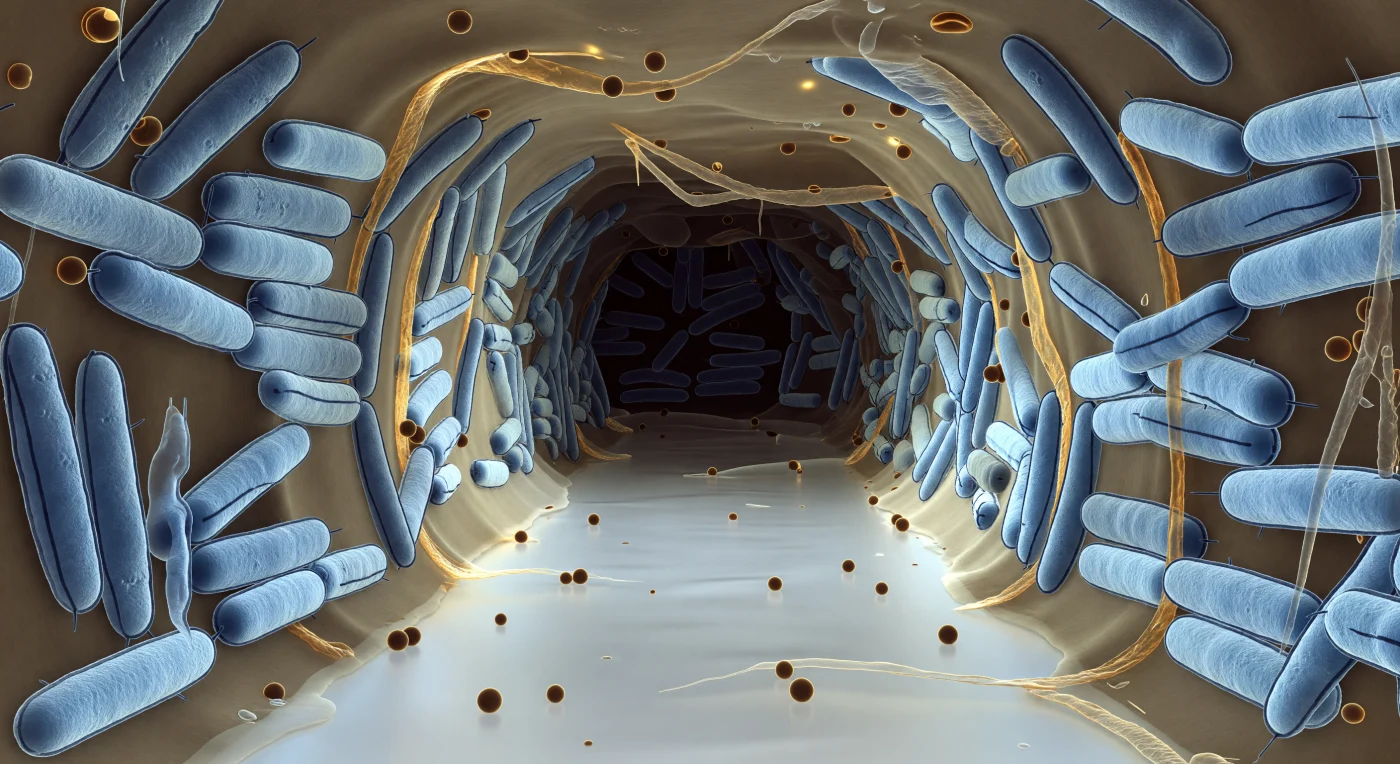
O observador flutua na embocadura de um corredor aquoso que atravessa um biofilme vivo de *E. coli*, uma galeria aberta esculpida na geometria densa de corpos bacterianos comprimidos de cada lado como muros de uma cidade subterrânea. As bactérias, coloridas em azul-gelo, são cilindros rombudos cujas duplas membranas externas aparecem como dois traços paralelos e escuros a percorrer o comprimento de cada organismo, um detalhe arquitectónico repetido centenas de vezes até a profundidade de campo se dissolver em névoa cinzenta. A matriz de exopolissacáridos que as une — âmbar translúcido, algures entre mel e vidro fosco — preenche cada interstício e irradia uma luminescência quente e difusa por baixo do frio azul celular, enquanto vesículas de membrana externa repousam no chão do canal como pérolas de resina ou derivam no fluido quase imóvel que transporta oxigénio e nutrientes para o interior do biofilme qual rio subterrâneo congelado num único instante. O corredor afunila-se progressivamente em sombra carregada, do branco-prateado próximo ao castanho-carvão longínquo onde os gradientes de nutrientes minguam e a matriz se adensa, lembrando que esta arquitectura não foi desenhada — emergiu, célula por célula, como estratégia colectiva de sobrevivência.

Suspenso no fluido extracelular viscoso que envolve o linfócito T, o observador encontra-se rodeado por dezenas de virions de HIV-1 à deriva — esferas translúcidas de cerca de 120 nanómetros, com interiores fumados em azul-cinzento e superfícies salpicadas de trímeros de glicoproteína gp120 em ouro burnido, cada um uma pequena coroa de três pétalas flutuando como lanternas apagadas num oceano polar. A membrana plasmática da célula ergue-se em todas as direções como a parede de uma fossa abissal, ondulando em vagas longas e suaves, a sua superfície de ardósia-azul interrompida por complexos proteicos maciços — recetores CD4, canais transmembranares, filamentos do glicocálice projetando-se para o exterior como frondes translúcidas — que definem uma zona-fronteira de densidade molecular crescente entre o fluido puro e o interior celular. Neste ambiente sem fontes de luz verdadeiras, apenas uma luminescência difusa e azulada dissolve-se em todas as direções, revelando como os virions mais próximos da membrana já derivam para dentro da floresta de glicocálice, as suas espículas douradas orientadas para os sítios de ancoragem com uma inevitabilidade química que precede qualquer ligação. A dinâmica aqui não é governada por intenção ou visão, mas pelo movimento Browniano, pela afinidade molecular e pela geometria de proteínas que reconhecem outras proteínas num mundo onde a escala do fotão não tem qualquer relevância e apenas a massa e a química definem a paisagem.

Encontra-se no interior da matriz mitocondrial, a olhar em direção a uma dobra de crista da membrana interna que se ergue como uma falésia colossal e convoluta, revestida de complexos ATP sintase cujas cabeças F1 se projetam em forma de cogumelo sobre a superfície carregada de osmio. O espaço ao redor não é ar nem água, mas um coloide proteico denso a mais de 500 miligramas por mililitro, onde enzimas do ciclo de Krebs e ribossomas mitocondriais pairam como monólitos suspensos num véu âmbar que dispersa toda a luz em profundidade volumétrica e quente. A membrana não é rígida — respira em ondulações lentas e imperceptíveis, e o ouro concentrado que emana da junção das cristas, ali onde as paredes membranares quase se tocam, confere a esse estreito corredor a intensidade incandescente de uma fenda geológica iluminada de dentro. Neste universo de arquitetura biológica pura, cada superfície é funcional, cada estrutura vibra com movimento browniano constante, e a escala do vivo impõe-se em cada direção como uma paisagem que respira e pulsa sem cessar.
