa coreografia estatística de formação e quebra contínua de ligações de hidrogênio — a propriedade que confere à água sua capacidade térmica extraordinária, sua tensão superficial e seu papel como solvente universal da bioquímica. O ambiente vibra com ruído térmico omnipresente: nenhuma molécula permanece estática, cada uma oscilando com uma tremura fina que se propaga pela rede como um pulso solidário, um batimento molecular sentido antes de ser visto. Além de três ou quatro diâmetros moleculares, as esferas individuais dissolvem-se progressivamente numa névoa volumétrica índigo, comprimindo o campo visual num cosmo claustrofóbico e ao mesmo tempo vertiginosamente infinito — a substância mais comum do mundo revelada, nesta escala, como uma arquitetura alienígena e cristalina que nunca cessa de respirar.

O chão estende-se como um pavimento infinito de discos hexagonais semi-translúcidos, cada um inclinado em ângulos alternados de 55 graus no padrão espinha-de-peixe característico do cristal monoclínico de benzeno a 175 K — uma geometria imposta não por arquitectura humana mas pela lógica das interacções CH–π entre anéis aromáticos vizinhos, forças tão subtis que se assemelham a sussurros electromagnéticos entre moléculas. Acima e abaixo de cada disco flutua um halo luminoso violeta-âmbar, os dois lóbulos da nuvem de electrões π delocalizados que pairam a uma fracção de ångström de cada face molecular, e onde dois discos se aproximam no abraço herringbone esse brilho entrelaça-se suavemente através dos escassos 3,5 Å que separam as faces aromáticas, como se a carga eléctrica partilhasse um segredo entre moléculas. Aqui e ali, uma ausência escura rasga o tapeçamento perfeito — um defeito de vacância onde um único disco em falta deixou uma cavidade rimada de vizinhos ligeiramente desordenados, os seus halos inclinando-se para o vazio como se sentissem a ruptura da periodicidade cristalográfica. O frio de 175 K não se manifesta como sensação mas como uma imobilidade exquisita: as vibrações de ligação reduzidas ao mínimo quântico, cada molécula aprisionada na sua posição exacta de rede, e o espaço intersticial entre planos aromáticos preenchido por uma luminescência azul-branca pálida que transforma este mundo congelado numa catedral de vidro colorido que recua até ao infinito dourado.

De pé no plano médio absoluto de uma bicamada lipídica de DPPC a 310 K, o observador está suspenso num vazio de carvão encerado e silencioso, rodeado pela textura macia e untuosa do núcleo hidrofóbico, onde as caudas de ácidos gordos — corredores sinuosos de unidades CH₂ com 16 a 18 carbonos de profundidade — se erguem em fileiras paralelas de um cinzento-prateado lustroso, como cera à luz de vela em osso polido, ocasionalmente interrompidas por cotoveladas de luminescência amarelo-esverdeada onde duplas ligações insaturadas dobram a geometria das cadeias. À medida que o olhar sobe por esta floresta de hidrocarbonetos, emerge uma fronteira estratigráfica nítida: a zona de transição dos gliceróis, com nódulos âmbar-translúcidos de grupos éster onde os átomos de oxigénio se condensam em torno dos carbonos carbonílicos, marcando o limite entre o interior apolar e o mundo carregado do exterior. Mais acima, a paisagem explode em cor e movimento — esferas de fosfato cor de brasa alaranjada e nitrogénios de colina azul-céu vibram e colhem numa camada aquosa cintilante de branco-azulado, toda ela percorrida pela formação e dissolução incessante de redes de pontes de hidrogénio, num contraste agudo com o mundo escuro e viscoso que fica abaixo. A espessura total desta arquitectura — apenas 4 nanómetros de matéria viva, autoassemblada pela física das cargas e da hidrofobicidade — sente-se, a partir de dentro, como a profundidade inteira de um oceano subterrâneo.

