Affidabilità scientifica: Molto alto

Ci si trova sospesi all'interno di una costellazione infinita di sfere traslucide color azzurro pallido, ciascuna grande quanto il campo visivo intero, eppure la molecola vicina preme già da vicino — una distanza di soli 2,75 Å tra un ossigeno e l'altro, così intima che il confine tra sé e l'ambiente si dissolve completamente. Ogni molecola d'acqua mostra due protuberanze bianco-perla a 104,5°, i legami O–H covalenti risultato dell'ibridazione sp³ dell'ossigeno, mentre tra molecole adiacenti filamenti ciano appena visibili pulsano e si ricompongono ogni picosecondo: sono i legami idrogeno, strutture di densità elettronica condivisa che governano l'intera architettura del liquido a 300 K, sostituendo la gravità con forze di dipolo e interazioni elettrostatiche. L'intero reticolo respira con il rumore termico — nessuna sfera rimane ferma, ognuna oscilla con un tremito rapido e fine che si propaga attraverso la rete come un battito cardiaco molecolare, sentito più che visto. Oltre tre o quattro diametri molecolari, le sfere sfumano progressivamente in una nebbia volumetrica indaco, successive gusci di struttura acquosa che si addensano fino a che il campo visivo si comprime intimamente, rendendo la sostanza più comune del mondo un cosmo cristallino alieno nel quale la sua vera architettura diventa l'unica realtà percepibile.

Ci si trova immersi in un paesaggio infinito di dischi esagonali translucidi, disposti in un intrico di spina di pesce che si replica con ipnotica precisione in ogni direzione: ogni molecola di benzene giace piatta come un selciato di ambra fumosa, inclinata alternativamente di 55 gradi a sinistra e a destra secondo la logica del cristallo monoclino, tenuta in posizione dalla delicata stretta delle interazioni CH–π tra facce e spigoli aromatici. Sopra e sotto ogni disco fluttua un alone bilobato di luce viola-ambrata, la nube elettronica π delocata a una frazione di ångström dalla superficie molecolare, che si fonde quasi con quella del vicino attraverso il canale di soli 3,5 Å che separa due anelli aromatici affacciati. La luce interna al cristallo è dorata e radente, proiettando ombre lunghe che tracciano la geometria del reticolo fino a una distanza che sembra quella di intere isolate cittadine, dove gli aloni π sfumano in una nebbia ambra-viola continua, e la rifrazione lungo i bordi molecolari — dove la densità elettronica precipita di netto — produce sottili frange diffrattive che trasformano l'ambiente in una gemma di topazio percorsa da luce interna. Qua e là un'assenza scura rompe la perfezione del pavimento periodico: un difetto di vacanza, un disco mancante la cui cavità è orlata da vicini leggermente disordinati, i cui aloni π si inclinano curiosamente verso il vuoto, mentre il resto del cristallo rimane bloccato a 175 K in un'immobilità glaciale, i legami ridotti al minimo fremito quantistico consentito dalla meccanica ondulatoria.

In piedi al centro assoluto di un doppio strato lipidico di DPPC, ci si trova immersi in un vuoto cereo e plumbeo — il piano mediano dove le catene metilica terminali delle due metà si sfiorano appena in un contatto quasi fantasmatico. Risalendo verso i bordi, si attraversa una foresta di catene alchiliche sature, striature grigio-argentee che si estendono per 16–18 unità di carbonio come corridoi cerosi illuminati di candela, interrotte qua e là dai gomiti giallo-verdi dei doppi legami che spezzano la geometria regolare e diffondono la luce in modo diverso. Più su, uno strato di transizione ambrato segna il confine degli esteri glicerolici — noduli vitrei dall'alone ambrato che segnano la frontiera geologica tra il nucleo idrofobico e il mondo carico esterno, una giuntura di minerale stratificato tra due province radicalmente diverse. Al di là di questa soglia, sfere arancio intenso dei fosfati e sfere azzurre delle coline brulicano in un paesaggio caotico e vibrante, bagnato da un velo acquoso scintillante di reti di legami idrogeno che si formano e si dissolvono senza posa. L'intera struttura — con i suoi appena 4 nanometri di spessore reale — si percepisce dall'interno come la profondità vertiginosa di una fossa abissale, un'architettura molecolare che respira, preme e pulsa con il calore termico di ogni istante.