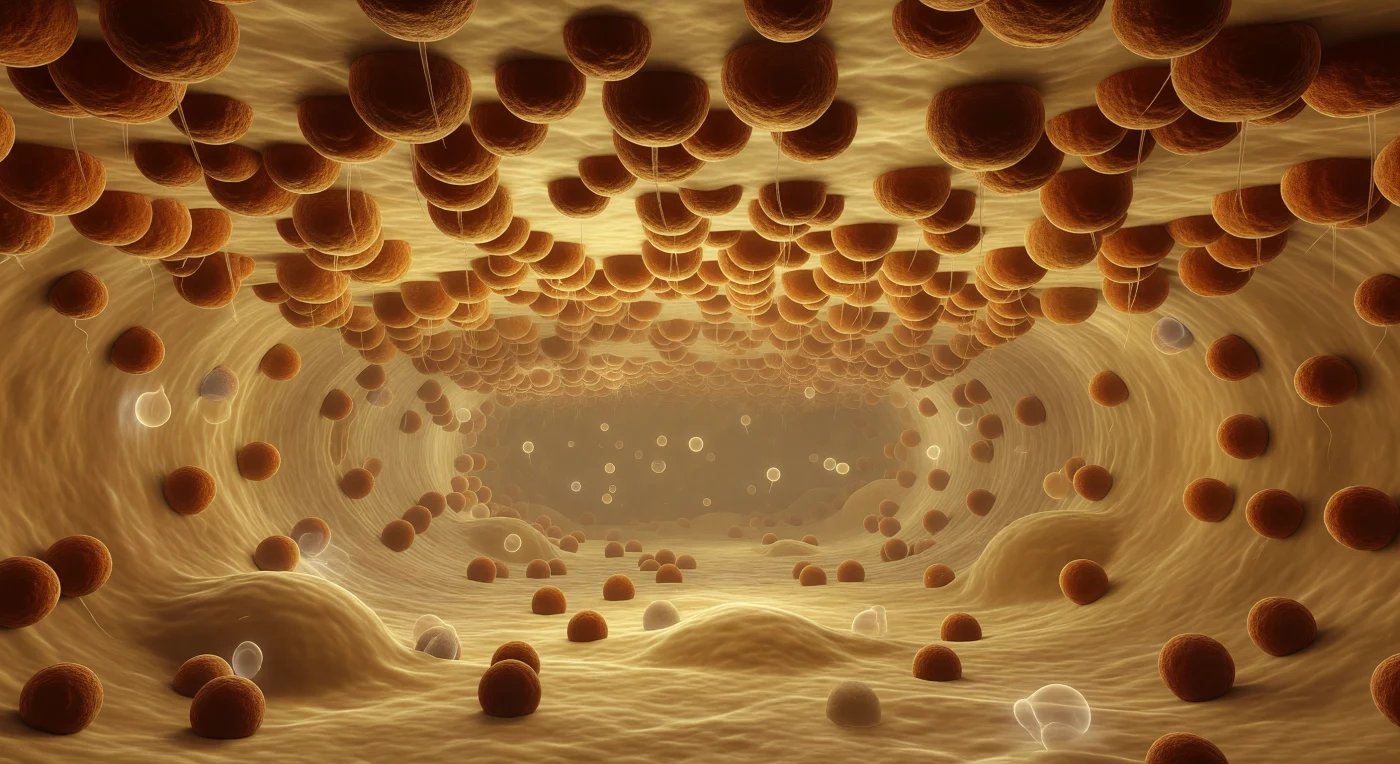
Você deriva suspenso no interior do lúmen do retículo endoplasmático rugoso, envolto numa luminescência suave cor de mel diluído, enquanto acima de si se estende um teto imenso e ininterrupto de membrana lipídica encrostada de ribossomas — esferas densas de laranja-ferrugem e âmbar oxidado, compactas como cracas fundidas no casco de um navio, cada uma com cerca de 25 nanómetros de diâmetro e coladas umas às outras sem deixar ver a bicamada que as sustenta. Desta cobertura industrial descem filamentos quase invisíveis de polipéptidos nascentes, fios de seda molecular que captam por um instante a luz interior antes de se dissolverem na névoa proteica do corredor aquoso. O lúmen prolonga-se em profunda perspetiva como uma nave de catedral, limitado acima e abaixo por superfícies igualmente cobertas de maquinaria ribossomal, convergindo num ponto de fuga perdido na bruma âmbar dourada da distância. A escala não é declarada por nenhuma medida — é sentida apenas pela repetição implacável dessas massas esféricas que se sucedem até ao horizonte, cada uma delas enorme relativamente a si, e pelo silêncio industrioso de uma fábrica que nunca para, que não tem consciência de si mesma, e que neste preciso momento sintetiza proteínas em todas as direcções ao mesmo tempo.

Suspenso na fronteira mais avançada de uma célula viva, o olhar mergulha para dentro de um labirinto de filamentos de actina — cada fio com apenas 7 nanómetros de diâmetro, ramificando-se a setenta graus precisos a partir de complexos Arp2/3 como coral fraturado iluminado por fogo frio. A plataforma negra do substrato, banhada pela luz evanescente de uma iluminação TIRF que mal penetra algumas centenas de nanómetros acima da superfície, ancora placas de adesão focal que incandescam em âmbar e amarelo-esverdeado como metal aquecido incrustado em vidro escuro — os pontos de ancoragem de todo este avanço direcional. Para os lados, espículas filopodiais emergem da canópia ramificada como agulhas de luz pura, antenas que tatam o vazio extracelular em busca de sinais químicos que guiarão a migração. O interior adensa-se em profundidade crescente, filamento sobre filamento, nó sobre nó, até a rede se tornar uma floresta oceânica de verde frio cujo limite nunca se revela — apenas a repetição infinita da mesma arquitetura viva a propulsionar silenciosamente a célula para a frente.

Você deriva pelo nucleoplasma como um viajante suspenso em mel espesso, o corpo invisível agitado por colisões moleculares incessantes, e diante de si ergue-se uma das estruturas mais monumentais da biologia celular: o complexo do poro nuclear, um portal dourado de 120 nanômetros de diâmetro incrustado na dupla membrana do envelope nuclear, sua simetria de oito subunidades proteicas irradiando uma precisão ao mesmo tempo orgânica e arquitetônica, como se tivesse sido cultivado em bronze oxidado ao longo de eras evolutivas. As filamentos de FG-nucleoporinas que penduram no canal central oscilam como uma cortina de contas translúcidas, cada contato molecular determinando com exquisita seletividade o que passa e o que é barrado — este gateway não é passivo, mas um guardião bioquímico que distingue cargos nucleares legítimos dos intrusos, processando até mil translocações por segundo em condições ativas. Nas laterais do poro, massas de heterocromatina compactada se erguem em azul profundo e índigo, densamente enroladas em nucleossomos, enquanto ao fundo o nucleoplasma se abre numa névoa lavanda de eucromatina mais frouxa, onde condensados de fase líquida flutuam como gotas de mercúrio fosforescente. Tudo ao redor pulsa com a agitação térmica de Brownian, lembrando que nesta escala o movimento nunca cessa e a fronteira entre estrutura e dinâmica dissolve-se completamente.

