Confiança científica: Alto
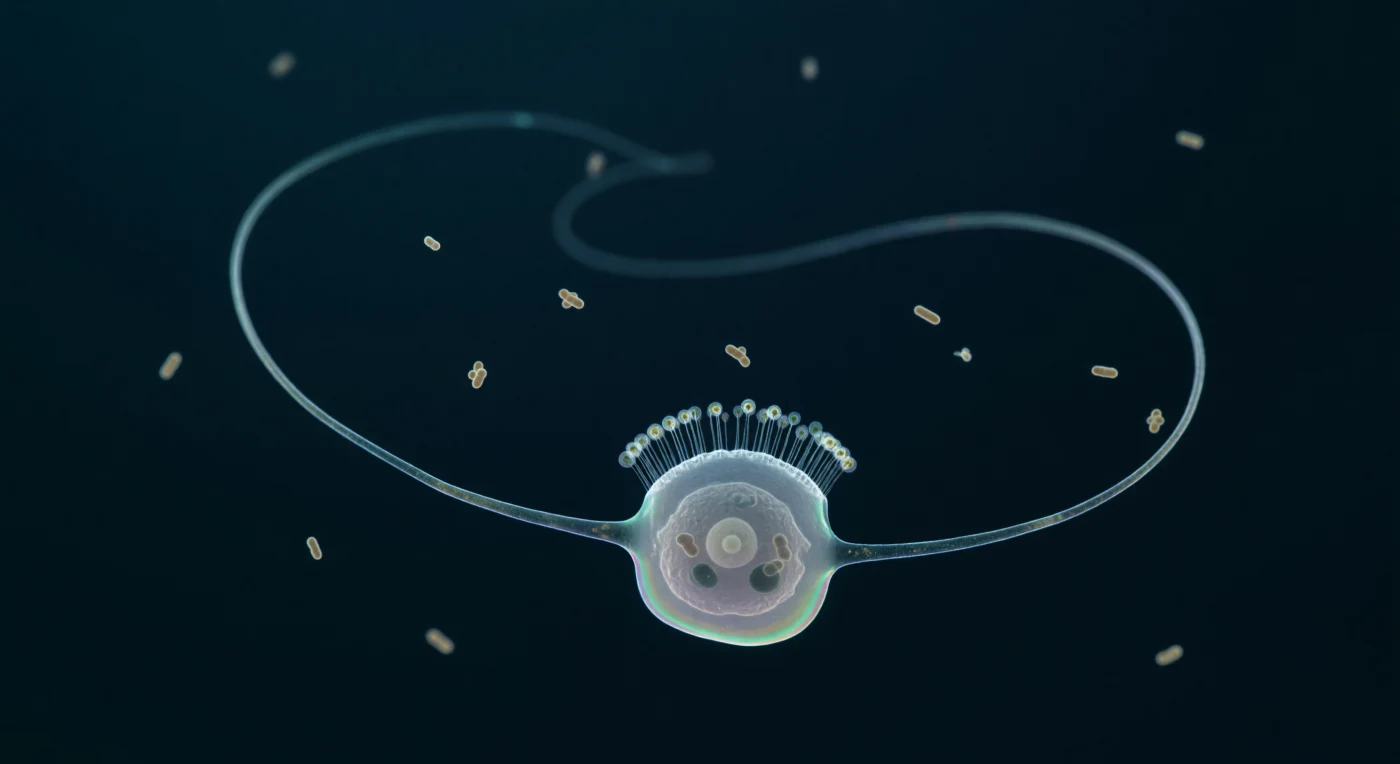
Você está suspenso numa escuridão pelágica absoluta, num meio tão viscoso a esta escala que pressiona contra si com a resistência silenciosa da glicerina fria — não água, não ar, mas um terceiro estado da matéria onde o movimento Browniano é o ruído de fundo permanente da existência. Dominando o seu campo de visão, um coanoflagelado único exibe o seu flagelo sinuoso em arco helical congelado, uma estrutura de vinte micrómetros que não é mais espessa que um fio de seda de aranha, coberta de glicocalice que dispersa a luz azul-verde descendente em iridescência oleosa, enquanto um rastro fantasma prateado — memória estrutural do batimento sinusoidal de uma fração de segundo atrás — ecoa no meio escuro como caligrafia apagada. A arquitectura celular revela-se através da membrana quase invisível: um núcleo vesicular pálido ocupa um terço do citoplasma translúcido, dois vacúolos alimentares densos guardam em seu interior a silhueta comprimida de bactérias a meio da digestão, e a colar de trinta e cinco microvilosidades — cada uma mais fina que o comprimento de onda da luz visível — refracta a iluminação apical numa coroa de halos de interferência em ouro frio e aguamarina. Em redor, uma dúzia de bactérias em forma de bastonete deriva em profundidades variadas, tremendo no lugar pelo jitter térmico do meio, algumas nítidas em âmbar quente, outras dissolvendo-se em bokeh suave na escuridão azul-negra que engole tudo a apenas alguns comprimentos de célula de distância.