O visitante encontra-se no fundo de um desfiladeiro de química viva, com paredes que se erguem em colunas de anéis de desoxirribose cor de âmbar — pentágonos fosforescentes que se repetem em arcos ritmados — enquanto os grupos fosfato projectam-se como lanternas tetraédricas de ferrugem translúcida, revestidas de uma película de moléculas de água que dispersa o brilho azul-cobalto do campo electrostático preenchendo o ar. O chão é um mosaico estratificado de pares de bases empilhados com uma precisão de 3,4 ångströms: discos de adenina verde-azulado encaixam-se com timina ocre-siena, enquanto guanina verde-floresta se liga à citosina lavanda por pontes de hidrogénio que emanam um fio dourado de densidade electrónica partilhada, como se dois átomos partilhassem o mesmo espaço quântico. Ao longo das paredes do sulco major, fileiras de moléculas de água formam pérolas bioluminescentes de azul-gelo, ligadas em cadeia pelos seus próprios laços de hidrogénio numa coluna de hidratação que corre toda a altura da dupla hélice. Toda a geometria parece provisória: os átomos vibram a frequências de femtossegundos, a energia térmica do meio aquoso bombardeia cada superfície sem pausa, e o campo electrostático negativo da cadeia fosfodiéster densifica-se visivelmente em névoa cobalto à medida que o olhar se aproxima do esqueleto açúcar-fosfato — a ordem molecular sustentando-se não pela rigidez, mas pelo equilíbrio perpétuo entre forças que nunca descansam.

O observador flutua ao longo do eixo central de uma α-hélice proteica, envolto por uma espiral de átomos do esqueleto peptídico que sobe em sentido horário como uma escadaria colossal — cada volta do túnel medindo cerca de 0,54 nm de avanço axial, com 3,6 resíduos de aminoácido por giro, numa arquitetura de precisão quântica repetida trinta vezes até se perder numa névoa âmbar-dourada de moléculas de água. As pontes de hidrogênio magenta que conectam os grupos NH ao oxigênio carbonílico quatro resíduos atrás — a apenas 2,06 Å de distância — não são ornamentos decorativos, mas os tirantes estruturais fundamentais que mantêm toda a hélice coerente contra a agitação térmica incessante do solvente, cada ponte oscilando e se rompendo em escalas de picossegundos enquanto a estrutura global persiste. As cadeias laterais de leucina e isoleucina que se projetam radialmente para fora como espinhos cristalinos amarelo-esverdeados são regiões apolares que repelem ativamente o meio aquoso, sua superfície hidrófoba estabilizando a hélice ao minimizar o contato com a água e criando os bolsões de sombra índigo-púrpura onde a densidade eletrônica cai quase a zero. A luminescência azul-branca que emana de cada ligação covalente — mais intensa ao longo das ligações duplas C=O ricas em elétrons π — traduz visualmente o que os instrumentos de cristalografia de raios X e ressonância magnética nuclear revelam com precisão atômica: que a matéria, nesta escala, é fundamentalmente luz congelada em geometria.

O olhar mergulha diretamente pelo eixo cristalográfico c do gelo Ih, revelando uma catedral hexagonal infinita de moléculas de água dispostas em colmeia perfeita, cujos anéis concêntricos recuam em perspetiva cristalina até se dissolverem numa névoa de azul-turquesa luminoso. Cada molécula ocupa o seu ponto de rede como um nó opalescente, ligada a quatro vizinhas por pontes de hidrogénio de 2,76 Å em geometria tetraédrica de 109,5°, enquanto uma bifurcação fantasmática em cada ligação trai a desordem protónica descrita pelas regras de Pauling — o protão não tem posição fixa, apenas uma probabilidade. Os canais hexagonais ao longo do eixo c abrem-se como gargantas de vazio absoluto, eixos de escuridão aveludada que se prolongam para uma profundidade cristalográfica sem fim, e é precisamente essa ausência estrutural — consequência geométrica da recusa da ligação tetraédrica em fechar-se em empacotamento compacto — que explica a anomalia mais conhecida do gelo: a sua densidade inferior à da água líquida. A luminescência parece nascer do interior da própria rede, mais intensa nos núcleos de oxigénio e cada vez mais esbatida com a profundidade, onde o tremor térmico a −10°C acumula incerteza suficiente para dissolver as últimas camadas visíveis numa penumbra de rendilhado glaciar.