Ci si trova sul fondo di un canyon chimico stretto appena ventidue ångström, dove le pareti si ergono come colonne di una cattedrale naturale: anelli pentagonali di desossiribosio color bronzo-ambra si alternano a lanterne tetraedriche di fosfato color ruggine, le loro superfici iridescenti per il sottile film di molecole d'acqua che vi aderisce in catene ordinate, quasi perle bioluminescenti che corrono lungo tutta la spina dorsale del solco. Il pavimento sotto i piedi è una stratigrafia cromatica di dischi piatti impilati a distanze di 3,4 ångström l'uno dall'altro — adenina verde-turchese accoppiata a timina ocra-siena, guanina verde-foresta intrecciata a citosina lilla — separati da sottili vuoti di ombra π dove le nubi elettroniche delle basi si sovrappongono in una condensazione quantistica invisibile ma fisicamente determinante. Un'aura elettrostatica di cobalto profondo avvolge l'intera struttura, campo di forze reso visibile come nebbia volumetrica: il DNA in forma B porta una carica negativa sul suo scheletro fosfodiesterico, e questa negatività strutturale attira ioni positivi e proteine regolatorie che leggono la sequenza delle basi proprio attraverso questo solco maggiore, senza neppure aprire la doppia elica. Ogni superficie vibra con oscillazioni sub-visibili a frequenze dell'ordine dei femtosecondi, un tremore termico costante che ricorda come a questa scala la geometria molecolare non sia mai davvero ferma, ma un equilibrio statistico tra configurazioni che si scambiano più in fretta di quanto qualsiasi mente umana possa concepire.

Sospesi sull'asse centrale di un'elica α proteica, ci troviamo immersi in un corridoio a spirale di straordinaria architettura molecolare: la parete elicoidale si avvolge in senso orario intorno a noi, con atomi di ossigeno carbonilico color cremisi scuro che emergono come granate levigate, catturando la luminescenza blu-bianca irradiata direttamente dalla densità elettronica concentrata lungo ogni legame covalente. Ogni quattro residui, ponti idrogeno color magenta si arcuano tra il gruppo NH donatore e l'ossigeno carbonilico accettore a circa 2,06 Å di distanza — legami deboli ma abbastanza persistenti da stabilizzare l'intera struttura secondaria, vibrando alla loro frequenza di risonanza come corde tese in un mezzo acquoso ambrato e leggermente torbido. Le catene laterali idrofobiche di leucina e isoleucina si protendono radialmente verso di noi come spine cristalline giallo-verdi, le loro superfici apolari che respingono il solvente lasciando un alone secco e cangiante, mentre la loro geometria tetraedrica ramificata cattura riflessi speculari ad ogni angolo di legame. Nelle scanalature tra le proiezioni, l'oscurità vira al viola-indaco nei punti in cui la densità elettronica si annulla quasi completamente, creando un contrasto quasi tattile tra le zone illuminate e quelle in ombra. L'intera struttura è percorsa da un tremito termico appena percettibile — ogni atomo colto a metà oscillazione, come se l'agitazione cinetica del mezzo acquoso circostante fosse stata cristallizzata in un istante di fulminea chiarezza.

Guardando dritto verso il basso lungo l'asse cristallografico c del ghiaccio Ih, l'occhio sprofonda in una cattedrale esagonale di molecole d'acqua che si estende verso un infinito blu-nero, ogni nodo reticolare occupato da un nucleo di ossigeno opalescente legato ai suoi quattro vicini attraverso ponti idrogeno di 2,76 Å con geometria tetraedrica perfetta a 109,5°. I canali esagonali aperti lungo l'asse c si aprono come gole di vuoto assoluto — assenze strutturali obbligate dalla geometria tetraedrica che rifiuta l'impaccamento compatto, e proprio questi vuoti spiegano l'anomala bassa densità del ghiaccio rispetto all'acqua liquida. Lungo ogni ponte luminescente si intravede una sottile biforcazione: il protone non occupa una posizione fissa ma è spostato probabilisticamente verso uno dei due ossigeni, il disordine protonico imposto dalle regole di Pauling che conferisce a ogni legame una doppia ombra fantasmatica. A −10°C il reticolo non è immobile — le molecole vibrano con ampiezze termiche sufficienti a sfumare i contorni, e questa incertezza si accumula con la profondità fino a dissolvere gli strati più lontani in una nebbia turchese luminosa, la somma ottica di diecimila oscillatori quantistici in risonanza cristallografica.