Você está suspenso na água escura como um grão de bactéria, e diante de você domina o campo visual inteiro uma esfera prateada e eriçada que, à sua escala, tem a imponência de um edifício — a colônia roseta de *Salpingoeca rosetta*, com trinta e dois células dispostas em geometria radial perfeita, cada uma apontando para fora com o seu flagelo estendido numa coroa de chicotes quase invisíveis que conferem ao conjunto a silhueta elétrica de um ouriço-do-mar congelado no instante. Cada célula é um ovóide levemente translúcido de três a cinco micrômetros, com a sua sombra nuclear visível como uma elipsóide densa no interior, e na posição das duas horas uma célula em divisão exibe o corpo levemente estrangulado em forma de amendoim, os dois núcleos-filhos deslocados dentro do citoplasma alongado. As células convergem nos seus pólos basais por finos filamentos citoplasmáticos, tão delgados que aparecem apenas como linhas ligeiramente mais claras traçando raios até um núcleo central de matriz extracelular de tom âmbar-mel, a única cor quente num campo quase inteiramente acromático de prata e azul-negro. A água em redor não é transparente no sentido humano — é um meio opalescente de crepúsculo profundo, animado por um tremor browniano contínuo e por flocos de neve marinha que derivam ao fundo como fragmentos de matéria orgânica e polissacarídeos, estabelecendo a profundidade espacial e lembrando que esta colônia flutua livremente num oceano vivo e particulado, numa fronteira evolutiva de 600 milhões de anos entre a unicelularidade e os primeiros tecidos animais.
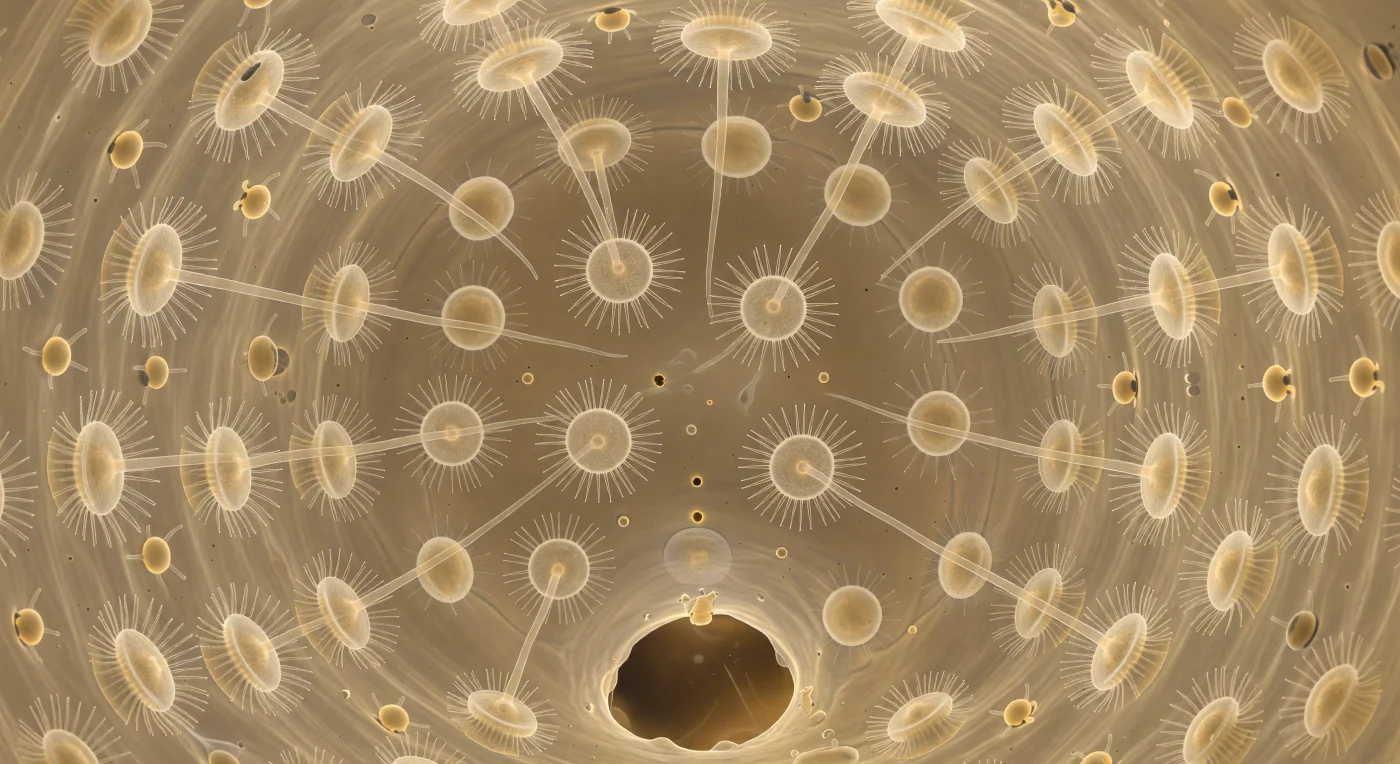
Você está suspenso no centro exato de uma câmara esférica com cerca de quarenta micrómetros de diâmetro, um espaço vivo cuja parede inteira é revestida por coanócitos alinhados ombro a ombro, cada célula coroada por um colar de microvilosidades translúcidas e um único flagelo que se projeta para o interior como um fio de vidro animado, todos batendo em fase desfasada para gerar uma ondulação contínua e orgânica que impele a água suavemente em direção ao poro apópilo aberto à sua frente. Dois poros prosópilo, visíveis como pequenas aberturas escuras na parede âmbar da câmara, admitem fios delicados de corrente carregados de bactérias e matéria orgânica dissolvida, alimentando este sistema de filtração que, multiplicado por milhões de câmaras idênticas, permite à esponja processar um volume de água equivalente a vinte mil vezes o seu próprio corpo a cada dia. Para além da camada de células, o mesohilo revela-se como uma malha fibrosa de ouro âmbar translúcido, percorrida por arqueócitos ameboides que se deslocam com a lentidão inevitável do vidro fundido, transportando nutrientes capturados para o interior do tecido. A luz que enche este espaço não tem origem pontual — é luz oceânica filtrada por várias camadas de membrana celular e matriz extracelular, chegando até si como um brilho quente e difuso que transforma a câmara num lampião vivo, cuja geometria perfeita existe inteiramente ao serviço da captura do invisível.