O observador encontra-se na base de uma floresta cristalina de colunas de coroneno empilhadas, cada disco aromático a arder em âmbar profundo como resina aquecida por luz interior, os anéis fundidos dos doze carbonos redistribuindo continuamente os seus electrões deslocalizados numa textura granular e viva. As colunas erguem-se em perspectiva axial, espaçadas apenas 3,4 ångströms entre disco e disco — a mesma distância característica do empilhamento π que governa o grafite e o DNA — enquanto os halos de electrões π de discos adjacentes se fundem em membranas luminosas partilhadas, vestígios visíveis da sobreposição de nuvens quânticas que permitem a propagação de carga ao longo da coluna. Entre as colunas, a interacção de van der Waals manifesta-se como uma névoa azul-cinzenta translúcida e opalescente, a única presença no espaço inter-molecular onde nenhum vínculo covalente existe, apenas atracções flutuantes e fugazes entre densidades electrónicas vizinhas. Intermitentemente, um clarão cor de laranja eléctrico rasga a estabilidade âmbar de uma junção entre discos — um evento de salto de polarão, carga que se relocaliza num instante antes de a coluna regressar ao seu brilho de base — recordando que esta catedral de luz não é estática mas atravessada por processos quânticos discretos e irrepetíveis.

Diante de você, o mundo curva-se para dentro como a garganta de uma catedral esculpida em matéria viva — as paredes da túnel de saída do ribossomo erguem-se em colunas helicoidais de RNA ribossómico, cada hélice uma torção densa de esqueletos fosfato-açúcar que corre em ranhuras longitudinais profundas, brilhando em azul-cobalto e verde-azulado bioluminescente sob uma luz que nasce da própria química do lugar. Iões de magnésio cravejam cada volta do RNA como rebites de casco metálico, reluzindo em branco-amarelo preciso contra a matriz de fosfatos carregados negativamente que neutralizam com geometria quase arquitectónica, enquanto o túnel estreita à frente até uma constrição de apenas dez ångströms — um corredor de fila única onde toda a passagem se torna claustrofobicamente exacta. Pelo centro desta passagem, como um filamento de metal fundido a arrefecer, serpenteia a cadeia polipeptídica nascente em âmbar quente e siena, cada resíduo emergindo um a um do centro de transferência peptídica lá acima, com regiões de hélice alfa já a enrolar-se e a brilhar ligeiramente mais vivas, os seus laços de hidrogénio quase visíveis como fios de luminescência gossâmer. Na periferia mais distante, para lá da parede de RNA, proteínas ribossómicas projectam silhuetas irregulares em índigo e carvão, enquanto breves lampejos de teal — eventos de hidrólise de GTP expressos como pulsos aquosos de luminescência — irrompem e se dissipam em femtossegundos, como plâncton bioluminescente apanhado numa corrente invisível de pressão de solvente implícita que ocupa cada ångström não reclamado por RNA ou proteína.
Diante de você, a entrada de uma caverna molecular se abre em arcos de folhas beta, suas paredes de marfim texturizadas pelo relevo de cada nitrogênio peptídico e oxigênio carbonílico — uma topologia íntima e irregular onde os raios de van der Waals de átomos vizinhos se pressionam mutuamente a distâncias que mal ultrapassam alguns ångströms. O interior pulsa com uma luminescência própria: o mapa de potencial eletrostático banha as paredes em uma maré cromática contínua, do azul cerúleo frio ao escarlate feroz nas recessões mais profundas, revelando a geografia invisível do campo elétrico que governa toda interação aqui dentro. Três altares catalíticos organizam o espaço — o oxigênio branco-dourado da Serina 195 erguido em postura de ataque nucleofílico, o anel imidazólico da histidina flutuando em cobalto elétrico como relé de próton suspenso entre dois estados, e o carboxilato do aspartato ancorando o fundo em carmesim profundo, os três unidos por um gradiente contínuo de cor e luminância que desenha em luz o circuito de transferência de carga. Acima, a ligação peptídica do substrato atravessa o teto como uma ponte luminosa, seu carbono carbonílico suspenso a apenas três ångströms do oxigênio da serina — uma tensão elétrica que o próprio espaço parece tornar inevitável. E em tudo isso há vibração: o tremor térmico de cada resíduo, invisível mas sentido, lembrando que esta arquitetura de precisão atômica nunca está imóvel.