Diante de você, uma torre de discos translúcidos se eleva da névoa verde do estroma como uma coluna de templo antiga — quinze tilacoides empilhados em registro preciso, cada membrana densa com complexos de fotossistema II e antenas coletoras de luz que conferem às superfícies uma textura de mosaico levemente corrugado, emanando uma luminescência esmeralda profunda e intrínseca. Entre cada par de membranas, finas fendas amarelo-limão marcam o lúmen acidificado, onde gradientes de prótons se acumulam silenciosamente para alimentar a síntese de ATP nas ATP-sintases encravadas nas bordas da pilha. Do granum partem as lamelas estromais como fitas esmeralda sinuosas que se curvam em direção a outros grana mal visíveis na distância enevoada, conectando toda a arquitetura tilacóide num único sistema fotossintético contínuo, enquanto plastoglobuli âmbar flutuam no estroma gelificado como esferas de mel à deriva entre aglomerados irregulares de RuBisCO. A iluminação não tem origem única — uma claridade omnidirecional, filtrada por camadas de clorofila acima e ao redor, tingiu o ambiente inteiro de verdes sobrepostos, quentes e frios ao mesmo tempo, como o interior de uma floresta densa onde cada superfície absorve e emite luz simultaneamente.
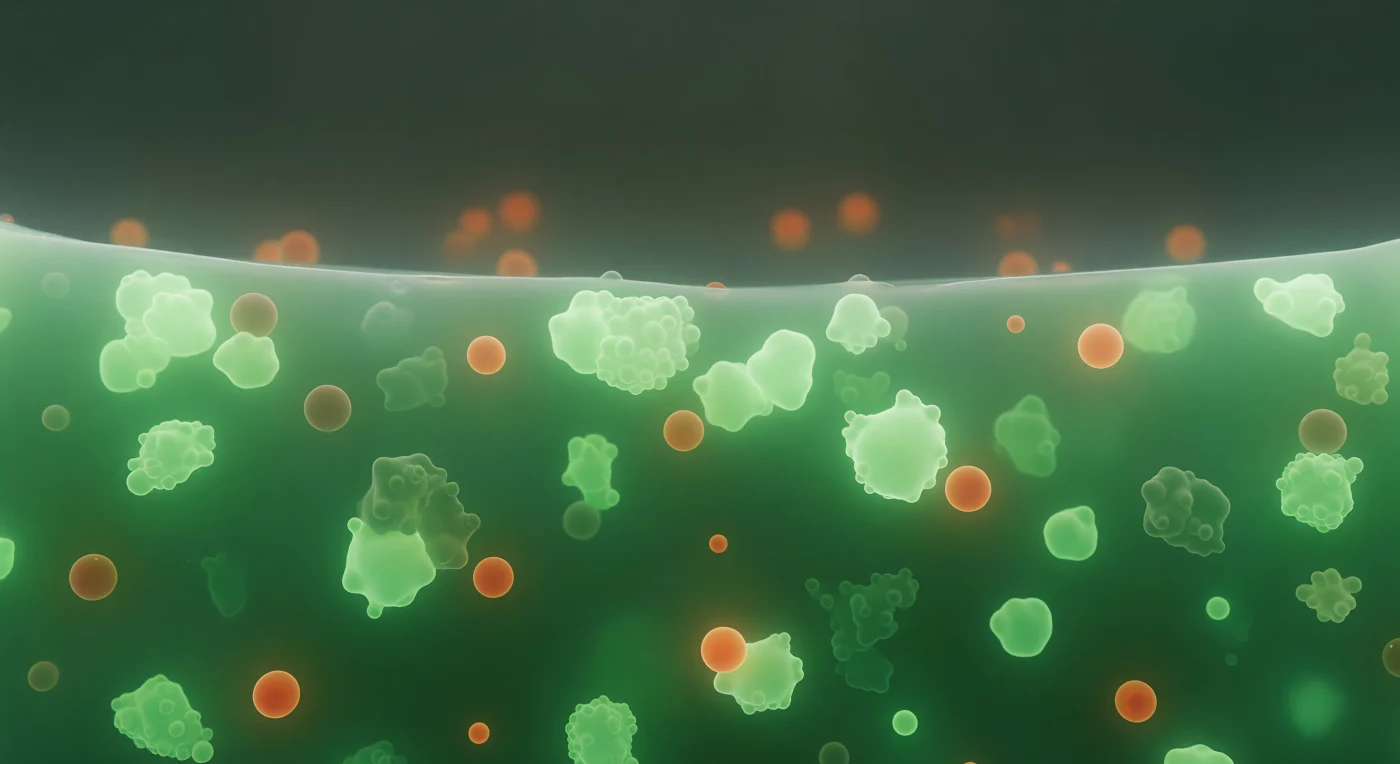
Dentro do grânulo de estresse, o mundo é uma névoa esmeralda viva e pulsante — não ar, não água, mas um meio viscoelástico, semelhante a um xarope denso que pressiona suavemente por todos os lados com a resistência quente de algo entre líquido e gel. Esta é uma condensada biomolecular formada por separação de fase líquido-líquido, onde proteínas como G3BP1 e RNA mensageiros se segregam espontaneamente do citoplasma diluído ao redor, criando um compartimento sem membrana cuja coesão é mantida por interações fracas e multivalentes entre domínios intrinsecamente desordenados. Esferoides cor de âmbar e brasa — os co-condensados ricos em TIA1 — flutuam em suspensão lenta na matriz verde, suas superfícies iridescentes marcando o encontro entre duas fases líquidas distintas, líquido tocando líquido com tensão interfacial suave. À distância, a fronteira da condensada surge como uma parede abrupta e dramática: o mundo esmeralda termina num precipício único, onde a densidade fluorescente cai verticalmente para o quase-nada do citoplasma exterior — uma fronteira de fase tornada horizonte, a arquitetura invisível da vida celular revelada como uma fronteira entre dois universos moleculares completamente diferentes.