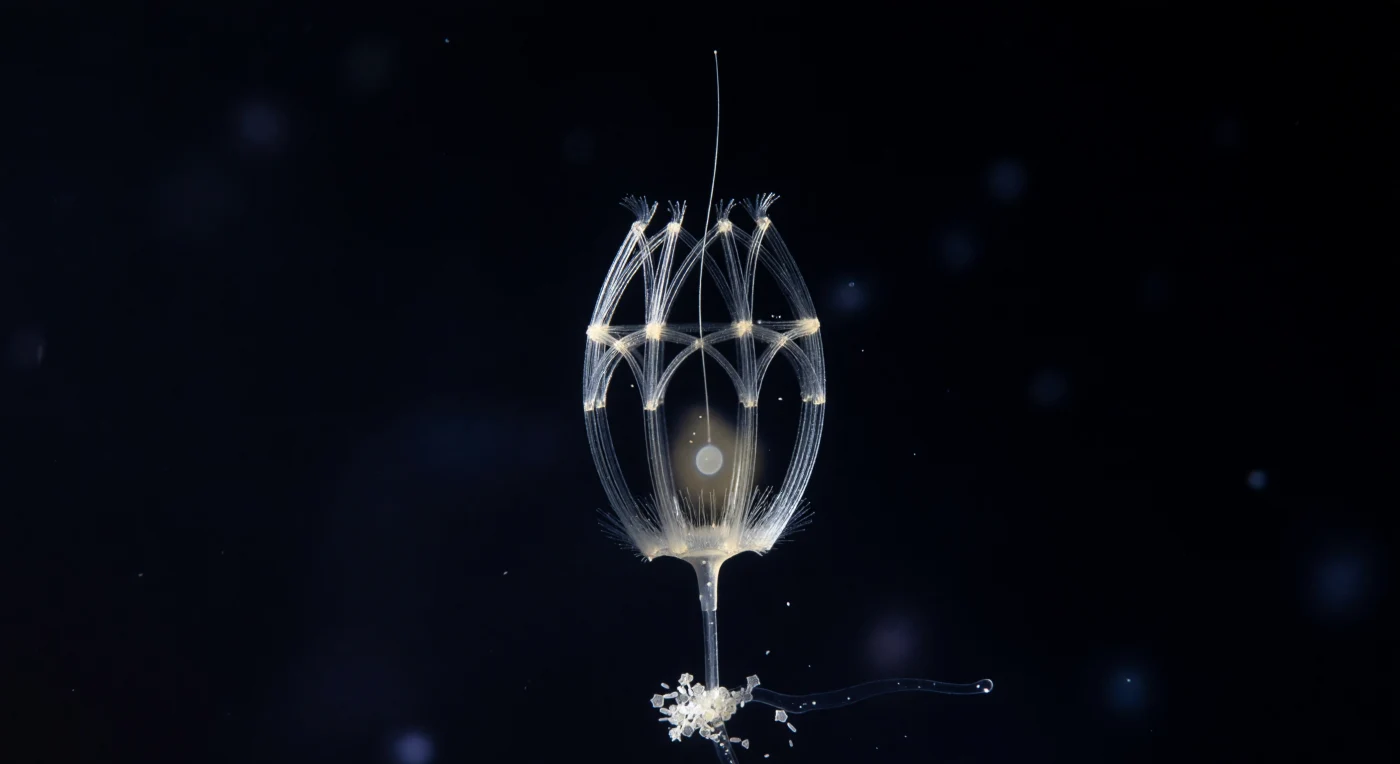
Suspensa numa coluna de água mesopelagial onde a escuridão tem a densidade de algo muito mais antigo do que a noite, a estrutura que ocupa o centro do campo visual impõe-se com a precisão perturbadora de um objeto arquitetónico: uma gaiola de sílica de vinte microns de diâmetro, montada a partir de dois sistemas de costal strips — costelas longitudinais que arqueiam da base ao ápice como as nervuras de uma abóbada gótica, apertadas a intervalos por arcos transversais — e onde a fonte de luz lateral rasante da esquerda toca cada varinha de vidro mineralizado, a sílica inflama-se num branco-prata de filamento incandescente, enquanto o lado oposto de cada haste mergulha em sombra completa, conferindo à lorica de Diaphanoeca grandis uma volumetria esculpida em chiaroscuro que a faz parecer ao mesmo tempo catedral, lanterna e ruína. No interior deste andaime luminoso, a célula viva é pouco mais do que um rumor: uma névoa citoplasmática de âmbar quase diluído até à invisibilidade, com o invólucro nuclear a ensaiar a sua forma como uma elipse de pérola apenas suficientemente densa para se fazer sentir, enquanto o flagelo sobe através do polo anterior aberto e se dissolve na escuridão acima como um fio de prata translúcida estendido em direção a algo que o organismo não consegue ver. O pedúnculo rígido ancora a lorica a um fragmento de neve marinha no bordo inferior do enquadramento — um agregado frouxo de frústulas de diatomáceas cujos painéis hexagonais de sílica funcionam como pequenos espelhos geométricos no feixe de luz, com bastonetes bacterianos dispersos entre eles, cada um uma fração do diâmetro da haste mais fina da gaiola, tudo envolvido numa membrana de muco transparente que capta a luz como uma película iridescente. A água circundante não é simplesmente negra: carrega a mais leve luminescência azul-violeta de bactérias bioluminescentes no limite extremo da visibilidade, e essa textura difusa da escuridão amplifica o isolamento da estrutura central, fazendo com que as hastes de sílica frágeis como vidro soprado pareçam permanentes como ferro e a célula no seu interior tão transitória quanto o vapor de respiração no frio.