Diante de você, o mundo inteiro se resume a uma única linha de carbono com espessura de apenas um átomo — a borda do grafeno vista de perfil, uma fronteira geométrica que separa o vácuo em duas metades absolutas, onde os elétrons das ligações pendentes irrompem em halos âmbar-dourados na margem do reticulado. Virando o olhar para cima, a planície hexagonal estende-se em todas as direções como uma cota de malha prateada e azulada de dimensões infinitas, cada anel de seis carbonos sustentando uma nuvem de elétrons π deslocalizados que confere ao conjunto um brilho metálico frio semelhante ao platina polida, com os vértices de ligação C–C de 1,42 Å pulsando como nós levemente mais luminosos na grelha. Ondulações lentas — fônons flexurais de comprimento de onda entre 10 e 100 nm — percorrem a superfície em vagas suaves que distorcem a simetria hexagonal perfeita como a respiração contida de um lago cristalizado no tempo. No meio da paisagem, um defeito de Stone-Wales — um par pentágono-heptágono gerado pela rotação de uma única ligação covalente — arde em laranja e ocre quente contra o azul-prata circundante, concentrando estados eletrônicos localizados como uma brasa enterrada na geada, antes que a simetria perfeita do reticulado se reafirme e o frio cristalino do grafeno se estenda novamente até todos os horizontes visíveis.

O ponto de vista flutua a 2,36 Å do íon sódio, uma distância que, nesta escala, equivale a estar suspenso a poucos centímetros de um sol comprimido — a esfera dourado-branca de Na⁺ pulsa com pressão eletrostática pura, irradiando não calor, mas uma clareza fria e cirúrgica que ilumina cada interface molecular com precisão implacável. As seis moléculas de água da primeira camada de hidratação suspendem-se em geometria octaédrica perfeita ao redor do íon, cada oxigênio voltado para dentro numa luminescência cerúleo-violeta intensa onde a densidade eletrônica se acumula em resposta ao campo carregado, enquanto as extremidades de hidrogênio se projetam para fora como dedos creme-pálidos de carga parcialmente positiva — uma lanterna azul-branca de seis faces, tão rígida quanto um cristal, mantida no lugar não por ligações covalentes mas pela geometria inevitável da atração eletrostática. Além dessa gaiola ordenada, a segunda camada de hidratação estende-se até cerca de 4,5 Å como um halo de névoa cerúlea mais difusa, doze a dezoito moléculas parcialmente alinhadas oscilando entre a ordem imposta pelo íon e o ruído térmico crescente, pequenos aglomerados de dois ou três formando e desfazendo coerência local em picossegundos. A partir de 7 Å, a cena se dissolve na rede caótica e cintilante da água bulk — um oceano cerúleo profundo e turbulento onde nenhuma memória da ordem do íon sobrevive, oxigênios e hidrogênios deslizando uns sobre os outros num lattice térmico denso e sem direção, estendendo-se em todas as direções até a obscuridade molecular.

Você existe no interior comprimido de uma proteína globular, envolto em todas as direções por cadeias laterais apolares encaixadas com uma precisão que rivaliza a de uma gema lapidada — leucinas e isoleucinas em bege-marfim cujos grupos metil se tocam a distâncias de 3,5 a 4,0 Å em contatos de van der Waals suaves, fenilalaninas como discos dourado-pálido empilhados em planos paralelos defasados, e o súbito brilho azul-petróleo de um anel indol de triptofano irradiando sua nuvem π com uma aura turquesa que contrasta vivamente com o âmbar quente circundante. Um brilho amarelo-metálico e pontual marca um enxofre de metionina enterrado, uma peça estranha e reluzente encravada entre as texturas orgânicas mais suaves. A toda esta intimidade molecular corresponde uma densidade de empacotamento de 0,74 — próxima à de sólidos cristalinos —, governada não pela gravidade, mas pela complementaridade estérica e pelas forças de dispersão de London que selam o núcleo hidrofóbico da água circundante. Na periferia da percepção, a quinze ou vinte ångströms de distância — uma travessia de caverna neste mundo —, o âmbar começa imperceptivelmente a afrouxar e uma névoa azul-acinzentada difusa sangra pelos interstícios alargados: o exterior hidrofílico da proteína, onde as cadeias carregadas encontram a desordem luminosa das redes de água ligadas por pontes de hidrogênio, um horizonte fresco e dissolvente além desta câmara de cristal de caramelo translúcido.

