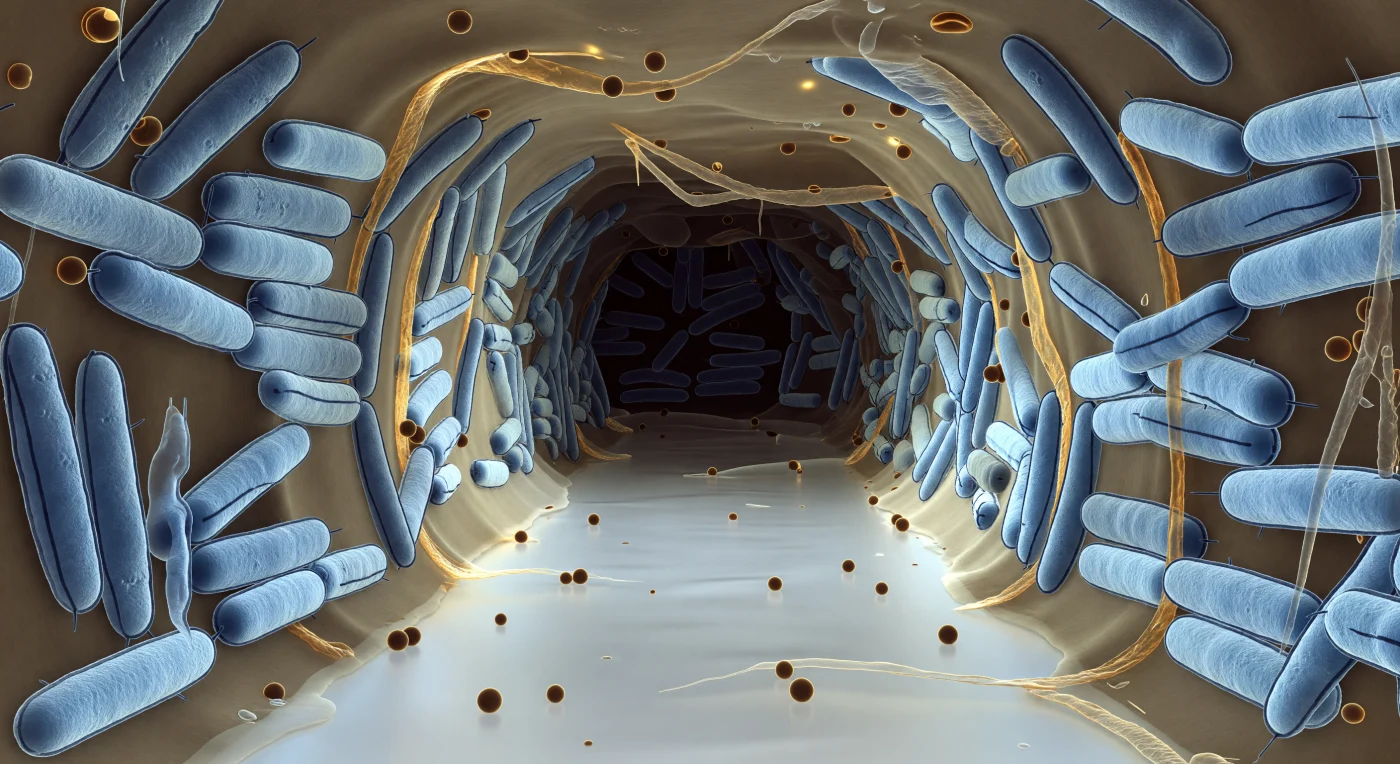
Je bevindt je aan de ingang van een waterkanaal dat dwars door een levend biofilm loopt — een lichtgevende, nagenoeg stille corridor begrensd door dicht opeengepakte *E. coli*-staven die als ijsblauwe cilinders oprijzen, elk vijf keer zo hoog als jijzelf, hun dubbele buitenmembranen zichtbaar als twee fijne donkere lijnen die zich honderden malen herhalen tot diep in de scherptediepte voordat ze vervagen in blauwig grijs duister. Tussen de bacteriën vult een doorschijnende, amber-bruine exopolysaccharidematrix elk tussenruimte op — een mengeling van honing en bevroren glas die de wanden een sluimerend warm schijnsel geeft, terwijl bolvormige buitenmembraanvesicles als amberkleurige kralen over de kanaalboden drijven of half ingebed rusten in de overhangende EPS-vouwen. Dit kanaal is geen lege doorgang maar een actieve transportader: opgelost zuurstof en nutriënten stromen inwaarts langs een gradiënt die geleidelijk uitdooft naar het verre, donkere einde van de corridor, waar de matrix dichter wordt en de chemische activiteit van het collectief terugvalt tot een fluistering. Fijne pilusdraden strekken zich uit van sommige celoppervlakken de vloeistofkolom in als tastende draadjes, en de EPS zelf trekt samen tot gedraaide strengen tussen naburige cellen om vervolgens te verdunnen tot een nauwelijks zichtbaar vlies langs de kanaalrand — een architectuur die niet is ontworpen, maar is ontstaan uit miljoenen chemische onderhandelingen tegelijk.

Je hangt roerloos in de extracellulaire vloeistof vlak buiten de membraanwand van een T-lymfocyt, omringd door tientallen HIV-1-virionen die in een trage Brownse suspensie ronddrijven — elk een wazige blauwgrijze bol van amper 120 nanometer doorsnede, bezaaid met gouden gp120-gp41-trimeren die als kleine drielobbige kronen op hun envelop zijn gezet. De vloeistof zelf is traag en lichtdoorlateend, doortrokken van opgeloste eiwitten en glycoproteïnen die op enige afstand oplossen in een parelachtige waas, alsof je zweeft in ijskoud poolwater waar helderheid overgaat in ondoordringbaar schemerduister. Achter je en aan alle kanten rijst het plasmamembraan op als de wand van een oceanische kloof — golvend, leien-blauw, bezet met enorme CD4-receptoren en glycocalyxdraden die als transparante fransen uitsteken en in het diffuse cryo-licht een zilveren schijnsel opvangen. De virionen die het dichtst bij de membraanwand zijn gedreven, buigen licht van vorm waar hun envelop de glycocalyx raakt, hun gouden spikes reikend naar ankerpunten met een geometrische onvermijdelijkheid die eerder chemie dan intentie verraadt. Dit is een wereld zonder schaduw en zonder lichtrichting, wherein massa en moleculaire affiniteit de enige landschapsvormende krachten zijn.