Suspenso no exterior da parede cilíndrica de um *Euplectella aspergillum*, o olhar encontra uma catedral de vidro erguida nas trevas absolutas do fundo oceânico: uma grelha hexagonal de espículas de sílica fundidas em cada intersecção, tão regular na sua geometria que parece desenhada por um arquiteto em vez de secretada por um ser vivo. Cada espícula conduz a luminescência azul-esverdeada da água circundante como uma verdadeira fibra ótica, emitindo luz a partir do interior do próprio bastão de sílica e projetando para o átrio central uma malha de sombras geométricas que se multiplica em anéis concêntricos de claridade e penumbra. Nas secções fraturadas dos nós mais próximos, anéis de laminação concêntrica traem o crescimento lento e paciente desta estrutura — vidro biogénico depositado camada a camada em torno de um molde orgânico, semi-transparente no núcleo âmbar e branco glacial na bainha exterior. No interior do átrio iluminado, dois camarões comensal aparecem como silhuetas cor-de-âmbar e rosa, as suas antenas translúcidas recortadas contra o brilho suave, criaturas seladas para toda a vida dentro desta câmara de luz, suspensas na imobilidade perfeita que a pressão silenciosa de toda uma coluna de água impõe a tudo o que existe aqui em baixo.
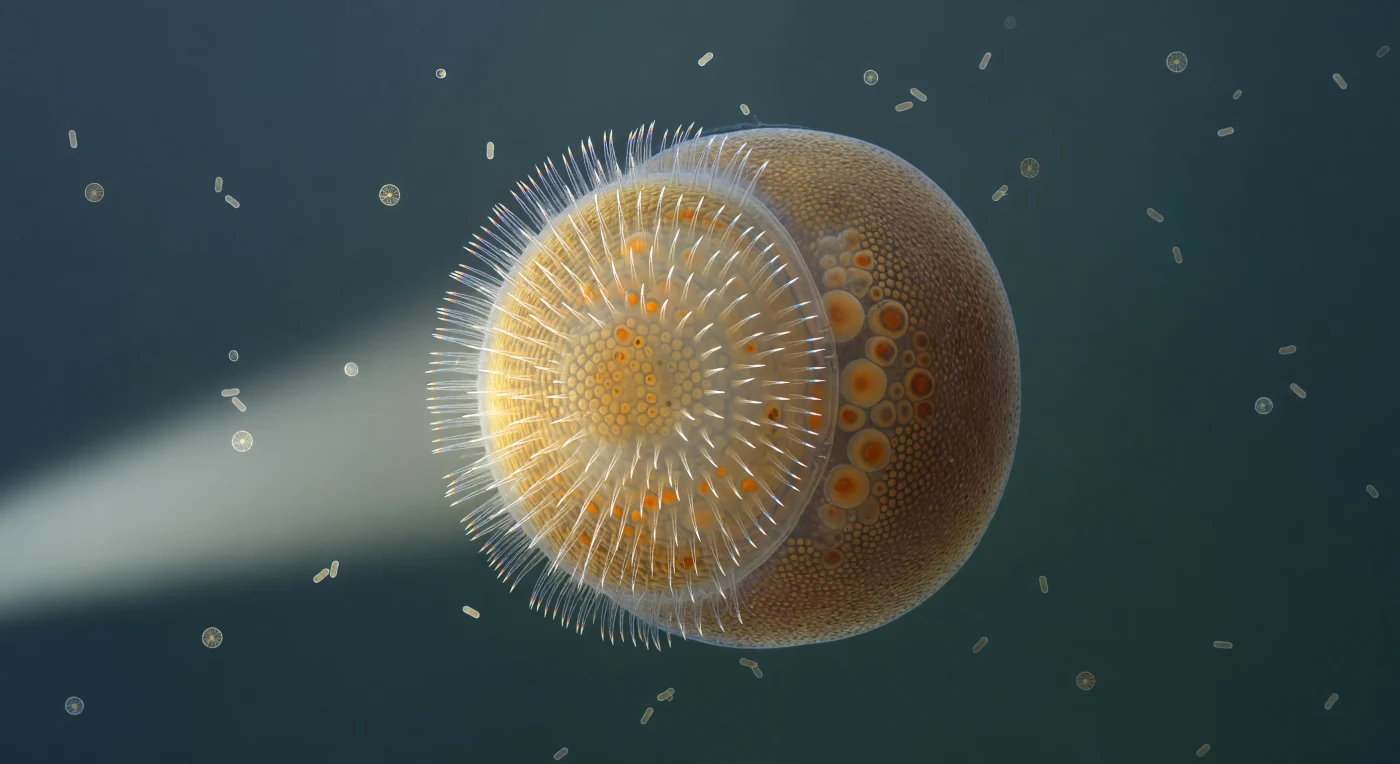
Você paira em plena coluna d'água como uma mota de poeira suspensa em gel luminoso, pressionado de todos os lados pelo frêmito invisível das moléculas dissolvidas, enquanto diante de você se ergue um mundo oval perfeito de cerca de quatrocentos micrómetros de diâmetro: a larva amphiblástula, uma forma de vida transitória e completa em si mesma, iluminada por um único feixe lateral de luz branca que a esculpe em crescente de ouro e sombra. O hemisfério anterior, denso de cílios batendo em ondas metacronais coordenadas, difrata a luz em uma coroa espectral frágil — violeta, ciano, âmbar pálido — que oscila e se reconstrói continuamente na borda equatorial como um prisma vivo, enquanto as membranas das células coanoblásticas subjacentes formam uma mosaico hexagonal quase imperceptível, translúcido e aquecido por dentro pelo brilho disperso dos corpúsculos lipídicos. O hemisfério posterior pertence a outro reino: maior, granuloso, carregado de arquéocitos repletos de vitelo âmbar-acastanhado cujas inclusões lipídicas se adivinham como pontuações escuras através da parede translúcida do corpo, toda a geometria interna revelada como um teatro de sombras suave. Esta larva — estrutura exclusiva das esponjas calcáreas, ponto de inflexão entre a dispersão e o assentamento, entre o ser pelágico e o ser séssil — carrega em si a lógica evolutiva dos primeiros metazoários: células ciliadas que propulsionam, células nutritivas que sustentam, e entre elas uma fronteira que é também uma história de mais de seiscentos milhões de anos.