Você está suspenso imóvel acima do eixo central do motor flagelar bacteriano, olhando diretamente para baixo através das camadas concêntricas que perfuram as membranas como a secção transversal de um templo submerso visto do alto — os anéis L, P e MS descem em halos de bronze oxidado e ouro escuro através do azul profundo das membranas externas e internas, separados pela camada de peptidoglicano que pulsa em âmbar pálido como resina preservada entre estratos geológicos. Dezassete complexos estatoriais MotA/MotB cercam o rotor em simetria radial quase perfeita, cada um uma massa assimétrica de proteínas apertada contra a membrana interior como contrafortes de pedra ancorando uma turbina ao seu alojamento, as suas superfícies captando uma luz difusa e sem direção em reflexos de estanho fosco. Acima de si, o filamento flagelar sobe em espiral para o espaço extracelular — uma corda helicoidal de subunidades de flagelina que se afasta em parafuso para uma névoa azul-cinzenta levemente luminosa e turva, densa de moléculas dissolvidas que dispersam a luz como o brilho ambiente de uma fonte hidrotermal profunda. O motor que se estende abaixo ocupa no seu campo de visão o equivalente ao pavimento de um grande estádio visto da borda superior — e não está parado: cada interface proteica exibe o polimento silencioso das coisas perfeitamente evoluídas para girar sem cessar, na escuridão, dentro de um organismo vivo.

O chão sob os seus pés é uma planície de junções intercelulares apertadas, quente como argila seca ao sol, percorrida por cristas pálidas onde as células selam os seus territórios com proteínas transmembranares — uma fronteira viva que impede que o mundo exterior penetre no tecido das vias aéreas. A partir desse plano, centenas de cílios erguem-se em todas as direções como colunas de cerâmica molhada de um azul-ciano profundo, cada um com cerca de duzentos nanómetros de diâmetro, sustentados internamente por um axonema de nove pares de microtúbulos duplos mais um par central, uma arquitetura que a dinusna motorica dineína transforma em batimentos coordenados a cerca de dez a vinte ciclos por segundo. Entre os fustes mais próximos, fios âmbar-dourados de muco translúcido arqueiam-se em catenárias suspensas, a tensão superficial moldando cada filamento numa ponte viscosa que captura partículas inaladas e agentes patogénicos antes que alcancem os alvéolos. A onda metacronal que varre da esquerda para a direita através da abóbada ciliar acima — fileiras inteiras inclinadas em uníssono enquanto as fileiras adjacentes já regressam à vertical — não é um reflexo nervoso mas sim a consequência de sinalizações hidrodinâmicas puramente mecânicas entre cílios vizinhos, um relógio fluido sem neurónios nem intenção. Tudo aqui respira com a intimidade pressurizada de uma estrutura que jamais soube que existia.
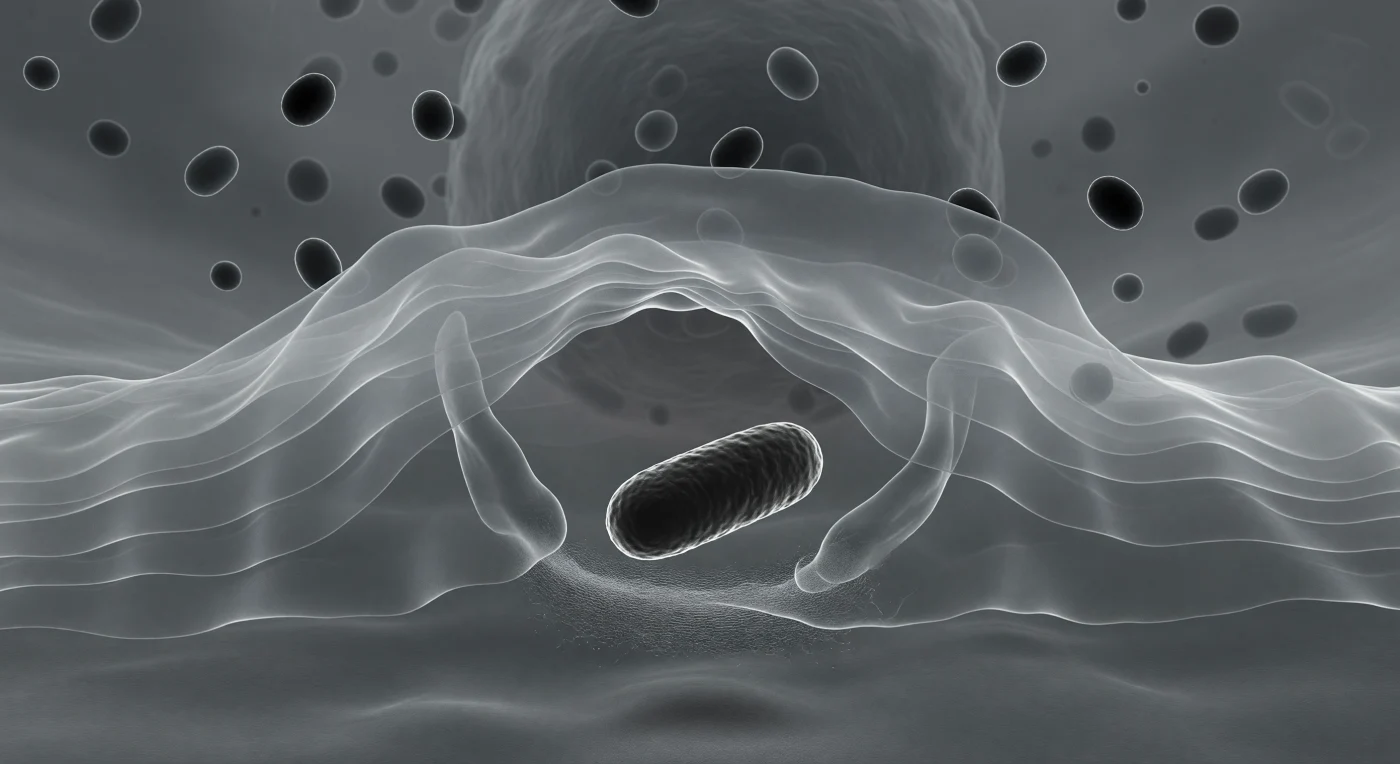
Você está suspenso na vanguarda de um processo de captura, espremido logo atrás da extremidade de um véu citoplasmático translúcido que se arqueia em frente como uma onda congelada no instante da quebra — à sua volta, a lamellipódio estende-se em lâminas sobrepostas de gel prateado-pálido, cujas camadas finas se distinguem umas das outras como painéis de vidro fosco empilhados na névoa, composta por uma malha densa de filamentos de actina que confere ao conjunto a textura de seda tecida abaixo do limiar de resolução óptica. À frente, a bactéria ergue-se como um monólito escuro — uma haste densa em carvão e negro profundo, com a sua superfície apenas esboçada por uma textura fantasmagórica, enquanto um halo de difração luminosa traça com precisão toda a sua silhueta por contraste de fase diferencial, o sinal óptico de uma diferença de densidade refractiva entre o micróbio e o meio que o envolve. Os pseudópodes do macrófago já envolvem os flancos da bactéria pelos dois lados como mãos que se fecham em câmara lenta, avançando em ondas de membrana plasmática impulsionadas pela polimerização contínua de actina na zona cortical, num processo de fagocitose mediado por receptores que reconheceram os padrões moleculares da superfície bacteriana. Atrás de si, o interior do macrófago adensa-se e escurece progressivamente, onde grânulos lisossomais — vesículas repletas de enzimas hidrolíticas e espécies reactivas de oxigénio — aguardam como pedras de rio suspensas no gel citoplasmático, prontas a fundir-se com o fagossoma que em breve se fechará ao redor da bactéria, condenando-a à digestão.