Je staat op de bodem van een kolossaal canyon-systeem waarvan de wanden geen steen zijn maar membraan — de binnenste mitochondriale membraan die zich in geweldige, kronkelige plooien aan alle kanten opheft, elke richel tientallen lichaamslengten omhoogklimmend voor ze terugkrult in de schemering. Het oppervlak van deze kliffen is diep houtskoolbruin, de kleur van osmiumgekleurde lipide-dubbellagen in cryo-EM-warmte, en over elk denkbaar stukje van dat oppervlak steken de ATP-synthase-complexen uit: duizenden gedrongen paddestoeltorens met ronde F1-koppen die naar buiten knikken in de matrix, verankerd in het membraan als palen in een donkere, bewegende kust. De matrix waarin je staat is geen lucht en geen water, maar een dichte, amberkleurige colloid vol eiwitten — meer dan vijfhonderd milligram per milliliter — die elk foton verstrooien tot een zacht, honingkleurig gloeien, terwijl reusachtige TCA-cyclus-enzymen als half-opgeloste keien opdoemen en mitochondriale ribosomen als donkere monolito's trillen in een Brownse beving zo constant dat hij aanvoelt als stilte. Ver voor je vernauwt de cristae-junctie zich tot een strakke, goudverlichte slotcanyon, de doorgang bijna dichtgemetseld door dicht opeengepakte ATP-synthase-koppen die het warme licht terugkaatsen in zachte highlights — een wereld van puur biologische architectuur, ademend, overvol en levend in iedere richting.
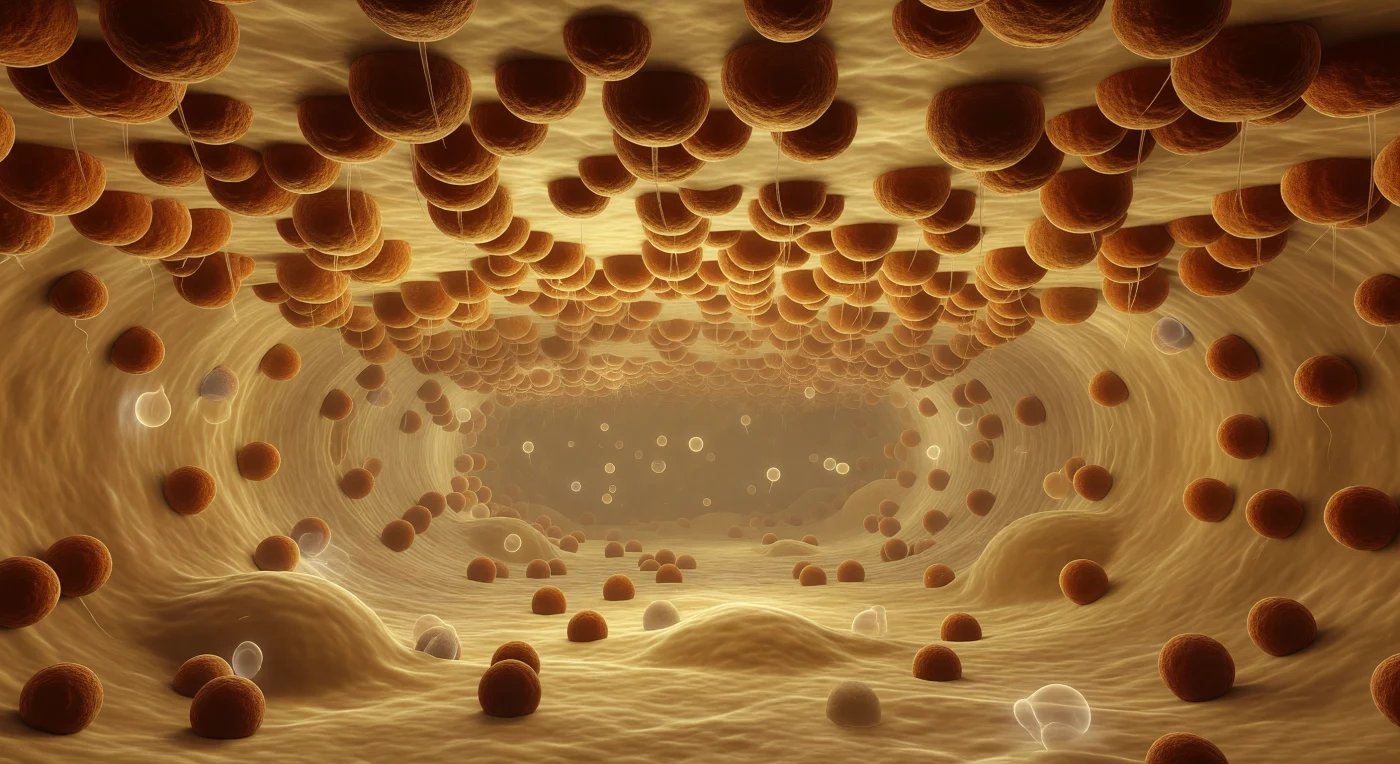
Je drijft in de holte van het ruw endoplasmatisch reticulum, omgeven door een warm, goudgeel glanzende ruimte vol opgeloste eiwitten die als trage, doorschijnende schaduwen langs je glijden. Boven je strekt zich een ononderbroken plafond van lipidedubbellaag uit, volledig bedekt met ribosomen — roestbruine, korrelige bollen van elk zo'n 25 nanometer doorsnede, zo dicht opeengepakt als mosselen op een havendam, zodat de membraanoppervlakte eronder volledig aan het zicht onttrokken is. Vanuit elk ribosoom daalt een nauwelijks zichtbare draad van een ontluikend polypeptide neer in de lumenruimte, opvlammerend in een vluchtige iriserende glinstering voordat hij oplost in de eiwitrijke nevel van de corridor. De gang zelf strekt zich in perspectief voor je uit als een kathedraalgang, begrensd boven en onder door ribosoombestudeerde membraanwanden die samenkomen in een verre, amberkleurige waas — een fabriekshal zonder pauze, zonder bewustzijn, waarin de cel onafgebroken en massaal haar eiwitten aanmaakt.

Je zweeft aan de uiterste grens van een levende cel, op het scherpste punt van het lamellipodium, terwijl een TIRF-verlichtingsgolf het obsidiaan-zwarte substraat van onderaf beschijnt met een koud blauw-wit schijnsel dat nauwelijks enkele honderden nanometers de hoogte in reikt. Vanuit dit donkere vlak rijst een verbijsterend vertakt netwerk van actinefilamenten omhoog — elk draad slechts zeven nanometer dik, oplopend in elektrisch teal-groen en branching op precieze zeventig-graden hoeken bij de Arp2/3-complexen die als koraalpoliepen in het halfduister pulsen. Op het substraat eronder ankeren focale adhesieplaques als gloeiend amber-goud ingebed in koud glas, de warme halos die ze uitstralen bloedend door het koele filamentenwoud omhoog en zo de enige oriëntatiepunten vormend in een wereld die anders grenzeloos fractaal is. Aan de flanken prikken filopodiale uitsteeksels de extracellulaire leegte in als chirurgische naalden van zuiver groen licht — elk een bundel parallelle actinefilamenten die de nog onverkende ruimte aftasten, hun uiteinden wegdovend tot een muisgloei tegen het absolute donker buiten de celgrens. Schaal bestaat hier niet als afstand maar als herhaling: elk vertakkingspunt identiek, elk filament een spiegelbeeld van het volgende, totdat het oog begrijpt dat dit levende bos zich oneindig naar binnen voortzet als de architectuur van gerichte beweging zelf.