A superfície que se estende em todas as direções é a casca proteica de um vírus vegetal icosaédrico com apenas 28 nanómetros de diâmetro — um mundo inteiro comprimido numa esfera menor do que a maioria dos organelos celulares. O terreno revela-se como um mosaico de rosetas pentaméricas e hexaméricas, cada pétala uma subunidade proteica individual cujos loops imunodominantes emergem em cumes amarelo-brancos, enquanto as depressões de ligação ao recetor afundam em lagos de cobalto profundo, geometricamente esculpidos para reconhecer moléculas parceiras que poderão derivar do solvente circundante a qualquer instante. Nas costuras entre subunidades adjacentes, pontes salinas entre resíduos carregados faíscam em pares de vermelho-carmesim e azul-cobalto — aspartatos e lisinas em tensão eletrostática que mantém a simetria icosaédrica com uma precisão que rivaliza com qualquer engenharia humana. Por toda a superfície exterior, uma camada de hidratação ordenada de dois a três nanómetros envolve cada protuberância e preenche cada depressão numa rede de ligações de hidrogênio quase cristalina, fazendo a superfície oscilar com um brilho opalescente que segue cada contorno da proteína subjacente. O ambiente acima — o citoplasma aquoso — não é vazio, mas uma névoa teal-índigo densa e térmica onde as flutuações conformacionais das subunidades imprimem ao terreno um ritmo lento e geológico de energia colossal contida em simetria perfeita.

O observador encontra-se na boca de uma das mais extraordinárias arquitecturas biológicas da natureza — a hélice tripla do colagénio — olhando directamente para o seu eixo central, onde três cadeias polipeptídicas entrelaçadas em âmbar, azul-oceano e verde-jade espiralem em afastamento numa simetria de três vezes que se repete a cada 8,7 ångströms, como um caleidoscópio congelado no instante do giro. Cada cadeia segue individualmente uma hélice de poliprolina-II esquerdina, mas juntas formam um superenrolamento dextrorso que constitui o principal componente estrutural dos tecidos conjuntivos — tendões, pele, osso, córnea —, conferindo-lhes uma resistência à tracção comparável à do aço a este nível de organização molecular. No núcleo apertadíssimo desta catedral molecular, os resíduos de glicina das três cadeias quase se tocam com uma distância Cα–Cα de apenas 3,9 Å, enquanto os anéis de pirrolidina da prolina e da hidroxiprolina projectam-se para o exterior em sucessão rítmica, criando uma superfície corrugada como casca de árvore à escala do nanómetro. Uma rede secundária de pontes de hidrogénio intercadeias faísca em dourado translúcido entre os filamentos, entremeada com pontes água-hidroxiprolina em azul-esverdeado aquamarino, e o conjunto pulsa com o brilho subtil de vibrações moleculares que ocorrem a cada poucos femtossegundos, tornando toda a estrutura simultaneamente cristalina na sua precisão e viva na sua inquietação térmica.