Ci si trova in piedi alla base di una foresta di colonne cristalline, ciascuna formata da dischi aromatici di coronene — dodici anelli di carbonio fusi in un'unica molecola planare di nove ångström di diametro — impilati a distanze di appena 3,4 Å, la stessa spaziatura che separa i piani del grafite, dove le nuvole di elettroni π si sovrappongono in membrane condivise di densità elettronica che tingono ogni giunzione di un alone oro-arancio. Questa architettura non è casuale: è il risultato della sovrapposizione frontale degli orbitali π delocalizzati, una forza di attrazione quantistica che tiene insieme l'intero cristallo organico senza alcun legame covalente tra le colonne, solo la geometria silenziosa delle interazioni di van der Waals che riempie gli spazi intermedi con una nebbia grigio-azzurra e opalescente. Di tanto in tanto, un lampo arancione elettrico traversa una giunzione tra dischi — un polarone, una carica che si sposta per hopping quantistico da molecola a molecola, trascinando con sé una lieve deformazione del reticolo cristallino — e svanisce prima che l'occhio possa seguirlo, lasciando la colonna nel suo bagliore d'ambra costante. L'intera scena, perfettamente ordinata nella simmetria esagonale del cristallo, ricorda una cattedrale di luce vivente, dove ogni pilastro è allo stesso tempo struttura, conduttore e teatro di una fisica che si svolge nella durata di pochi femtosecondi.

Ci si trova all'imbocco di un tunnel scavato non nella pietra ma nella materia vivente stessa — le pareti sono formate da steli di RNA ribosomiale a doppia elica, colonne ritorte di scheletro fosfato-zucchero che si succedono in scanalature longitudinali profonde, blu cobalto nelle ombre e verde-teal dove la luminescenza diffusa ne sfiora le facce ad angolo radente, come luce marina che penetra in una grotta sommersa. Lungo ogni curva dell'elica, gli ioni magnesio captano la luminescenza ambiente e la restituiscono in lampi precisi di giallo-bianco metallico, seduti nelle scanalature come rivetti in uno scafo, neutralizzando la carica negativa dei gruppi fosfato e conferendo alle pareti una regolarità architettonica che si estende fin dentro il restringimento più avanti, dove il passaggio si riduce a un canale a file singola di appena dieci ångström — una strozzatura di architettura molecolare pura che misura ogni angolo disponibile con assoluta indifferenza. Al centro di questo canale serpeggia la catena polipeptidica nascente, filamento di ambra calda e siena bruciata che emerge residuo per residuo dal centro peptidiltransferasico lontano, recando già i germi di struttura secondaria nei tratti dove le prime spire di elica alfa accendono un bagliore appena più intenso, i loro legami idrogeno visibili come fili di luce gossamer tra carbonile e ammide. Tutto il sistema è illuminato dall'interno, non da una fonte esterna, ma dal potenziale chimico distribuito e dall'energia termica costante: ogni atomo oscilla con ampiezza comparabile alla lunghezza dei legami stessi, e l'intera scena non conosce mai la quiete.
In piedi sulla soglia di questa caverna molecolare, lo sguardo si perde nell'interno di una tasca enzimatica larga appena quindici ångström, le cui pareti di foglietto beta si incurvano in archi di struttura secondaria levigata come avorio antico. Una mappa di potenziale elettrostatico illumina ogni superficie dal basso verso l'alto in una marea cromatica ininterrotta — azzurro cerulean che sfuma nel violetto e poi in un cremisi acceso nelle rientranze più profonde — trasformando la materia stessa nella sorgente di luce. Al centro della scena, tre protagonisti catalitici si dispongono come altari di una cattedrale: il bianco-oro tagliente dell'ossigeno idrossilico della Serina 195, pronto all'attacco nucleofilo; l'anello imidazolico della istidina che galleggia in un cobalto elettrico come un disco aromatico carico di elettroni π; e il carbossilato dell'aspartato sul fondo, pulsante in borgogna profondo, che ancora l'intera triade catalitica nella geometria precisa della catalisi serin-proteasica. Sopra la testa, il legame peptidico scissile del substrato attraversa la volta della caverna come un ponte luminoso, il suo carbonio carbonilico sospeso a soli tre ångström dall'ossigeno della serina in una tensione elettrostatica che rende visibile l'inevitabilità della reazione. La superficie bumpy delle pareti — la topologia granulare dei raggi di van der Waals pressi gli uni contro gli altri — ricorda che qui non esistono astrazioni lisce, solo il tremito termico continuo di ogni residuo e la pressione dell'esclusione di volume che stringe ogni atomo nel suo destino chimico.