Je zweeft door de dichte, gelachtige nucleoplasma en nadert een van de meest verfijnde structuren in de cel: het kernporcomplex, dat zich als een massaal gouden rozet voor je uitstrekt, precies 120 nanometer breed, ingebed in de donkere, houtskoolkleurige dubbele kernmembraan die zich aan weerszijden uitstrekt als twee parallelle gewelven van oud metaal. Acht eiwitsubcomplexen in perfecte radiale symmetrie vormen een bronzen portaal waarvan de bogen samenkomen in een centraal kanaal, behangen met halfDoorzichtige FG-nucleoporinefilamenten die als een parelengordijn zacht golven en flikkeren telkens wanneer een molecuul erdoorheen breekt. Aan weerszijden van de porie rijzen compacte heterochromatinmassa's op in diepe indigoblauw en middernachtsblauw, ruw van oppervlak door de nucleosoomkralen die het omgevingslicht absorberen in plaats van weerkaatsen, terwijl achter je het euchromatiné losser en lichtvoller wordt in een lavendelgrijze nevel van sijdezachte chromatinedraden. Dit complex reguleert met architecturale precisie het verkeer van tienduizenden moleculen per seconde tussen cytoplasma en celkern — geen mechanische poort, maar een levend, selectief filter dat de moleculaire identiteit van elk doorgangsmolecuul bewaakt.

Voor je rijst een toren van jade — vijftien thylakoidmembranen op elkaar gestapeld met de precisie van een levend bouwwerk, elke schijf bezaaid met duizenden fotosysteem II-complexen en lichtoogstende eiwitten die in de vloeibare lipidedubbellaag zijn ingebed en een zacht smaragdgroen licht lijken uit te stralen. Tussen de membranen scheiden dunne citroengelе lumengaten de schijven als lichtzuigende voegen, gevuld met een zuur vloeistof doorlopen door een protongradiënt die de synthese van ATP in het hele chloroplast aandrijft. Vanuit de toren strekken stromale thylakoidlamellen zich horizontaal uit als groen kronkelende linten die naburige grana verbinden in een samenenhangend netwerk van membraankanalen, terwijl de stroma zelf aanvoelt als een opaalachtig gel, vol met RuBisCO-enzymen die als bleke keien elke kijkrichting bezetten en amberkleurige plastoglobuli die als druppels honing in het diffuse groene licht drijven. De architectuur van dit geheel — membraan na membraan, toren na toren — is de materiële uitdrukking van fotosynthese: de omzetting van lichtkvanten in chemische energie, hier te aanschouwen als structuur.
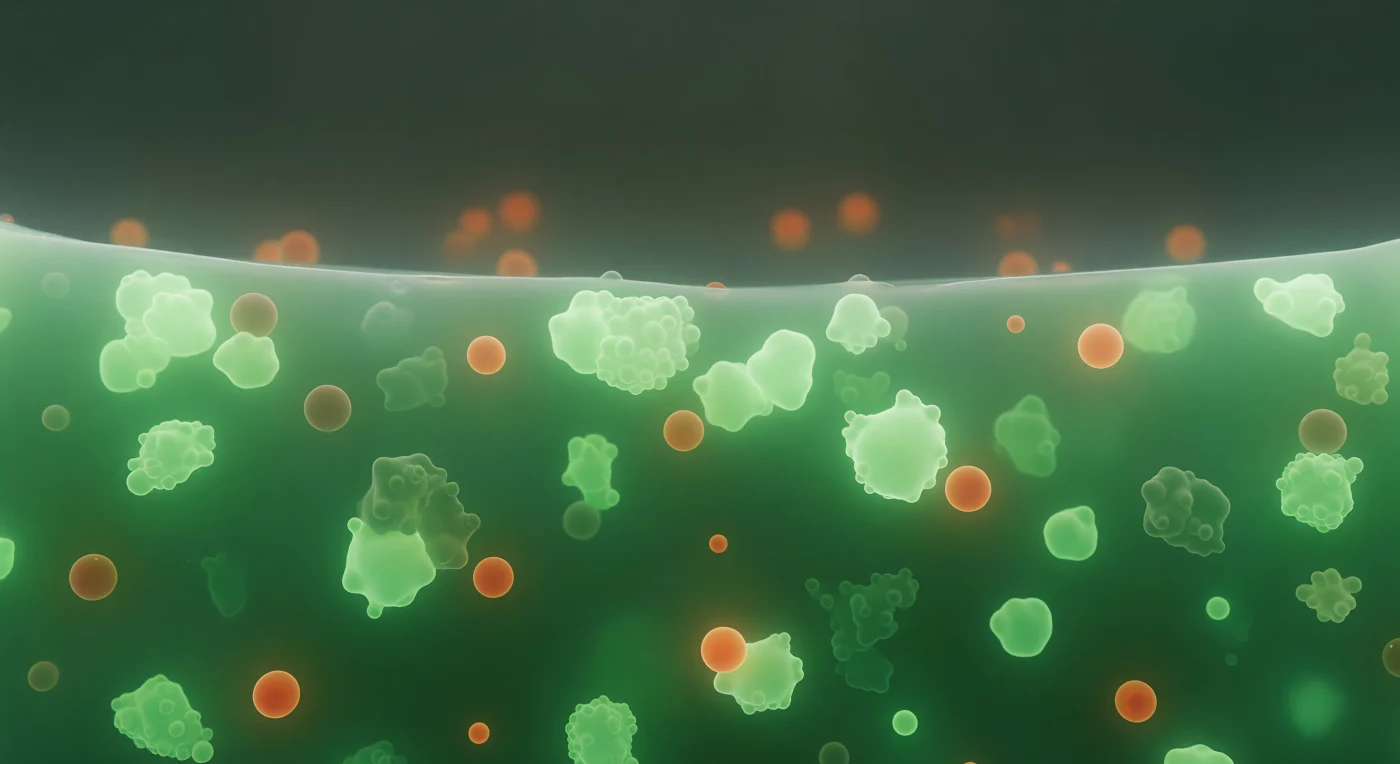
Je bevindt je in het hart van een stressgranule, een levende druppel die de cel aanmaakt wanneer ze in nood is — een tijdelijk toevluchtsoord voor mRNA en eiwitten die wachten op betere tijden. Om je heen pulseert een diepe, smaragdgroene nevel, het gecondenseerde netwerk van G3BP1-eiwitten dat fluorescerend oplicht met een gelijkmatige, diffuse intensiteit die geen enkel middelpunt heeft maar overal vandaan lijkt te komen, als een wereld die zichzelf verlicht vanuit haar eigen moleculaire massa. Dit medium is geen vloeistof en geen vaste stof, maar iets daartussenin: een viscoelastisch condensaat dat zich gedraagt als een extreem langzaam vloeiende gel, bijeengehouden door zwakke, veelvuldige wisselwerkingen tussen RNA-bindende eiwitten die voortdurend aan elkaar hechten en loslaten in een dynamisch evenwicht. Dichter bij je drijven warme, barnsteenkleurige bollen van TIA1-rijke co-condensaten, vloeibare eilanden binnen een vloeibaar oceaan, gescheiden door fasegrenzen die trillen van interfaciale spanning — twee fases in coëxistentie, elk met zijn eigen moleculaire samenstelling, elk stabiel door de thermodynamica van fasescheiding. En aan de rand van deze wereld eindigt alles abrupt: een steil, glinsterend grensvlak waar de dichte groene gloed met één enkele stap oplost in de dunne, bijna lege cytoplasma daarbuiten, de horizon tussen twee volledig verschillende vormen van moleculair bestaan.