O observador paira suspenso na água morna e salgada, diretamente sobre a borda de um imenso esponja barril, olhando para baixo em direção a uma abertura circular de quatro centímetros — o ósculum — cuja parede interna cor de terracota queimada desce em espiral para canais exalantes ramificados que mergulham na penumbra âmbar. Este orifício é a saída terminal de uma arquitetura de filtração viva: no interior do corpo da esponja, dezenas de milhares de coanócitos — células com flagelos batendo a 30–60 Hz — geram correntes coordenadas que atravessam câmaras esféricas de 25 a 50 micrômetros de diâmetro, filtrando bactérias e matéria orgânica dissolvida antes de expelir a água processada para cima, em jato contínuo, a uma taxa que pode atingir 20 000 vezes o volume corporal da esponja por dia. O jato exalante torna-se visível como uma coluna de distorção termodinâmica e turbulência interfacial, salpicada de partículas translúcidas — esferas âmbar de detrito orgânico, nuvens pálidas de bactérias, fragmentos de material silicoso — todas carregadas para cima pela corrente e dispersas em espirais na água aberta, captando a luz caustica azul-turquesa que desce da superfície. Ao redor do ósculum, a superfície rugosa da esponja estende-se em todas as direções, pontilhada de óstios escuros e colonizada por algas coralinas lavanda e rendas brancas de briozoários, enquanto corais ramificados fora de foco e peixes recifais cobalt-e-amarelo ocupam o fundo luminoso, lembrando que esta estrutura respiratória e microscópica existe imersa num dos ecossistemas mais complexos e iluminados do planeta.
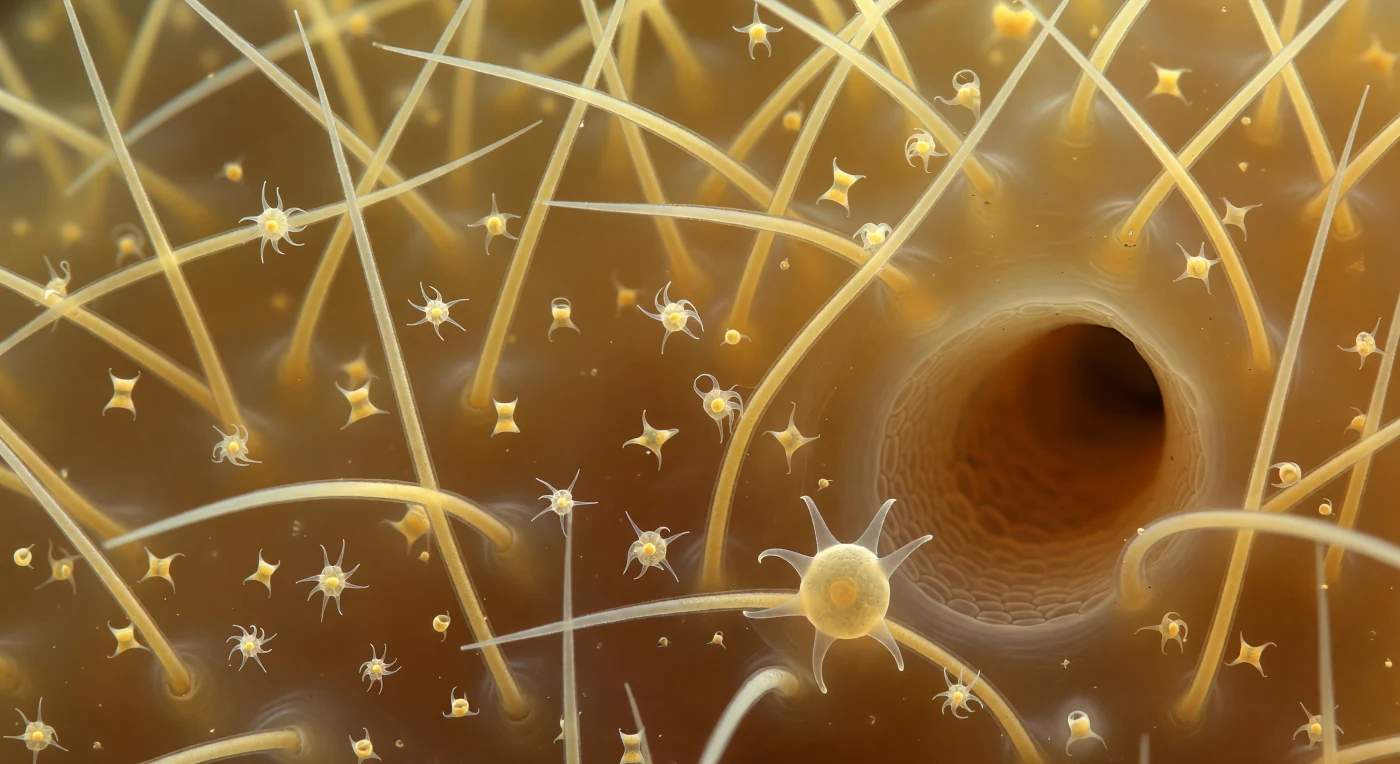
Dentro do mesohyl de uma esponja demosponge, o observador deriva como uma presença fantasma num cosmos âmbar e translúcido, rodeado por megaescleras oxea — agulhas de sílica amorfa com cerca de 200 micrómetros de comprimento, apontadas nas duas extremidades e percorridas por reflexos especulares brilhantes que traem a sua natureza essencialmente vítrea. Este esqueleto mineral não é inerte: as escleras são secretadas individualmente por células esclerócitas especializadas e mantidas em posição por fibras de espongina, cabos orgânicos cor de mel que correm em catenárias entre as bases das agulhas, conferindo ao conjunto uma rigidez tensional semelhante à de um andaime têxtil. Flutuando no gel coloidal entre as grandes escleras, microscleras sigma e quelas em espiral — peças com