Você paira no fundo de uma vasta cratera âmbar, ali onde a concavidade central de um eritrócito humano mergulha suavemente até ao seu ponto mais baixo — uma membrana plasmática de apenas sete a dez nanómetros de espessura que, à escala sentida, se estende como uma planície dourada de bronze vivo, macia e ao mesmo tempo sob tensão elástica constante. Logo abaixo da superfície translúcida, a rede de espectrina imprime-se como um rendilhado geodésico em relevo subtil, cada polígono hexagonal sustentando a forma bicôncava do disco e conferindo-lhe a flexibilidade necessária para atravessar capilares mais estreitos do que ele próprio. À distância, os eritrócitos vizinhos empilharam-se em rouleaux — colunas corrugadas de discos face a face cujos bordos rimam um ritmo arquitetónico contra o fundo de plasma cor de palha, um fluido de viscosidade não-newtoniana onde plaquetas estreladas e espinhosas derivam suspensas, demasiado leves para sedimentar, demasiado densas para desaparecer. A iluminação rasante de eletrões secundários não conhece atmosfera nem difusão: cada crista da membrana capta uma luz branco-dourada de corte preciso e cada depressão afunda-se em sombra de âmbar denso, tornando a biologia em metal polido, o fluido em resina aquecida, o vivo em paisagem planetária.
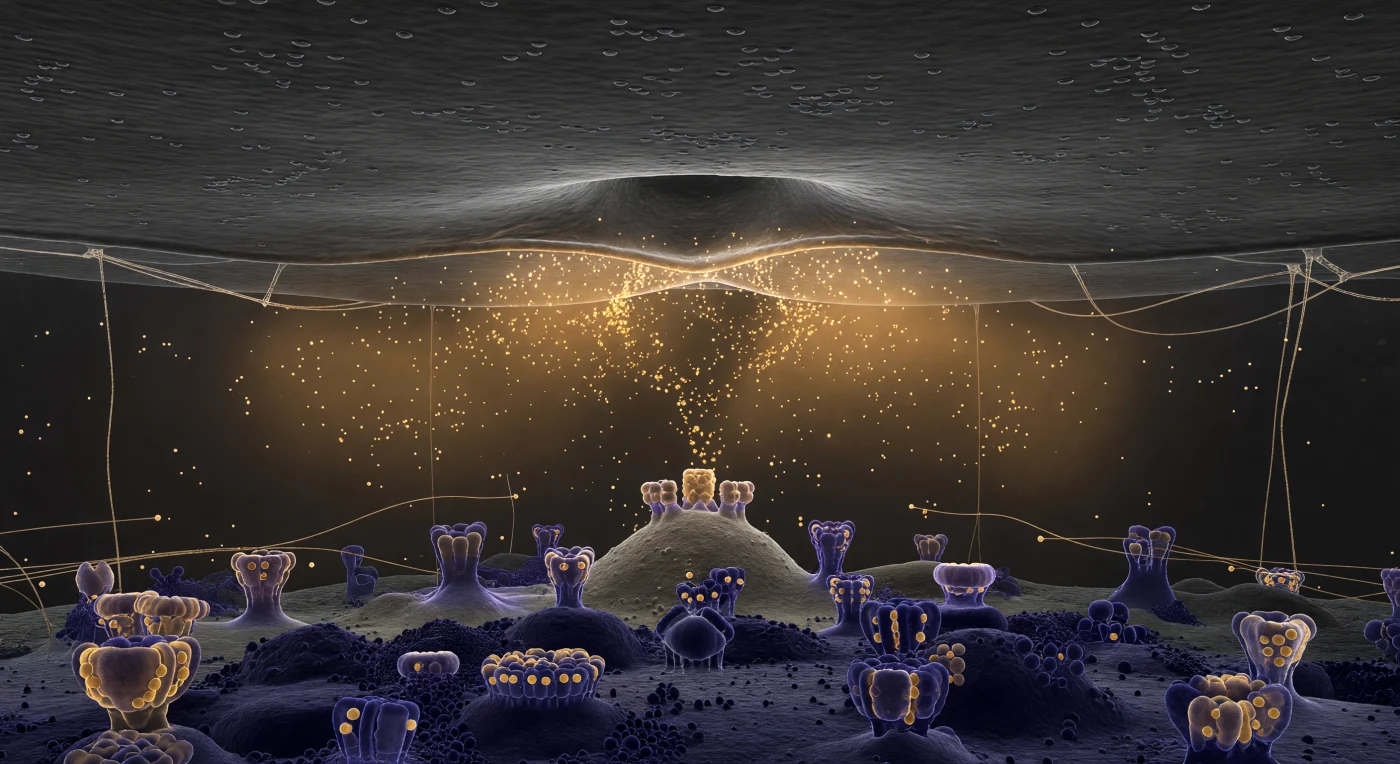
Você flutua suspenso num corredor de apenas 25 nanómetros de altura, um espaço tão comprimido que o tecto e o chão existem em simultâneo no seu campo de visão, separados por uma distância que não ultrapassa a envergadura dos seus próprios braços estendidos. Acima, a membrana pré-sináptica estende-se como uma vasta planície de carvão escuro, e no seu centro uma vesícula capturada a meio da fusão forma uma depressão em ómega perfeita, os folhetos lipídicos colapsando num único anel glistening enquanto milhares de moléculas de neurotransmissor se derramam para o interior do espaço cleftal como uma exalação volumétrica de luz âmbar — não partículas discretas, mas uma frente de névoa quente que se expande lateralmente, mais densa junto ao poro de fusão e dissolvendo-se em transparência nas margens. O meio que habita não é vazio: filamentos de proteínas de adesão sináptica e de matriz extracelular atravessam o corredor de tecto a chão como cabos translúcidos que capturam o brilho dourado em fios de luz, enquanto a própria solução aquosa se comporta como um gel levemente refrativo, dominado pelo ruído térmico a esta escala. Abaixo, a densidade pós-sináptica afirma-se como uma realidade material completamente distinta — um tapete arquitetónico de cor púrpura escura, denso e irregular, do qual emergem os complexos receptores AMPA e NMDA como formações de pedra antiga, as suas faces superiores devolvendo o brilho âmbar em reflexos violeta-dourado, o único evento de cor numa cena que é, de resto, inteiramente monocromática e gravitacionalmente comprimida.