Je zweeft roerloos boven de centrale as van de bacteriële flagellairmotor, en kijkt recht omlaag in een van de meest verfijnde mechanische structuren die de levende natuur heeft voortgebracht. Onder je dalen concentrische ringen — de L-, P- en MS-ringen — als de doorsnede van een antieke tempel neer door gelaagde membranen: de buitenste ring glinstert als koud brons in het lipopolysaccharide van de buitenmembraan, de peptidoglycaanlaag gloeit amberkleurig daartussen, en het MS-ringcomplex vult het binnenste membraanvlak met geoxideerd goud en gehard staal, zijn oppervlak geribbeld met eiwitgrenzen die het licht vangen als fijne schubben. Zeventien statorcomoplexen van MotA en MotB verdringen zich in bijna perfecte radiaalsymmetrie rondom de rotor, elk een asymmetrische massa die als een machtig steunbeer tegen de binnenmembraan drukt — samen vormen ze een ringvormig array van moleculaire machines dat oogt als de cilinderwand van een turbine op industriële schaal. Boven je schroefdraait het flagellaire filament als een lang, gelijkmatig spiralend kabel het extracellulaire milieu in, terwijl de omringende ruimte gevuld is met een zwak lumineuze, troebele suspensie van opgeloste moleculen die het licht verstrooien tot een diffuus blauwgrijs waas — want zelfs de leegte hier is niet leeg, maar doordrenkt van de chemische drukte die dit geheel onophoudelijk laat draaien.

De blik richt zich omhoog vanuit de vloer van een luchtwegepitheel: honderden gladde, diepblauwe ciliaschachten van elk slechts 200 nanometer in doorsnede rijzen als zuilen van een ondergedompelde kathedraal recht de hoogte in, hun oppervlakken vaag oplichtend van binnenuit alsof het glas is dat koude gloed vasthoudt. Tussen de naastgelegen schachten hangen ambergouden slijmdraden in trage, gravitationele bogen — doorschijnende bruggen van mucus die zich versmallen tot haardunne filamenten waar ze aan de ciliumwand verankeren, elk een catenairekoord van organisch hars gespannen in een onmogelijk intiem schip. Boven, waar de tips van de cilia samendringen tot een golvend baldakijn, is een metachroonse golf in één bevroren ogenblik gevangen: een hele colonnade buigt in unisono opzij alsof een onzichtbare wind erdoorheen strijkt, terwijl de aangrenzende rij alweer rechtop staat — het ritme dat mucus en deeltjes richting de keel vervoert, in steen gehouwen als een reliëf van opeenvolgende beweging. Onder de voeten strekt zich een gewarm oker-beige vlakte van strak aaneengesloten celbegrenzing uit, de nauwe verbindingen als gedroogd klei-aardewerk op een rivierbedding, bezaaid met rondborstige microvillistompjes die nauwelijks groter zijn dan de ribbels van de axonemale doubletparen die als zijdedraad door de ciliumwand drukken.
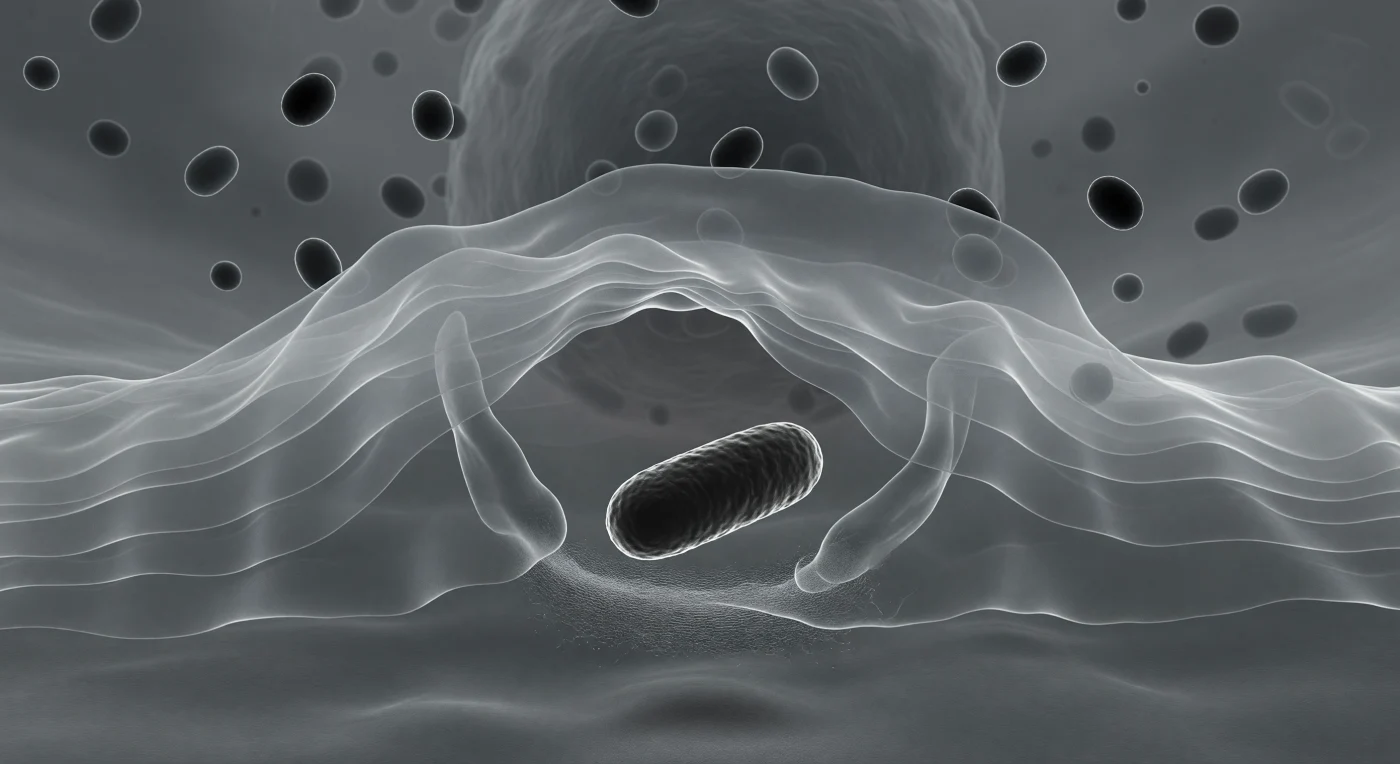
Je bevindt je op de allervoorste rand van een fagocyterende macrofaag, ingeklemd achter een doorschijnend cytoplasmatisch vlies dat zich als een bevroren golf om een staafvormige bacterie sluit — de lamellopodium strekt zich voor je uit in gelaagde, zilverachtige vliezen van actinerijk gel, elk stratum nauwelijks te onderscheiden van het volgende, als gestapelde panelen bevroren mist. Direct voor je domineert de bacterie het gezichtsveld als een donker monoliet van antraciet en zwart, haar omtrek verlicht door een heldere diffractiekrans die haar maakt tot een gloeiend silhouet tegen het grijze medium — terwijl de pseudopodiën van de macrofaag zich al langs haar flanken sluiten in een langzame, onvermijdelijke omhelzing, aangedreven door de constante polymerisatie van actinefilamenten die samen een weefsel vormen te fijn om individueel te zien maar tastbaar als geweven zijde. Achter je verdikt het cytoplasma zich en worden lysosomale granula zichtbaar als donkere, ovale vormen die traag in de gelachtige matrix zweven, elk met een subtiele lichtcorona, als stenen in suspensie — de chemische arsenalen die wachten op de gevangene die elk moment volledig ingesloten zal zijn. Het hele tafereel, weergegeven in de koude, richtingloze belichting van differentieel interferentiecontrast, spreekt van een mechanisch doel op cellulair niveau: een systeem dat al honderden miljoenen jaren bestaat, dat elke indringer herkent, omsluit en vernietigt met de architectonische precisie van een levend organisme dat zijn grenzen bewaakt.