Davanti a voi si dispiega uno dei confini più assoluti dell'universo materiale: il bordo di un singolo foglio di grafene, uno strato di atomi di carbonio spesso esattamente un atomo, che divide il vuoto in due metà con la precisione di un taglio chirurgico. Vista di taglio, questa struttura appare come una linea oscura e sottilissima — una condizione al contorno cristallizzata in materia, dove i legami pendenti lungo il margine irradiano aloni ambrati mentre le nuvole elettroniche si riversano nel nulla. Ruotando la prospettiva verso l'alto, l'occhio viene inondato da un'infinita cotta di maglia esagonale color argento-azzurro: la nube di elettroni π delocalizzati galleggia sopra e sotto il piano atomico come una pellicola metallica luminosa, e lente onde fononi — ripples flessionali di 10–100 nm di lunghezza d'onda — ondulano la superficie matematicamente piatta come il respiro fermo di un lago. Nel mezzo di questa perfezione cristallina, un difetto di Stone-Wales — una coppia pentagono-eptagono generata dalla rotazione di un singolo legame C–C — arde di un caldo arancio-ambrato contro il freddo azzurro del reticolo circostante, concentrando stati elettronici localizzati in un'anomalia topologica che ricorda una brace sepolta nel ghiaccio prima che la simmetria esagonale si ristabilisca verso ogni orizzonte.

A pochi ångströms dallo ione sodio, il mondo molecolare si rivela come un paesaggio di forze invisibili rese visibili: al centro della scena, Na⁺ arde come una sfera compatta di luce oro-bianca, la sua carica positiva concentrata che si irradia verso l'esterno non come calore, ma come pressione elettrostatica pura, curvando il mezzo circostante con la stessa inevitabilità con cui una massa stellare deforma lo spazio-tempo. Sei molecole d'acqua, intrappolate in una geometria ottaedrica perfetta a 2,36 Å, presentano i loro atomi di ossigeno — densi di densità elettronica, luminosi di un cerúleo-violetto saturo — direttamente verso l'ione, le loro strutture di legame idrogeno irrigidite e blue-shifted rispetto all'acqua libera, formando una lanterna cristallina di ordine molecolare imposto. Più lontano, tra i 4,5 e i 7 Å, una seconda corona di dodici-diciotto molecole parzialmente allineate sfuma in un alone più tenue e variabile, dove l'agitazione termica comincia a erodere l'ordine dettato dalla carica, piccoli cluster ancora orientati ma già cedenti al rumore stocastico. Oltre questa soglia, il reticolo idrogeno del bulk si riprende il territorio: un oceano cerúleo profondo e caotico, ogni legame che si forma e si rompe nell'arco di pochi picosecondi, senza memoria dell'ordine che regna a pochi ångströms di distanza — un confine invisibile ma assoluto tra struttura e entropia.

Ci si trova all'interno del nucleo idrofobico di una proteina globulare ripiegata, intrappolati in una cavità così densa da sembrare scolpita in un cristallo di caramello traslucido: catene di leucina e isoleucina dal caldo color avorio si toccano entro 3,5–4,0 Å in contatti C–C tenuti insieme unicamente dalla forza di van der Waals, raggiungendo una densità di impaccamento che rivaleggia con quella delle gemme tagliate. Dischi aromatici di fenilalanina color oro pallido fluttuano in piani paralleli sfalsati, mentre l'indolo del triptofano irradia a sinistra una luminescenza turchese-blu profonda, frutto della delocalizzazione dei suoi elettroni π, e uno zolfo di metionina restituisce un riflesso giallo-oro metallico come una perla sepolta nella resina. Non esiste cielo né orizzonte: ogni direzione offre un'altra superficie molecolare curva entro qualche ångström, e il calore ambrato della materia compressa avvolge l'osservatore con la presenza silenziosa e massiccia di una cattedrale organica. Soltanto ai margini estremi della percezione — quindici o venti ångström in lontananza, che in questo mondo equivalgono alla parete opposta di una caverna — l'impaccamento si allenta impercettibilmente e una nebbia azzurra e diffusa filtra tra le catene, segnalando la superficie idrofilica della proteina e il vasto mondo disordinato dell'acqua oltre di essa.