apenas vinte micrómetros — giram lentamente sob agitação Browniana, a sua geometria interna visível através da transparência do vidro biogénico, enquanto um túnel de canal escuro atravessa o plano médio como a entrada de uma caverna, delimitado pelo pinacoderma liso e as suas células poligonais tesseladas. Numa das escleras de primeiro plano, um arqueócito — célula totipotente responsável pelo transporte de nutrientes e pela diferenciação celular — agarra a superfície vítrea com pseudópodos estendidos em lâminas finas como película de sabão, o núcleo denso visível como uma esfera âmbar no interior do citoplasma translúcido: uma única célula animal sustentando, com química e toque deliberado, toda a arquitectura de um organismo que já existia quando os primeiros animais complexos ainda eram uma possibilidade evolutiva remota.
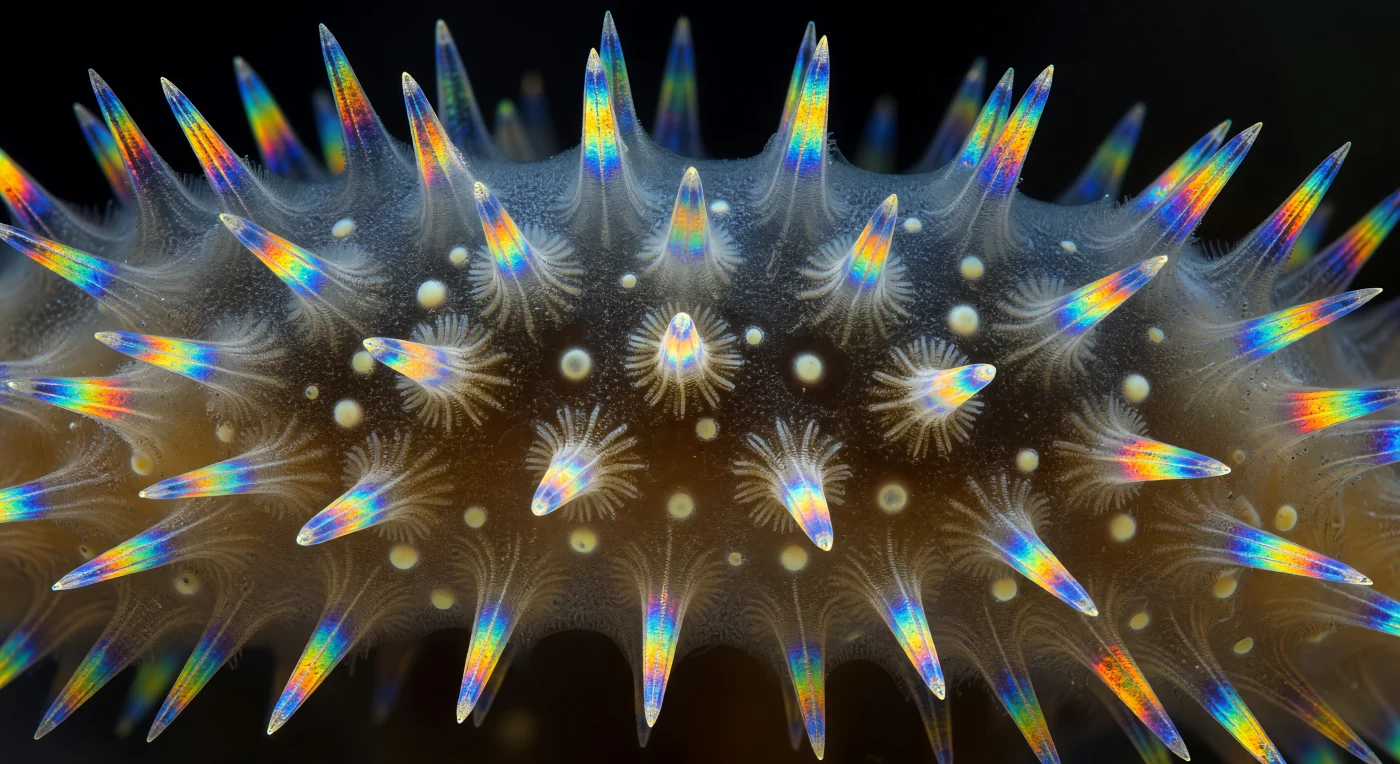
Dentro da parede seccionada de um *Sycon ciliatum* — uma esponja calcária pouco maior que uma unha —, o mundo revela-se como uma catedral de geometria viva: canais radiais abrem-se em corredores cilíndricos forrados por camadas de coanócitos, cujos cílios e colares de microvilosidades filtram a água que percorre o sistema aquífero do animal. A arquitectura repete-se em simetria radial perfeita, cada canal separado do seguinte por nervuras de mesoílo — um gel âmbar-escuro de matriz extracelular fibrosa, pontuado pelos núcleos pálidos de arqueócitos dispersos como lanternas na penumbra. O que verdadeiramente domina o campo visual são as espículas de calcite, triactinas e tetractinas, que irrompem pelo mesoílo em fogo birefringente: sob luz polarizada, cada cristal capta e decompõe a luz em azul cobalto elétrico, amarelo enxofre, laranja fundido e verde esmeralda intenso, enquanto o terceiro raio de cada triactina se projeta perpendicularmente para fora da parede como uma agulha apontada ao observador. Onde as espículas se cruzam, os padrões de interferência óptica acumulam-se em auréolas iridescentes que sangram de âmbar para violeta e azul-petróleo, evocando um vitral iluminado por um sol de inverno. A cena inteira — parte mineral, parte tecido vivo, parte puro efeito óptico — é a filtração do oceano reduzida à sua geometria essencial, suspensa num instante de luz espectral contra um fundo de negro absoluto.