Je zweeft boven het diepste punt van een immens amberkleurig bekken — de centrale instulping van een menselijke rode bloedcel — waarvan het membraanoppervlak zich in alle richtingen uitstrekt als de bodem van een reusachtige inslagkrater op een warm, okerkleurig wereld. Vlak onder dat trillende, translucente goud tekent het spectrine-cytoskelet zich af als een nauwelijks zichtbaar hexagonaal netwerk, geperst tegen de binnenkant van het membraan als een geodetisch patroon in gehamerd bladgoud, de elastische spanning van dit eiwitrooster verantwoordelijk voor de uitzonderlijke vervormbaarheid waarmee rode bloedcellen zich door haarvaten van minder dan vijf micrometer weten te persen. Op middellange afstand torenen stapels rode bloedcellen als monolithische zandstenen zuilen boven het strogele plasma uit — de zogenoemde rouleaux, gevormd door fibrinogeenbruggen die de cellen als munten op elkaar doen stapelen — terwijl stervormige bloedplaatjes met hun stekelige uitsteeksels verspreid tussen die celkolommen hangen, enorm ten opzichte van het membraanweefsel onder je voeten maar nietig naast de gewelfde celwanden die hen omringen. Het scherpe, schuinse licht treft elk membraankammetje zonder atmosferische verstrooiing, werpt harde schaduwen in de concave holten en geeft het hele landschap de tastbare, gegoten kwaliteit van biologisch brons — een wereld die tegelijk intiem en planetair aanvoelt, opgebouwd uit niets dan levend oppervlak en warm, secreet lichaamsvocht.
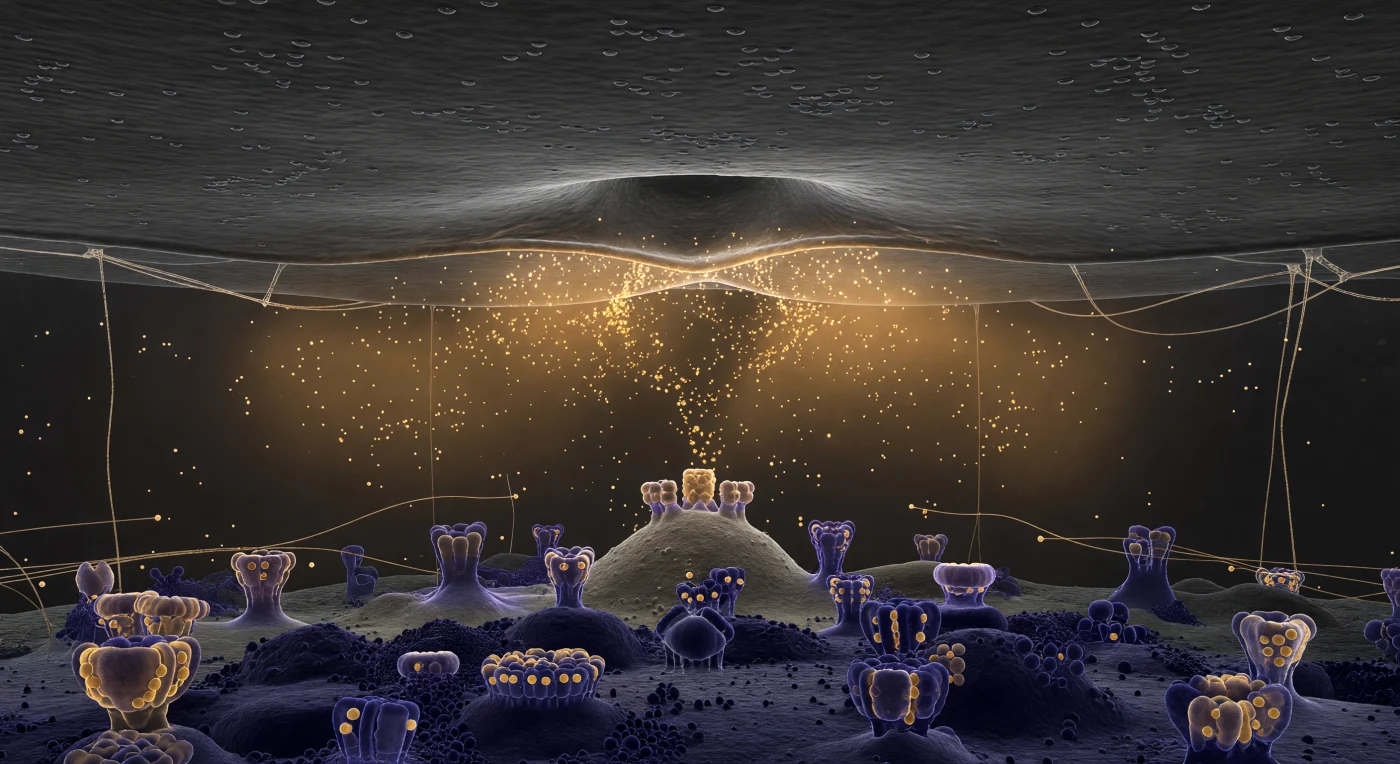
Je zweeft in een corridor zo smal dat plafond en vloer gelijktijdig aanwezig zijn — de presynaptische membraan boven je strekt zich uit als een uitgestrekte, donkergrijze vlakte, bezaaid met de nauwelijks zichtbare reliëfs van ingebedde eiwitten en lipide-microdomeinen, terwijl precies boven je een solitair blaasje in het moment van fusie is bevroren: zijn lipide-dubbellaag smelt samen met het membraanplafond in een perfecte omega-vorm, de twee bilayers versmeltend tot één doorlopend vel. Vanuit die fusieporie stroomt een warm, amber licht naar buiten — geen afzonderlijke moleculen maar een diffuse, volumetrische golf van duizenden neurotransmitters tegelijk, een zachte vloed van goud-oranje nevel die lateraal door de spleet drijft en uitdunt naar de randen, daar waar dunne strengen synaptische adhesiemoleculen en extracellulaire matrixfilamenten als doorschijnende kabels van plafond naar vloer lopen en de ambergloed opvangen in fijne gouden haarlijnen. Beneden kondigt de postsynaptische dichtheid zich aan als iets wezenlijk anders: een architecturaal landschap van donkerpaars, elektronendicht steiger werk, onregelmatig en dicht opeengepakt, waaruit AMPA- en NMDA-receptorcomplexen als oude steenformaties omhoog steken en de ambergloed van de neurotransmitters terugkaatsen in zwakke violet-gouden schitteringen op hun bovenzijde. Deze wereld is in wezen monochroom — houtskoolgrijs plafond, paarse vloer — maar de amberkleurige vloed is de enige kleurgebeurtenis, een enkele warme uitbarsting van chemie in een anders grauwgekleurde, samengedrukte ruimte van slechts vijfentwintig nanometer hoog.