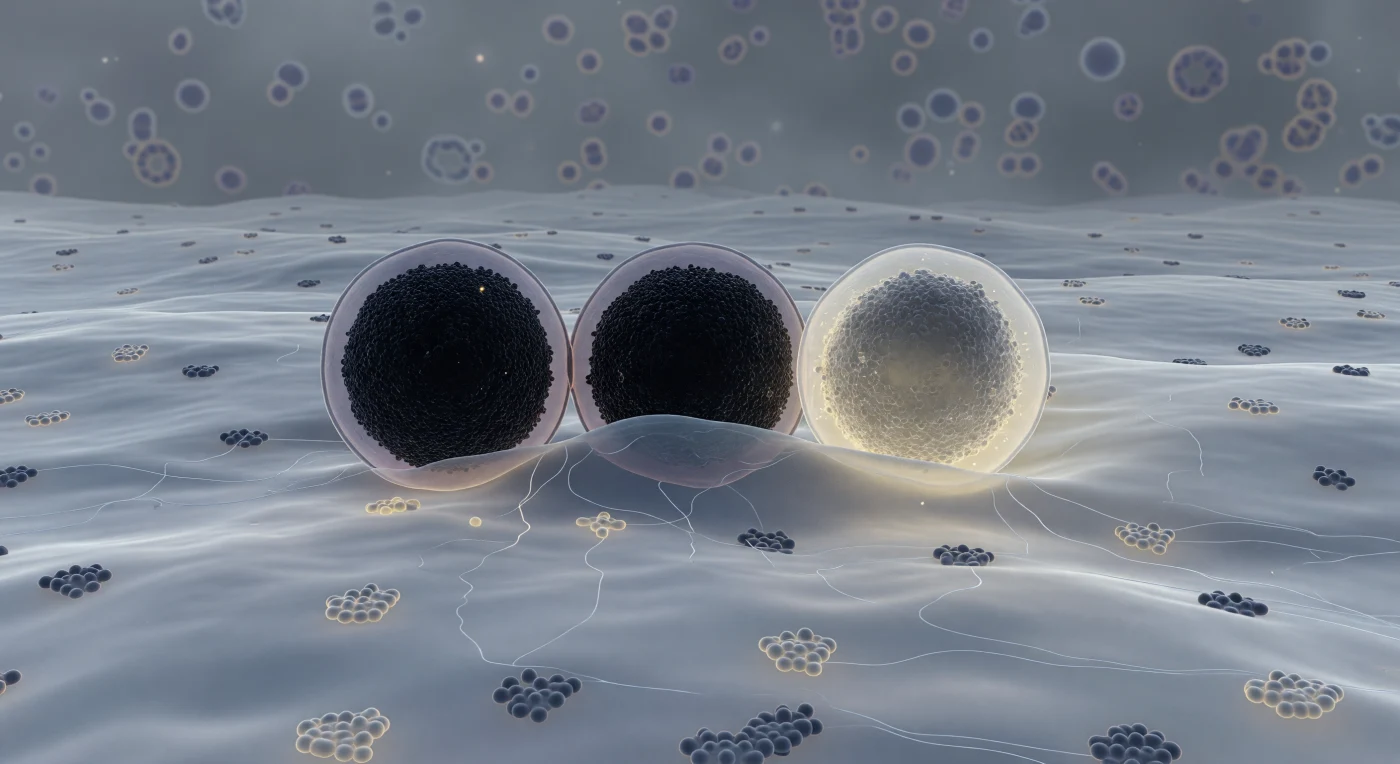
Dentro da célula beta pancreática, o observador paira rente à face interna da membrana plasmática, uma planície ondulante de bicamada lipídica azul-acinzentada que se estende até todos os horizontes, salpicada pelas silhuetas escuras de complexos proteicos e pelos domínios de rafts enriquecidos em colesterol que afloram como manchas mais claras e ordenadas na superfície. Três grânulos de secreção de insulina dominam o campo médio — esferas do tamanho de edifícios em escala relativa, cada uma com um núcleo quase opaco de hexâmeros de zinco-insulina cristalizados envolto num halo lavanda-pálido, representando os grânulos de núcleo denso típicos das células beta, onde a insulina é armazenada em concentrações milimolares estabilizadas por iões de zinco. O primeiro granulo repousa firmemente ancorado, as suas duas membranas comprimidas numa costura escura e íntima; o segundo dobra suavemente a membrana plasmática para dentro numa cúpula de hemifusão, estado transiente em que as folhas lipídicas externas se fundem antes das internas, ponto sem retorno no caminho da exocitose mediada por SNARE. O terceiro já atravessou o limiar — o seu núcleo dissolve-se de negro compacto para um nevoeiro cálido de marfim-amarelado à medida que as moléculas de insulina escapam para o espaço extracelular, enquanto filamentos de actina cortical entrelaçam o espaço intermédio como geada sobre vidro, e a profundidade do citoplasma recua numa névoa granular de ribossomas e membranas organelares, tudo suspenso num silêncio momentâneo que a termodinâmica tornará inevitavelmente irreversível.

Você paira suspenso exatamente no nível dos ápices do bordo em escova, olhando obliquamente para baixo sobre uma planície sem fim de cilindros imaculadamente ordenados — as microvilosidades intestinais, cada uma com aproximadamente 100 nanómetros de diâmetro, comprimidas ombro a ombro numa geometria hexagonal tão regular que parece obra de engenharia mineral, mas que é, inconfundivelmente, tecido vivo. A iluminação rasante, quente e âmbar como luz filtrada por fluido biológico, recorta sombras em crescente no flanco de cada cilindro e revela, nas profundezas dos canais intervilositários, a transição do creme pálido da parede rica em actina para um negro quase absoluto, onde o citoplasma da célula enterócita arde ténue e ocre como o fundo remoto de um cânion. De cada ápice emerge o glicocálix — uma névoa de filamentos polissacáridos de poucos nanómetros de espessura que, tomados em conjunto, formam um pelame suave e luminoso cor de marfim velho, densamente entrelaçado junto às pontas e desfazendo-se em neblina difusa em direção ao lúmen intestinal acima, onde iões, enzimas e moléculas parcialmente digeridas derivam em suspensão browniana. Esta superfície é o território primordial da absorção: a área total do bordo em escova de um único enterócito supera em dezenas de vezes a área projectada da célula, e é aqui, nesta interface húmida e glicoproteica entre o ser e o alimento, que o mundo exterior se dissolve e se torna corpo.