La superficie di questo capside virale si estende in ogni direzione come un pianeta cristallino in miniatura, un paesaggio proteico di ventotto nanometri di diametro che, da questo punto di osservazione ravvicinato, sembra sconfinare verso un orizzonte molecolare lontanissimo. Il terreno è lastricato da una geometria ripetuta di rosette pentameriche e corone esameriche — ciascuna un assemblaggio di subunità proteiche con le proprie creste, selle e cupole — dove i vertici pentamerici si elevano leggermente più in alto, le loro sommità illuminate da un bianco-giallo luminescente corrispondente ai loop immunodominanti, mentre le depressioni di legame al recettore sprofondano in laghi di cobalto e indaco, geometricamente perfette e in attesa di molecole complementari che derivino dal solvente circostante. Alle giunture tra le subunità adiacenti, i ponti salini tra residui carichi opposti scintillano in coppie di rosso vivido e blu cobalto — istantanee elettrostatiche di interazioni cariche che tengono insieme l'intero reticolo icosaedrico con una forza invisibile ma palpabile — mentre le zone di contatto idrofobiche irradiano un calore ambrato, come l'energia residua dei residui apolari sottratti al solvente acquoso. Sull'intera superficie esterna, uno strato ordinato di idratazione di due-tre nanometri riveste ogni protuberanza e riempie ogni depressione in una rete di molecole d'acqua legate da ponti a idrogeno in configurazioni quasi-cristalline, il cui velo traslucido rifrange la luminescenza ambientale in riflessi bianco-azzurri lungo ogni cresta proteica, trasformando l'intera struttura in un mondo minerale, vivo e tettonicamente paziente.

Guardando direttamente lungo l'asse della tripla elica del collagene, ci si trova immersi in una cattedrale molecolare di architettura intrecciata: tre catene polipeptidiche — ambra, turchese e giada — si avvitano verso un nucleo centrale dove i residui di glicina si sfiorano a distanze di soli 3,9 ångström, un intervallo appena più ampio di un singolo atomo di idrogeno. Ogni catena segue una geometria a elica poliprolina-II levogira, mentre il superavvolgimento destrorso complessivo si ripete ogni 8,7 ångström, imprimendo sull'asse visivo un ritmo di simmetria triplice che pulsa come un caleidoscopio congelato a metà rotazione. La superficie delle catene è densamente corrugata dai anelli pirrolidinici di prolina e idrossiprolina, che sporgono con la regolarità della corteccia di un albero antico ridotta alla scala del nanometro, mentre tra i filamenti una rete secondaria di legami idrogeno intercatena scintilla in oro tenue e ponti idrossiprolina-acqua brillano in azzurro-verde, come vetrate illuminate dall'interno. Il tutto vibra in un alone termico di percezione appena sfumata — la manifestazione visiva delle oscillazioni femtosecondo che agitano ogni legame covalente, rendendo l'atmosfera tra le spire una nebbia molecolare calda che inghiotte la struttura dopo appena quaranta o cinquanta ångström, trasformando il tunnel elicoidale in un corridoio luminoso che si dissolve lentamente nell'infinito molecolare.