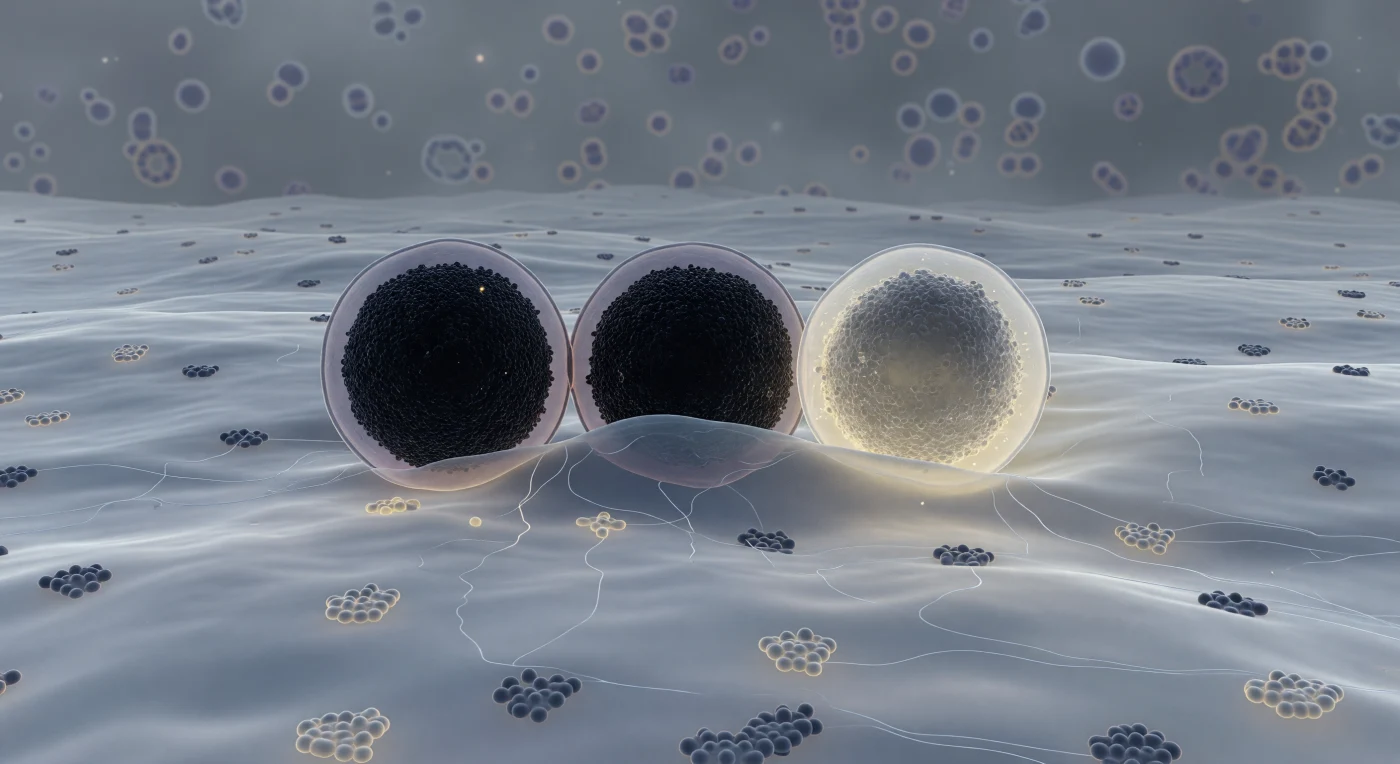
Je zweeft vlak boven het binnenoppervlak van de plasmamembraan van een pancreatische betacel, een onafzienbaar, zachtgolvend landschap van blauwgrijs lipide dat zich tot aan elke horizon uitstrekt, bezaaid met de donkere silhouetten van ingebedde eiwitcomplexen en iriserende cholesterolrijke raftdomeinen. Drie reusachtige, bolvormige insulinegranules domineren het middenplan — elk zo groot als een klein gebouw op jouw schaal — en bevinden zich in opeenvolgende stadia van exocytose: de eerste drukt zijn membraan in nauwkeurig contact tegen de plasmamembraan, zijn kern bijna ondoorzichtig zwart van kristallijn zink-insuline; de tweede vervormt de membraan eronder tot een brede, ondiepe deuk waar twee bilayers beginnen samen te vloeien in een hemi-gefuseerde kraag van onbestemde grijstint. De derde granule heeft de grens al overschreden — zijn kern lost op van pikzwart via houtskoolgrijs naar een warme, ivoor-gele nevel terwijl zink-insulinemoleculen uiteenwaaieren in de extracellulaire ruimte, een lichtgevende amberkleurige leegte die aanvoelt als open zee. Tussen de granules door slingeren ijle actinefilamenten als rijpsporen op glas, terwijl het achterliggende cytoplasma wegvalt in een dicht, macromoleculair gesluierd diepte — dit is exocytose gevangen in één onherroepelijk, bevroren ademtocht.

Je zweeft op de exacte grens waar de cel ophoudt en het leven van de darm begint: de toppen van duizenden microvilli vormen een onafzienbaar geometrisch veld van cilindrische schachten van elk slechts honderd nanometer breed, zo dicht opeen gepakt als een honingraat, hun wanden lichtdoorlatend als geperst was en subtiel gestreept door de onderliggende actinebundels die elke microvillus zijn rigiditeit geven. Schuin invallend licht werpt precieze halvemaanvormige schaduwen langs de lijzijde van elke cilinder, zodat de tussenliggende smalle kanaaltjes donker en vochtig lijken, gevuld met een eiwitrijke vloeistof waarin voedingsstoffen in trage Brownse suspensie drijven voor actief transport door de enterocyt eronder. Van elke punt rijst de glycocalyx op als een zachte ivoorkleurige waas — onvertakte polysacharidefilamenten van slechts enkele nanometers dik, die het harde geometrische ritme hieronder verhullen en samensmelten tot een continu levend vilt dat de scheiding vormt tussen cel en darmlumen. Deze haargrens is geen passieve laag: de glycocalyx herbergt digestieve enzymen, herkent moleculen, filtert en concentreert substraten, en trilt voortdurend mee in de biologische onrust die dit bevroren moment nauwelijks kan verhullen.