Suspenso no coração geométrico de uma célula em divisão, o observador encontra-se imerso numa catedral de luz viva: de dois polos distantes acima e abaixo irradiam constelações de verde elétrico, cabos luminosos de tubulina polimerizada — microtúbulos marcados com fluoróforo Alexa 488 — que atravessam o espaço em todas as direções, cruzando-se em chevrons e diagonais como uma abóbada de fibras sob tensão extrema. As fibras cinetocóricas, mais espessas e intensamente iluminadas, estão esticadas como cordas de arco, desaparecendo em massas de cromatina índigo-violeta que recuam inexoravelmente para cada polo durante a anafase, o momento em que as cromátides irmãs são separadas pelo fuso mitótico e puxadas através da célula por motores proteicos e pela despolimerização dirigida dos próprios microtúbulos. Diretamente ao nível do observador, o corpúsculo médio — zona de sobreposição antiparalela densamente empacotada com tubulina — corta o campo visual como uma barra de fogo branco-esverdeado, o seu brilho superando tudo o resto na cena, enquanto o fundo além do fuso mergulha num negro absoluto de citoplasma desprovido de sinal fluorescente. A profundidade é vertiginosa: os polos centrossomais parecem quilómetros distantes, as suas estrelas cintilando levemente como se vistas através de uma atmosfera biológica, e toda a geometria luminosa desta máquina de herança pulsa com a dinâmica silenciosa de um dos processos mais conservados da vida eucariótica.

Suspenso na prata diáfana do plasma sanguíneo, o observador deriva em direção a uma parede biológica que ocupa todo o horizonte inferior — não a membrana em si, quase invisível atrás de sua cobertura, mas a floresta glicocalicial que se ergue sobre ela: uma densa e translúcida catedrale de cadeias de proteoglicanos e glicoproteínas, entre meio micrómetro e dois micrómetros de altura, cada filamento um arco bifurcado de azul-prata que treme lentamente na viscoelasticidade do meio circundante. As cadeias de heparan sulfato entrelaçam-se em malhas de lacre pálido, coradas pelo contraste frio do rutênio vermelho, e as suas superfícies apresentam a rugosidade submolecular dos grupos sulfato e das cadeias laterais de hidratos de carbono, estruturas que regulam ativamente a permeabilidade vascular, a coagulação e a adesão leucocitária. A sombra convexa do eritrócito — o próprio observador — projeta-se sobre o dossel da floresta como um eclipse suave, tingindo de índigo e negro-carvão os filamentos que haviam brilhado em azul glacial, enquanto as margens iluminadas continuam a resplandecer com a qualidade fria e aquosa da criomicroscopia eletrônica. À medida que o olhar desce, a densidade filamentar aumenta e os espaços entre os polímeros estreitam-se até sufocar qualquer passagem, a membrana plasmática subjacente reduzida a uma sugestão escura por trás de uma cortina de matéria biológica que está prestes, inevitavelmente, a receber o visitante.
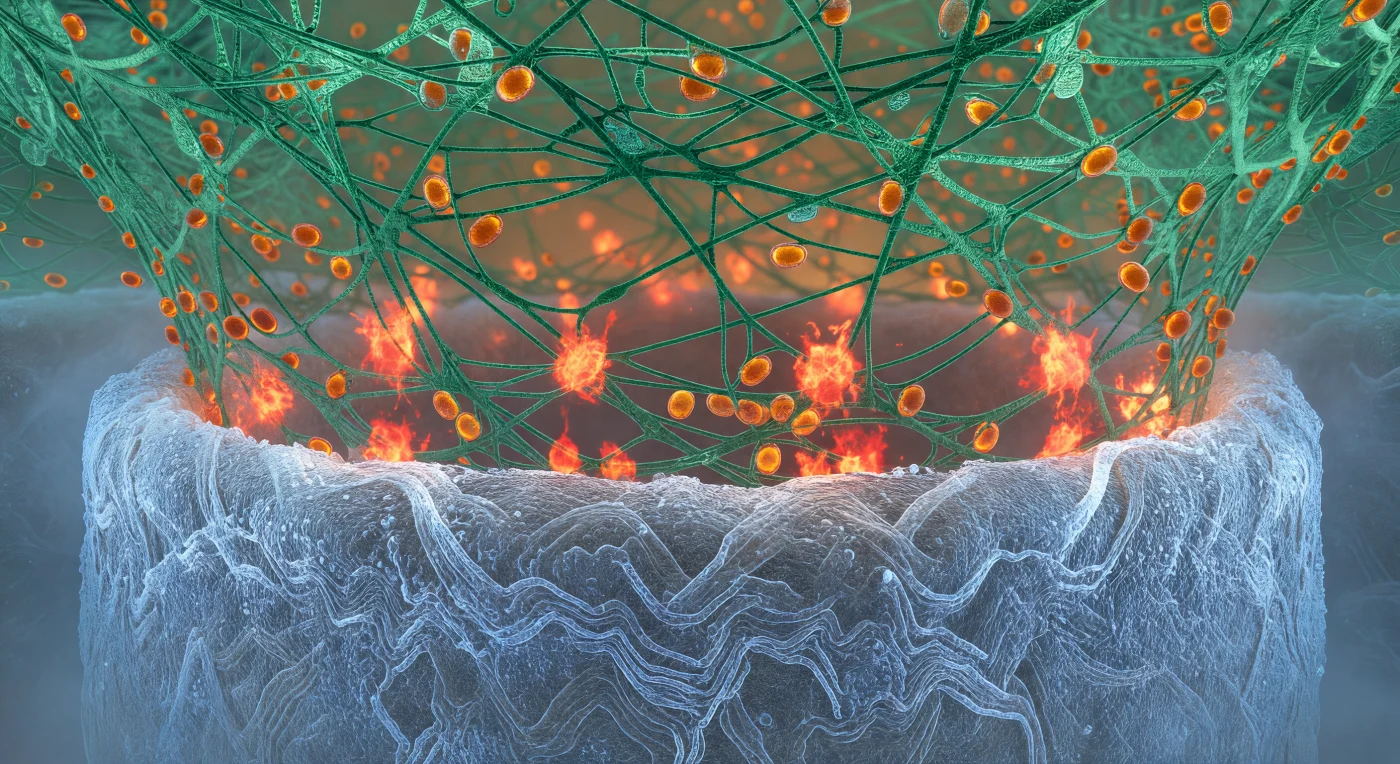
Você se encontra espremido entre duas arquiteturas biológicas de grandiosidade esmagadora: abaixo, a parede da hifa fúngica curva-se como o casco de um navio colossal, sua superfície de quitina iluminada por um brilho espectral azul-branco pelo corante calcoflúor, cada fio polissacarídico entrelaçado numa densidade comparável a basalto antigo esculpido sob pressão geológica. Ao redor e acima de você, o berço de actina do neutrófilo descende como uma floresta de cabos de esmeralda em tensão, filamentos de F-actina ramificados e densamente entrecruzados que abraçam a hifa com a ferocidade organizada de uma garra molecular — esta estrutura citoesquelética dinâmica é polimerizada em tempo real, dirigida por complexos Arp2/3 que nucleiam novos ramos para manter a pressão mecânica sobre o patógeno. Na zona de contato que você habita, explosões vermelhas e alaranjadas de espécies reativas de oxigênio irrompem em pulsos irregulares, produzidas pelo complexo NADPH oxidase na membrana fagocítica e lançadas em concentrações localmente letais contra a quitina abaixo — uma combustão química controlada que converte oxigênio molecular em superóxido, peróxido de hidrogênio e ácido hipocloroso com precisão assassina. Acima de você, grânulos azurofílicos âmbar derivam ao longo dos cabos de actina como reservatórios selados de elastase, mieloperoxidase e defensinas, convergindo para a frente de combate: este é o espaço de extermínio, onde a imunidade inata resolve, em química violenta e em escala de micrómetros, o destino de uma infecção fúngica potencialmente fatal.