Numa escuridão absoluta e gelada a duzentos metros de profundidade, as torres de *Aphrocallistes vastus* erguem-se do sedimento calcário como agulhas de uma catedral submersa, os seus esqueletos de espículas de sílica fundidas formando uma rendilhado hexagonal tão preciso que parece ter sido engenhado antes da existência dos animais terrestres — e, de facto, os hexactinelídeos são dos metazoários mais antigos, arquitectos do fundo oceânico há mais de quinhentos milhões de anos. O feixe do ROV atinge a coluna mais próxima de baixo, e a sílica conduz a luz ao longo de cada travessa como fibra óptica, fazendo a torre irradiar de dentro para fora numa luminescência creme-âmbar que ilumina câmaras internas aninhadas em sombras dentro de sombras, enquanto o sedimento ao redor faísca com fragmentos de espículas partidas, um tapete de agulhas de vidro microscópico. Estrelas-serpente drapeiam os seus braços bandeados pelas malhas da estrutura e um peixe-rocha paira imóvel no corredor de escuridão entre as torres, o olho captando a luz como um único ponto brilhante antes de se dissolver na sombra. Na periferia do feixe, onde a luz se extingue em azul-negro profundo, um rasto de bioluminescência azul-verde atravessa o campo de visão em três segundos e desaparece, lembrando que este recife se estende por centenas de metros em todas as direcções — uma floresta de paciência geológica e silêncio com textura própria.

Suspenso na camada bentónica de fronteira, o campo de visão inteiro é consumido por um platô alienígena de algas coralinas incrustantes — uma tundra magenta-carmesim calcificada, partida em placas poligonais irregulares com bordas levantadas como cristas tectónicas, salpicada de frústulas de diatomáceas que se erguem como monumentos de sílica vitrificada, captando a luz azul-esverdeada que desce filtrada pela coluna de água e projetando micro-sombras suaves sobre o biofilme bacteriano âmbar-dourado que preenche as depressões com géis de exopolissacáridos iridescentes. No centro desta paisagem, uma larva de esponja calcária recém-fixada — um disco creme-âmbar de cerca de 300 micrómetros de diâmetro — encontra-se no momento exato da metamorfose: a sua hemisfério ciliar colapsa para dentro numa inversão morfogenética irreversível, enquanto os futuros pinacocitos da margem periférica já se achatam em mosaicos pavimentosos, espalhando-se lateralmente a partir do ponto de fixação central como frentes de cristalização lentas. A larva capta a luz transmitida pela sua margem translúcida como uma membrana de vitral âmbar, revelando o neoglicocalix nascente em interferências de verde-azulado e ouro pálido. Esta é a fundação de um animal — uma célula única de decisão ecológica que, em poucas horas, reorganizará completamente a sua geometria celular para dar início a uma vida séssil de filtração contínua, herdeira de uma linhagem que partilha com os coanoflagelados as bases moleculares da adesão celular e da sinalização multicelular animal.