Je zweeft precies in het middelpunt van een delende cel, opgehangen in de equatoriale vlakte terwijl anafase op zijn hoogtepunt raast om je heen — en de wereld die je omringt is een kathedraal van levend licht. Vanuit twee polen hoog boven en diep onder je stralen tientallen microtubuli als kolossale lichtende kabels in elektrisch groen naar je toe, elk een cilinder van gepolymeriseerd tubuline van immense dikte op deze schaal, hun oppervlakken fijn gegroefd en doordrenkt met de koude jadekleur van Alexa 488-fluorochroom. De kinetochoorfibers spannen strak als boogsnaren van staal, en ze trekken boven en onder je de blauwviolette chromosomenmassa's meedogenloos poowaarts weg, terwijl de interpolaire microtubuli elkaar dwars door jouw positie kruisen als een driedimensionaal rooster van fosforescerende diagonalen. Precies op jouw hoogte doorsnijdt het midlichaam het beeldveld als een balk van puur wit-groen vuur — een antiparallelle overlapzone zo dicht gepakt met tubuline dat ze alles om haar heen overstraalt, haar centrale donkere lijn nauwelijks zichtbaar in de gloed. Buiten de spil heerst absolute zwarte stilte van het cytoplasma, niet leeg maar dicht van macromoleculaire drukte, waartegen de aureolen van melkachtig groene mist aan de randen van de microtubuli zacht vervagen — dit is geen architectuur van glas of metaal, maar van warm, dynamisch eiwit in beweging.

Je drijft door het bleekzilveren medium van bloedplasma richting een immense biologische wand die de volledige onderste helft van je blikveld inneemt: het glycocalyx, een kathedraal-dicht woud van proteoglycanen en glycoproteïnen dat als blauwe zilveren filamentbossen oprijst uit het endotheel van het bloedvatwand. De individuele heparansulfaatketens — elk slechts nanometers breed maar gezamenlijk een laag van een halve tot twee micrometer dik vormend — vertakken zich als kale winterberken op extreme vergroting, hun oppervlakken trillend van gebonden water en elektrostatische lading, lateraal verbonden door gossamerachtige kruisbruggen die het geheel de kwaliteit geven van een lichtgevende, viskeuze nevel die vast is geworden. Diep onder dit levende weefsel is het plasmamembraan van de endotheelcel amper zichtbaar: een donkere antracietlijn die in gefragmenteerde flarden door het nagenoeg ondoordringbare polymeergordijn schimert, zoals een stormdonkere oceaanbodem door een bosluifel heen. Boven je werpt je eigen schaduw — de brede convexe koepel van de naderende rode bloedcel — een zachte eclips over het glycocalyxdak, die ijsblauwe strengen in indigo en bijna-zwart verdiept, terwijl de verlichte randen van het filamentenwoud oplichten in het koude, ionisch gekleurde blauw van een rutheniumrood-TEM-kleurschema.
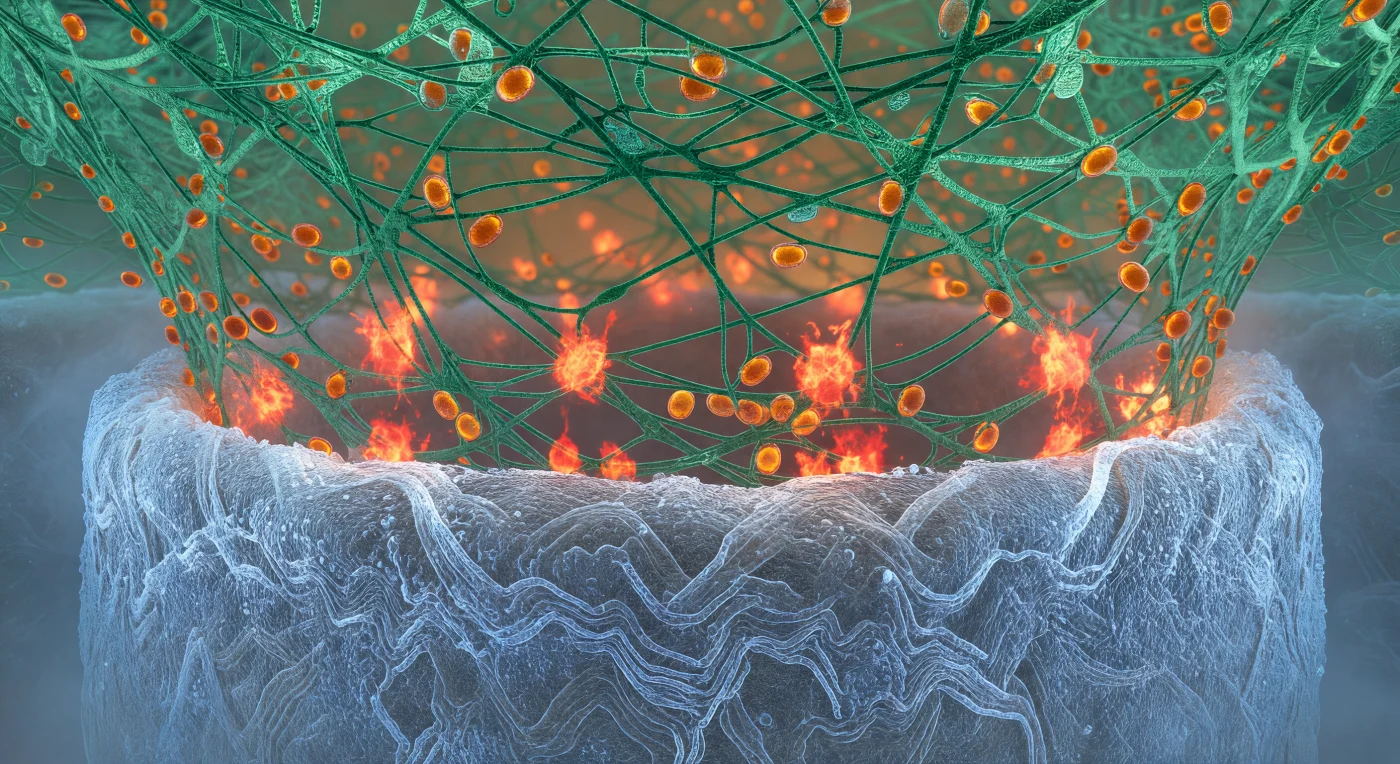
Je staat gevangen tussen twee biologische architecturen van onvoorstelbare complexiteit — onder je welft de celwand van de Candida-hyfe weg als de romp van een reusachtig schip, haar chitin-oppervlak stralend in koud blauwwit licht alsof elk polysacharidevezel van binnenuit gloeit, de textuur zo dicht en gelaagd als geperst zout of basalt dat door millennia van druk is gevormd. Van boven daalt de actinekorf van de neutrofiel neer als een gespannen koepel van vertakte F-actinefilamenten, elk draad scherp en harsachtig groen, de diepere lagen vervagende tot een zacht jadekleurig waas waar het netwerk te dicht wordt om te volgen. Op de contactzone — de dodingsgrens waarop je je bevindt — ontploffen onregelmatige uitbarstingen van roodoranje licht als trage chemische detonaties: geconcentreerde clusters van reactieve zuurstofverbindingen die de nabijgelegen actinedraden van onderen in koperkleurig vuur zetten, een smeulend verbrандingsfront dat drukt tussen chitinwand en immuunsteigers. Door het groene netwerk boven je drijven azurofiele granules als barnstenen druppels, ovale organellen geladen met enzymatisch materiaal, die zich traag maar gericht langs de actinekabels bewegen naar de heetste reactiepunten — verzegelde reservoirs die het front benaderen als munitie op weg naar de strijd.