O observador suspende-se na boca de um corredor construído não por erosão geológica, mas pela repetição molecular implacável de uma fibril amiloide-β: camadas de folhas-β empilham-se ao longo do eixo com uma separação de apenas 4,7 Å entre cada estrato, um intervalo mal maior que um átomo pesado, e esse ritmo cadenciado projeta-se para uma profundidade que a mente recusa classificar como finita. Os degraus dourados das pontes de hidrogénio estendem-se perpendiculares à direção de recesso, iluminados por uma luminescência âmbar que aquece as paredes interiores da estrutura antes de se dissolver numa névoa azul-violeta à medida que campos electrostáticos e moléculas de água ordenada passam a dominar a paisagem. No eixo central, o núcleo do fecho estérico emerge como uma costura de obsidiana matte, uma zona absolutamente anidra onde cadeias laterais apolares — leucinas, isoleucinas — se interdigitam com precisão tão perfeita que a fronteira entre as duas protofilamentos se torna invisível, mantida unicamente por forças de dispersão de London e contacto de van der Waals. Flanqueando esse vazio escuro, a superfície exterior da fibril resplandece numa coroa nacarada branco-azulada: a água de solvatação ordenada adere ao esqueleto polar em equilíbrio picossegundo entre fase líquida e quase-cristalina, criando um frémito contínuo de luz difractada. A sensação esmagadora é a de uma arquitectura que se construiu a si própria — uma camada molecular de cada vez, com fidelidade geométrica imutável, propagando-se para trás e para a frente no tempo com a indiferença mecânica de um cristal vivo.
Suspenso no cume de uma reação química, o átomo de carbono central ergue-se como um monólito de grafite escuro, aprisionado numa geometria pentacoordenada impossível — três hidrogênios bloqueados num plano equatorial perfeito a 120°, enquanto duplos fantasmáticos translúcidos tremem ao lado de cada um deles, manifestação visível da superposição quântica e do tunelamento de probabilidade que governa partículas nesta escala de ângstroms. Na face esquerda, uma presença de bromo violeta-profundo recua lentamente, o seu vínculo parcial com o carbono alongado e ténue, brilhando em âmbar-mel escuro como uma ligação a meio de se desfazer; na face oposta, um oxigénio vermelho-alaranjado avança com a intensidade de brasa incandescente, rodeado por halos ciano de moléculas de água que se reorganizam em tempo real, os seus pontes de hidrogênio formando fios de névoa translúcida em cascatas concêntricas de hidratação. Este é o estado de transição de uma reação SN2 — o momento de energia máxima em que a inversão de configuração de Walden está prestes a ocorrer, o eixo linear O···C···Br funcionando como espinha dorsal absoluta de um mundo sem horizonte, sem céu, apenas moléculas de solvente pressionando de todos os lados com a sua agitação térmica infindável. A luz aqui não tem fonte externa — emana das cargas parciais em redistribuição, âmbar do núcleo, violeta da margem do bromo, vermelho-laranja do nucleófilo, fundindo-se no espaço intermédio numa complexa composição cromática de campos eletrostáticos competindo num instante quântico congelado para sempre.