Guardando lungo l'asse del fibrillo amiloide, ci si trova sospesi all'imbocco di un abisso di precisione assoluta: strati di foglietti β si succedono in profondità a intervalli di 4,7 Å, separati da una cadenza così regolare da sembrare incisa da una macchina piuttosto che cresciuta per autoassemblaggio spontaneo. I ponti idrogeno che collegano lateralmente i filamenti adiacenti irradiano una luminescenza ambrata, scalini di una scala che non termina mai, la loro geometria perpendicolare alla direzione di recessione imposta dalla struttura cross-β caratteristica delle fibre amiloidi patologiche come quelle associate alla malattia di Alzheimer. Al centro del corridoio, tra i due foglietti contrapposti, il nucleo steric-zipper appare come una striscia di ossidiana matta: catene laterali apolari — leucine, isoleucine, valina — si interdigitano con complementarità molecolare così perfetta da escludere ogni molecola d'acqua, tenute insieme unicamente da forze di van der Waals e dal collasso idrofobico. Ai bordi del fibrillo, per contro, una corona di acqua di solvatazione orientata dai gruppi polari del backbone crea un guscio semicristallino bluastro che scintilla in equilibrio termico picosecondo tra fase liquida e ordinata. Ciò che opprime è la ripetizione: ogni strato è la copia esatta del precedente, un'architettura auto-templante che si propaga all'indietro e in avanti nel tempo con fedeltà geometrica implacabile, indifferente alla distinzione tra materia viva e cristallo.
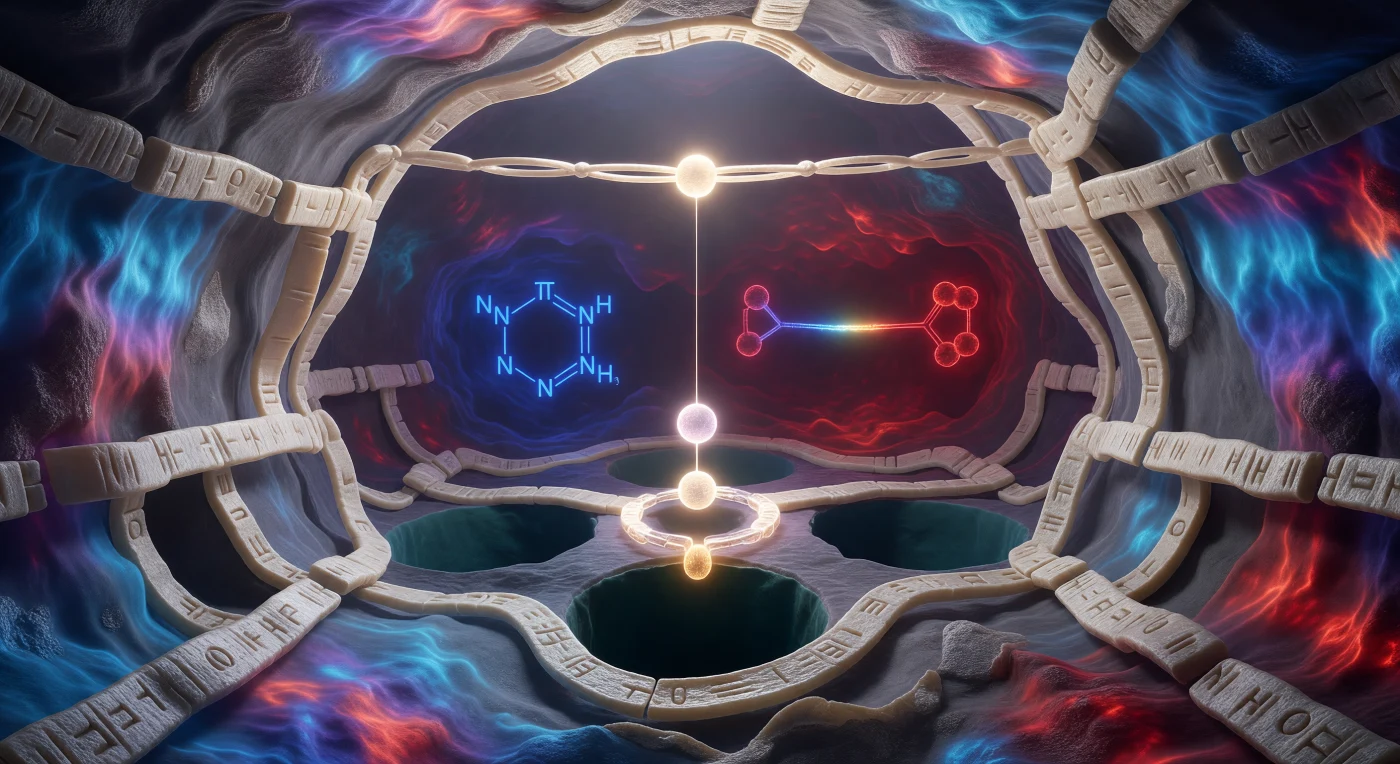
Nel cuore di questa scena sospesa nel tempo, ci si trova immersi nell'istante più fugace della chimica organica: lo stato di transizione di una reazione SN2, congelato al vertice della sua barriera energetica. L'atomo di carbonio centrale si erge come un monolite grafite in geometria pentacoordianta impossibile, con i tre idrogeni bloccati in un piano equatoriale perfetto a 120° — la simmetria trigonale bipiramide del momento di massima tensione orbitalica — mentre i legami parziali ambra-dorati pulsano al mezzo ordine di legame, né interi né spezzati, testimoni quantistici di una transizione elettronica in atto. Sul lato sinistro, la massa viola-indaco del bromo si allontana lentamente come una tempesta in ritirata, la sua nube elettronica densa e carica di densità negativa parziale che sfuma in un alone di carica dissolventesi; sul lato opposto, l'ossigeno nucleofilo avanza incandescente come rame fuso, stretto nella sua corona di molecole d'acqua cianotrasparenti che si riorganizzano in tempo reale attorno alla carica emergente, rendendo visibile la solvatazione dinamica che stabilizza il sistema polare. I doppi fantasmatici degli atomi di idrogeno — copie traslucide sospese appena fuori fase con i loro originali — rivelano la sovrapposizione quantistica del tunneling protonico, ricordando che a questa scala la posizione non è una certezza ma una distribuzione di probabilità ondulatoria. Il solvente acquoso tutt'intorno non è vuoto ma una folla densa e premente di sfere ciano e argento legate da filamenti idrogeno blu-bianchi che si tendono e si deformano sotto la redistribuzione elettrostatica irradiante dal centro, trasformando ogni molecola vicina in un sensore passivo del campo chimico in evoluzione.