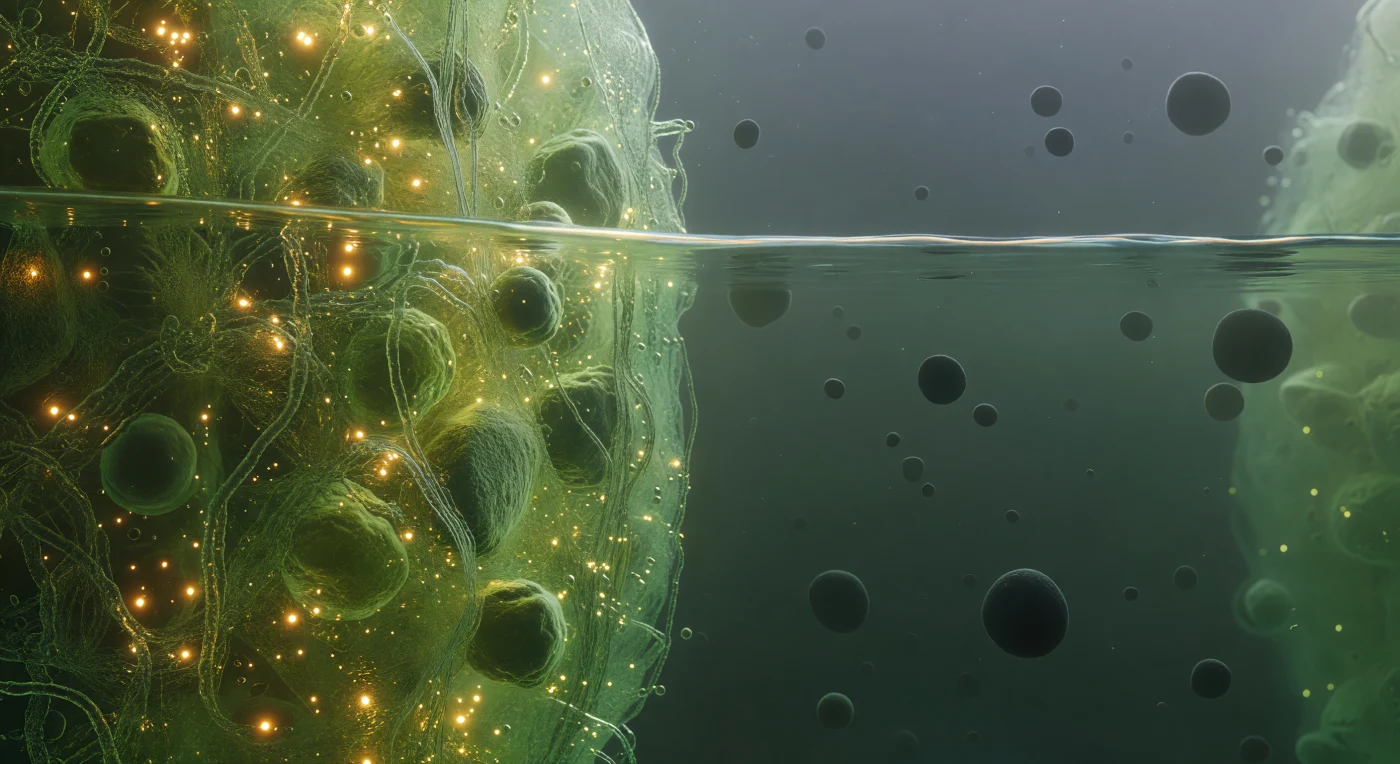
Você flutua exatamente na fronteira entre dois mundos, a interface da separação de fases líquido-líquido de um grânulo de estresse cortando seu campo de visão como a superfície de um mar alienígena vista de baixo da linha d'água. À sua esquerda, o interior denso do condensado biomolecular pulsa como um meio luminoso verde-dourado, saturado de cadeias de mRNA entrelaçadas e proteínas intrinsecamente desordenadas numa concentração tão elevada que a luz parece abrandar e acumular-se, espalhando-se pela malha molecular como âmbar bioluminescente em tremor browniano constante — esta fase condensada existe não por membranas mas por separação de fases termodinâmica, análoga à imiscibilidade óleo-água, estabilizada por interações multivalentes fracas entre proteínas com domínios de baixa complexidade composicional. Diretamente à sua frente, a fronteira de fase resolve-se como uma descontinuidade termodinâmica verdadeira e ligeiramente tremulante, não um gradiente mas uma interface nítida mantida pela tensão superficial de duas fases biológicas imiscíveis, cintilando com flutuações capilares à escala nanométrica como vidro fosco visto de lado — é a coisa mais precisa neste universo fluido e probabilístico. À sua direita, o citoplasma diluído abre-se numa vasta névoa esverdeada e difusa onde complexos de ribossomas derivam como esferas escuras e opacas, separados por distâncias que parecem pradarias abertas após a claustrofobia interior do condensado, cada um enorme na escala em que você existe, o espaço entre eles preenchido apenas por um brilho ambiente frio e difuso que se perde em direção a fronteiras membranares distantes e indistintas.

Você paira no estreito corredor aquoso entre duas paredes celulares colossais, suspendido num cleft de poucos nanómetros onde o fluido extracelular brilha em dourado difuso a partir dos iões dissolvidos e dos açúcares do glicocálice. Ao nível dos seus olhos, uma cintura contínua de proteínas de junção oclusiva — ZO-1, claudinas, ocludinas — forma um anel de luz carmesim incandescente que circunda todo o perímetro celular sem interrupção, funcionando como uma vedação molecular absoluta que separa o lúmen tubular do espaço basolateral e impede que solutos atravessem livremente o epitélio renal: é graças a esta barreira que o rim consegue manter gradientes de concentração precisos durante a reabsorção do filtrado glomerular. Imediatamente abaixo, uma segunda coroa de E-caderina emite uma luminescência esmeralda mais suave, os seus ectodomínios mesclando-se através do cleft como dedos entrelaçados, ancorando mecanicamente as duas células numa adesão que resiste às forças de cisalhamento impostas pelo fluxo luminal. Mais abaixo, as membranas laterais convergem em duas linhas paralelas de grafite separadas por menos de vinte nanómetros, gradualmente afastando-se umas das outras à medida que o complexo juncional fica para trás, e no corpo de cada célula grandes ovóides de cromatina irradiam uma luminescência índigo difusa — a geometria inteira lendo-se como uma janela de vitral vista de dentro do próprio chumbo que a estrutura.