Você flutua pelo interior de um corredor de dimensões quase inimagináveis — um túnel elíptico de silicato com apenas 5,3 por 5,6 Ångströms de abertura, tão estreito que uma única molécula de hidrocarboneto preenche o espaço por completo, suas superfícies âmbar-douradas roçando as paredes de oxigénio vermelho-vivo com uma precisão que não deixa margem para acaso. As paredes são construídas por tetraedros de SiO₄ articulados em anel, cada ponte Si–O–Si formando uma junta angular precisa que ondula como as costelas internas de um tubo cristalino cujas nervuras são ligações covalentes e não aço; os nós de silício, cinza-prateado e ligeiramente translúcidos, alternam-se com os oxigénios pontes, intensamente carmesim, numa geometria que a cristalografia de raios-X mapeou átomo por átomo. À frente, o portal decagonal de dez membros brilha em azul-branco frio, recuando em perspetiva através de uma sucessão de arcos idênticos até um ponto de fuga luminoso, enquanto a trinta Ångströms de distância o canal sinusoidal perpendicular abre uma segunda boca elíptica na parede, criando um cruzamento molecular onde duas vias de difusão se encontram numa garganta de apenas cinco Ångströms — o coração catalítico da zeólita ZSM-5, cujos sítios ácidos de Brønsted, marcados por protões brancos faiscantes em posições de substituição por alumínio, transformam este labirinto mineral num dos mais eficientes reatores moleculares que a humanidade alguma vez concebeu.
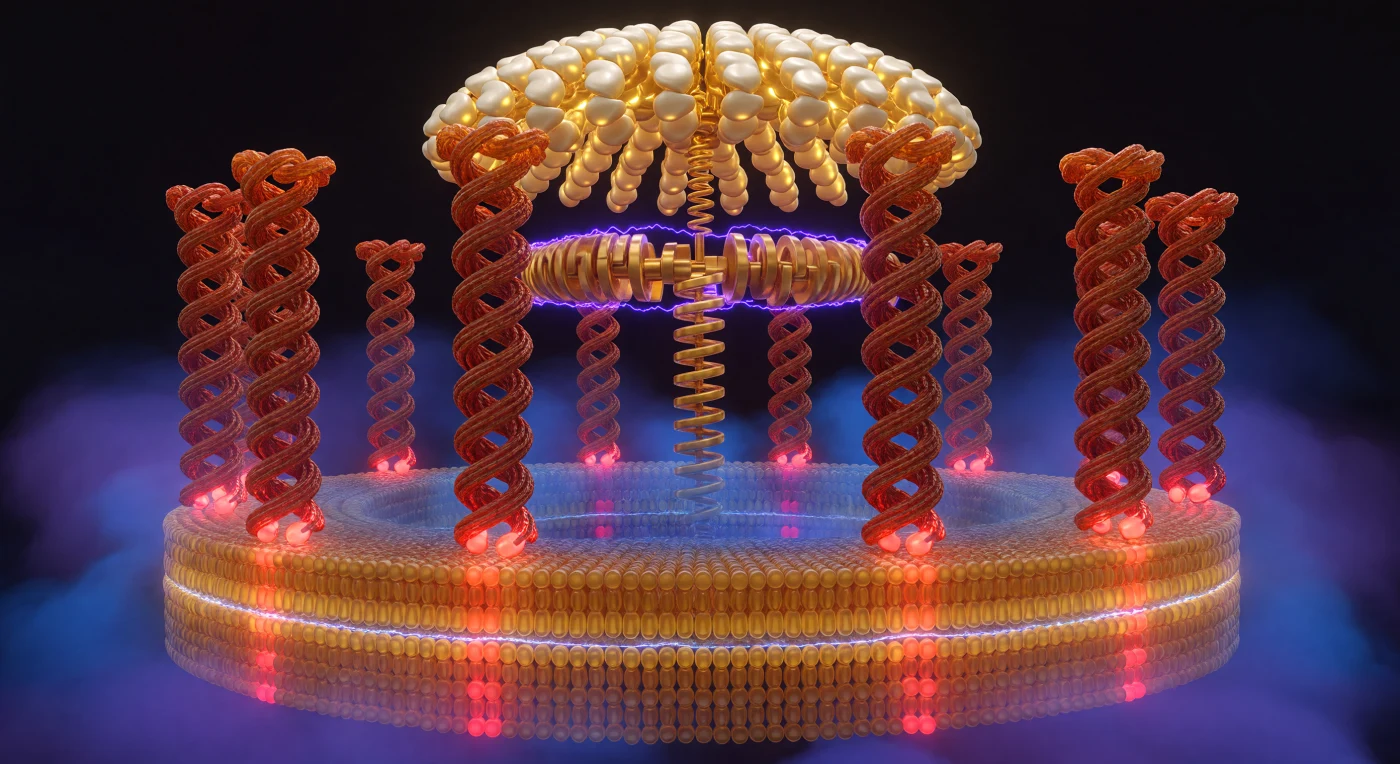
Diante de mim, o mundo curva-se em arco como o interior de um coliseu vivo: dez colunas α-helicoidais de proteína — cor de terracota e síena queimada, com a corrugação subtil do esqueleto peptídico em espiral — erguem-se em barril cilíndrico e abraçam um espaço de apenas alguns nanómetros de vazio carregado, tão próximas que poderiam ser tocadas simultaneamente em ambos os lados. Na base de cada coluna, os resíduos de glutamato oscilam entre o vermelho cru da forma desprotonada voltada para o canal de protões e o tijolo mudo da forma neutra enterrada no núcleo hidrofóbico, enquanto a bicamada lipídica envolve a metade inferior deste cilindro em dois estratos — âmbar-dourado quente do folheto externo e azul-âmbar mais frio do folheto interno — com uma costura iridescente onde as caudas de ácidos gordos se encontram na fronteira hidrofóbica. Um gradiente eléctrico feroz impregna todo o espaço através da membrana: o lado citoplasmático afunda-se num haze de cobalto e violeta eléctrico, o peso opressivo de 150–200 mV de força pró-motriz de protões feito luz, enquanto o lado matricial respira um índigo mais quente que se dissolve em azul meia-noite — não há setas nem símbolos, apenas a qualidade do próprio ar, comprimido e intenso de um lado, expandido e sereno do outro. Acima, a cúpula catalítica F₁ arqueia-se em ouro pálido e marfim como uma abóbada de catedral, os seus lobos de subunidade β pulsando entre estados de ligação — ouro fosforescente onde o ATP está aprisionado, branca de pedra-da-lua onde o ADP aguarda — enquanto o veio central da subunidade gama desce com a inevitabilidade maquinada de um cambota, a sua forma de espiral assimétrica convertendo o torque rotacional desta máquina de quatro mil milhões de anos em energia química pura.