Galleggiando all'interno del canale rettilineo della zeolite ZSM-5, ci si trova immersi in un corridoio ellittico di appena 5,3 × 5,6 ångström — un tunnel così stretto da sfiorare le pareti ad ogni respiro molecolare, le cui superfici sono costituite da una sequenza ininterrotta di tetraedri SiO₄ che si concatenano in ponti Si–O–Si, con i centri di silicio color grigio-argento pallido e gli atomi di ossigeno ponticellanti di un rosso cardinale vivido, a conferire alla parete una texture sinusoidalmente corrugata come le nervature interne di un tubo cristallino di ordine atomico. Ogni portale decagonale si apre davanti in prospettiva perfetta, un arco di dieci membri che risplende di un biancoazzurro freddo mentre retrocede verso un punto di fuga luminoso, e punti bianco-accecanti segnano i siti acidi di Brønsted — nodi di alluminio sostituiti al silicio le cui idrossili protoniche brillano come archi di saldatura in miniatura, reattivi e intensi in una superficie altrimenti cristallograficamente serena. A trenta ångström di distanza si apre l'incrocio molecolare: il canale sinusoidale che arriva perpendicolare spalanca un secondo tunnel ellittico nella parete laterale, creando un crocevia di soli cinque ångström attraverso cui le molecole devono negoziare il passaggio in un collo di bottiglia più stretto di una singola molecola d'acqua. Stipate nel canale in calde tonalità ambrato-dorate, le molecole idrocarburiche riempiono lo spazio a quasi perfetto contatto di van der Waals con la superficie ossidrilata del poro, le loro superfici lisce e leggermente traslucide che rispecchiano la geometria corrugata del tunnel con una precisione quasi meccanica, ciascuna molecola incastrata nel suo ospite cristallino come una chiave nella sua serratura.