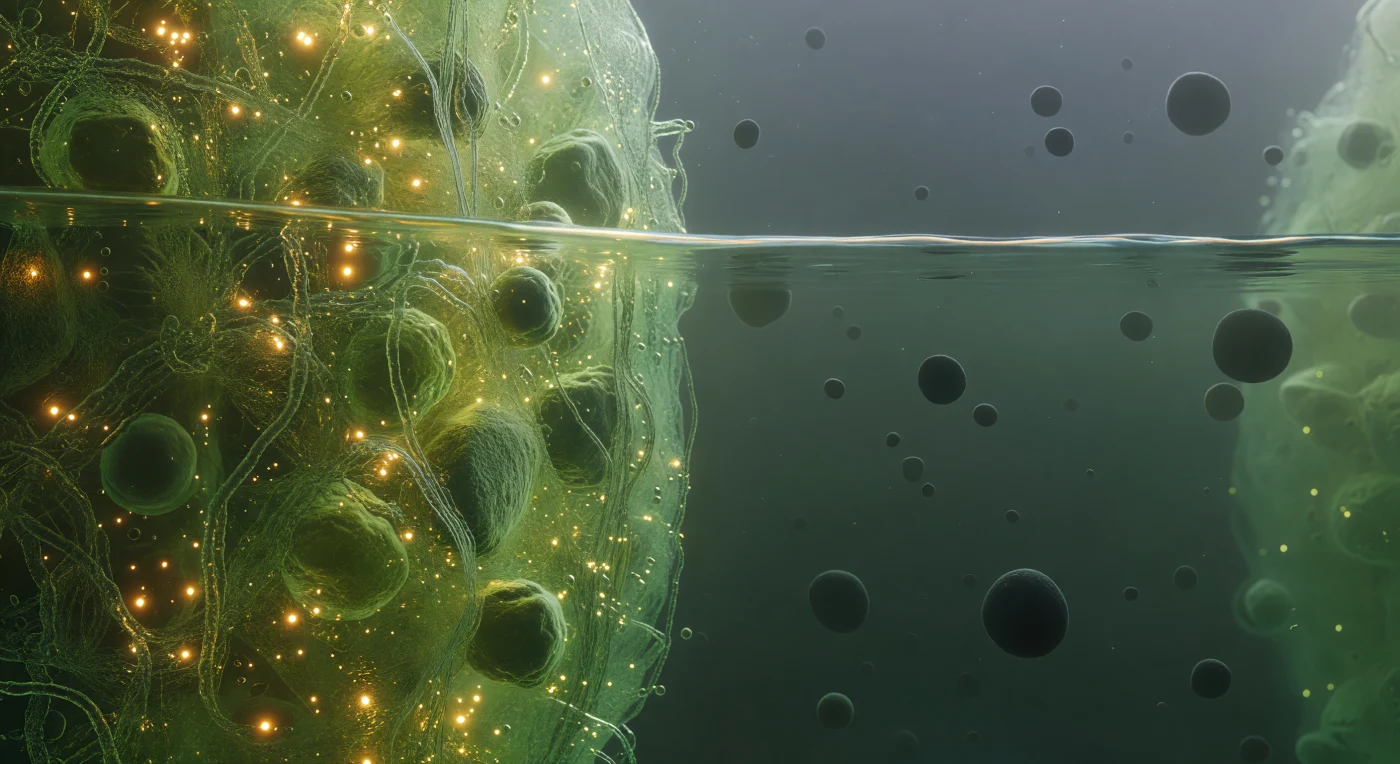
Je zweeft precies op de grens tussen twee werelden: aan je linkerhand zwelt het interieur van het stressgranule op als een diepgoudgroene zee, zo dicht opeengepakt met verstrengelede mRNA-ketens en intrinsiek ongeordende eiwitten dat het licht erin lijkt te vertragen en te poelen tot een warm barnsteen dat zachtjes pulseert van de Brownse beweging van tienduizenden macromoleculen. Dit is een biomoleculair condensaat, ontstaan door vloeistof-vloeistof fasescheiding — hetzelfde thermodynamische principe dat olie van water scheidt, maar hier uitgevoerd door eiwit- en RNA-sequenties met zwakke, meerwaardige bindingen die plotseling een dichte, vloeibare fase afzonderen van het omringende cytoplasma. Recht voor je bevindt zich de faseovergang zelf: geen geleidelijke overgang, maar een scherpe, licht trillende discontinuïteit die flauw iridesceert met capillaire fluctuaties op de nanoschaal, als matglas gezien van de zijkant, verlicht van binnenuit zodat een dunne lichtnaad de grens markeert waar moleculaire dichtheid abrupt instort. Rechts opent zich de ijle cytoplasma — een uitgestrekte, koelblauwe schemer waar afzonderlijke ribosomen als donkere, stille boulders door de nevel drijven, elk enorm in verhouding, gescheiden door afstanden die aanvoelen als open vlakte na de claustrofobische drukte van het condensaat.

Je zweeft in de nauwste ruimte die levend weefsel kent: de intercellulaire spleet tussen twee niertubulus-epitheelcellen, een vochtige corridor van nauwelijks twintig nanometer breed, gevuld met een goudachtig extracellulaire vloeistof die naar jou toe golft als een onderstroom van opgeloste ionen en glycocalyxsuikers. Boven je opent de ruimte zich plotseling in het tubulaire lumen — een zwarte, kathedraalse leegte, koud verlicht van onderen zoals de opening van een oceaantrog gezien vanuit de diepte. Wat je positie markeert is een ononderbroken gordel van stralend karmozijnrood: de strakke afsluiting van de tight junction, een gloeiende naad van claudines en ZO-1-eiwitten die de volledige celomtrek omcirkelt als een band van gesmolten metaal ingelast in levend membraan, de moleculaire sleutel die het lumen scheidt van het bloed en zo de selectieve reabsorptie van water, glucose en ionen door de nier mogelijk maakt. Vlak daaronder volgt een tweede ring van zacht smaragdgroen: de E-cadherine adherens junction, wier ectodomeinen als ineengestrengelde vingers over de spleet reiken en de mechanische cohesie tussen de cellen bewerkstelligen, terwijl dieper in de cellichamen enorme kobaltblauwe kernen hun chromatine als indigolicht door de kernmembraan uitstralen. De geometrie van het geheel — gloeiende loodlijnen, gekleurde glazen panelen van cytoplasma, de absolute duisternis van het lumen erboven — doet denken aan een glas-in-loodraam van binnenuit bezien, een levende architectuur die continu wordt onderhouden, gerepareerd en bewaakt door eiwitten die nooit slapen.