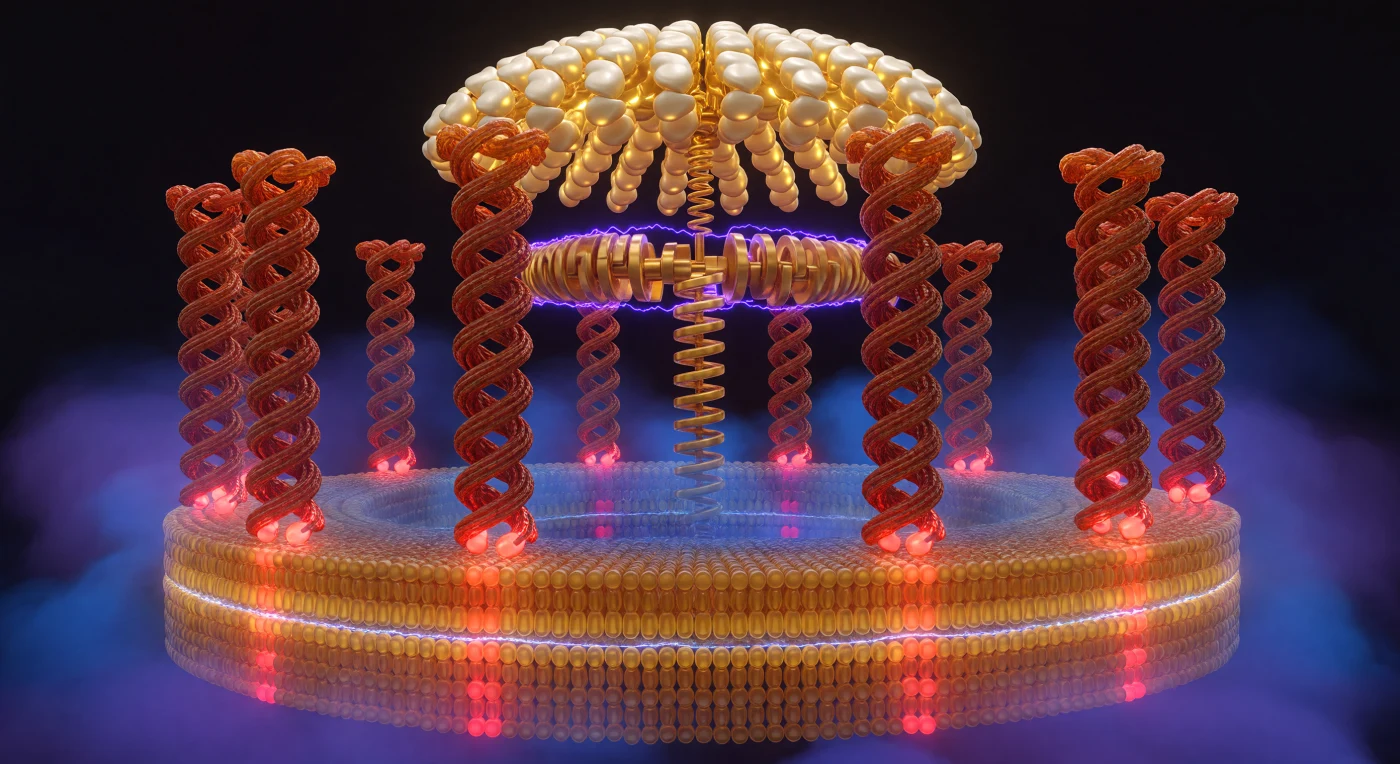
A pochi nanometri di distanza, dieci colonne elicoidali di proteina si curvano intorno all'osservatore come le pareti interne di un colosseo vivente, i loro profili terracotta e ruggine corrugati dalle spirali del backbone peptidico che costituiscono i sottodomini transmembrana della subunità c dell'ATP sintasi — una macchina molecolare che converte il flusso di protoni attraverso la membrana mitocondriale interna in legami chimici ad alta energia. Alla base di ciascuna colonna, i residui di glutammato alternano tra uno stato cremisi acceso, deprotonato e elettrostaticamente instabile nel canale protonico, e uno stato mattone spento, neutro e sepolto nel cuore idrofobico del doppio strato lipidico, un'oscillazione che scandisce la rotazione del rotore c con la cadenza di un meccanismo a scatto antico di quattro miliardi di anni. Il gradiente di forza proton-motrice di 150–200 mV non si manifesta con simboli astratti, ma nella qualità stessa della luce: cobalto e viola intensi si addensano sul versante citosolico come un'atmosfera compressa sull'orlo della scarica, mentre il versante matriciale respira un indaco più caldo e rilassato, il campo elettrostatico visibile nella densità del colore stesso. Sopra, il dominio catalitico F₁ si arcua come una volta gotica di oro pallido e avorio, i suoi sottodomini β che pulsano tra stati di legame distinti mentre l'albero a camme della subunità γ trasmette la torsione rotazionale in sintesi di ATP — un accoppiamento meccanochimico che trasforma il moto casuale di ioni in ordine molecolare impeccabile.