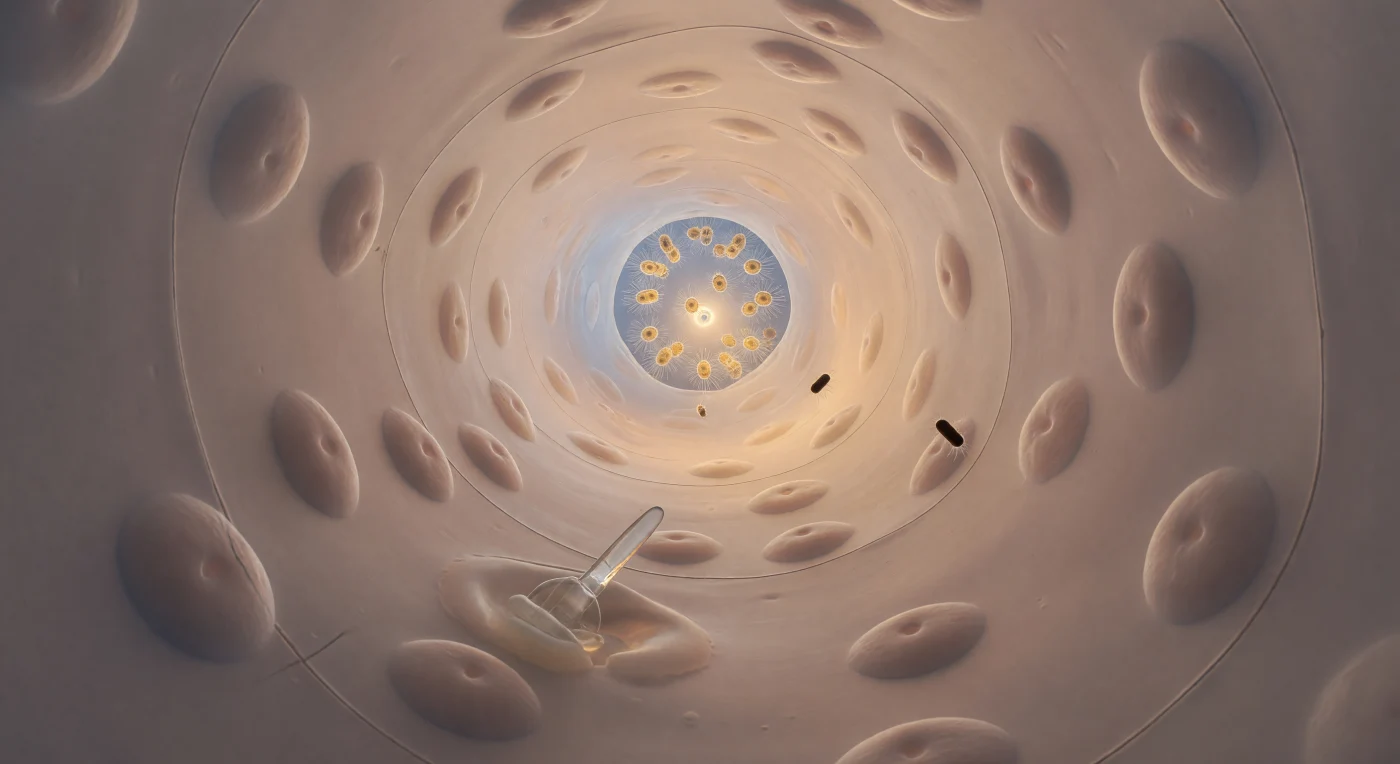
Você está suspenso na entrada de um túnel vivo com cerca de 70 micrómetros de diâmetro — não mais largo do que uma fração de cabelo humano — olhando ao longo de uma passagem cilíndrica e levemente curva revestida por endopinacocitos achatados como seda molhada, cujas membranas translúcidas de marfim e pêssego captam a luz como nácar polido, cada núcleo formando um suave montículo geológico sob a superfície quase vítrea. Na parede inferior, uma espícula de carbonato de cálcio atravessa o pinacoderme a baixo ângulo, dispersando um leve brilho prismático sobre as células vizinhas, enquanto um arqueócito ameboide empurra o seu citoplasma como cera morna entre a junção celular e a base da espícula, deformando-se e recuperando a forma numa lenta dança de remodelação tecidular. Atrás de si, luz oceânica azul-branca inunda a entrada do canal, iluminando a corrente laminar de regime de Stokes que o transporta suavemente para a frente — uma corrente tão viscosa a esta escala que duas bactérias em forma de bastão, com o brilho de cilindros de mogno polido, derivam sem turbulência ao seu lado, uma terceira a tombar lentamente sob o impulso do movimento browniano. No término do canal, a prósopila é um portal luminoso de apenas cinco micrómetros de largura, emoldurado pelas margens apressadas de dois pinacocitos, através do qual irradia o âmbar quente da câmara de coanócitos — dezenas de células com colarinho batendo os seus flagelos em uníssono, filtrando o oceano com uma paciência acumulada ao longo de seiscentos milhões de anos.
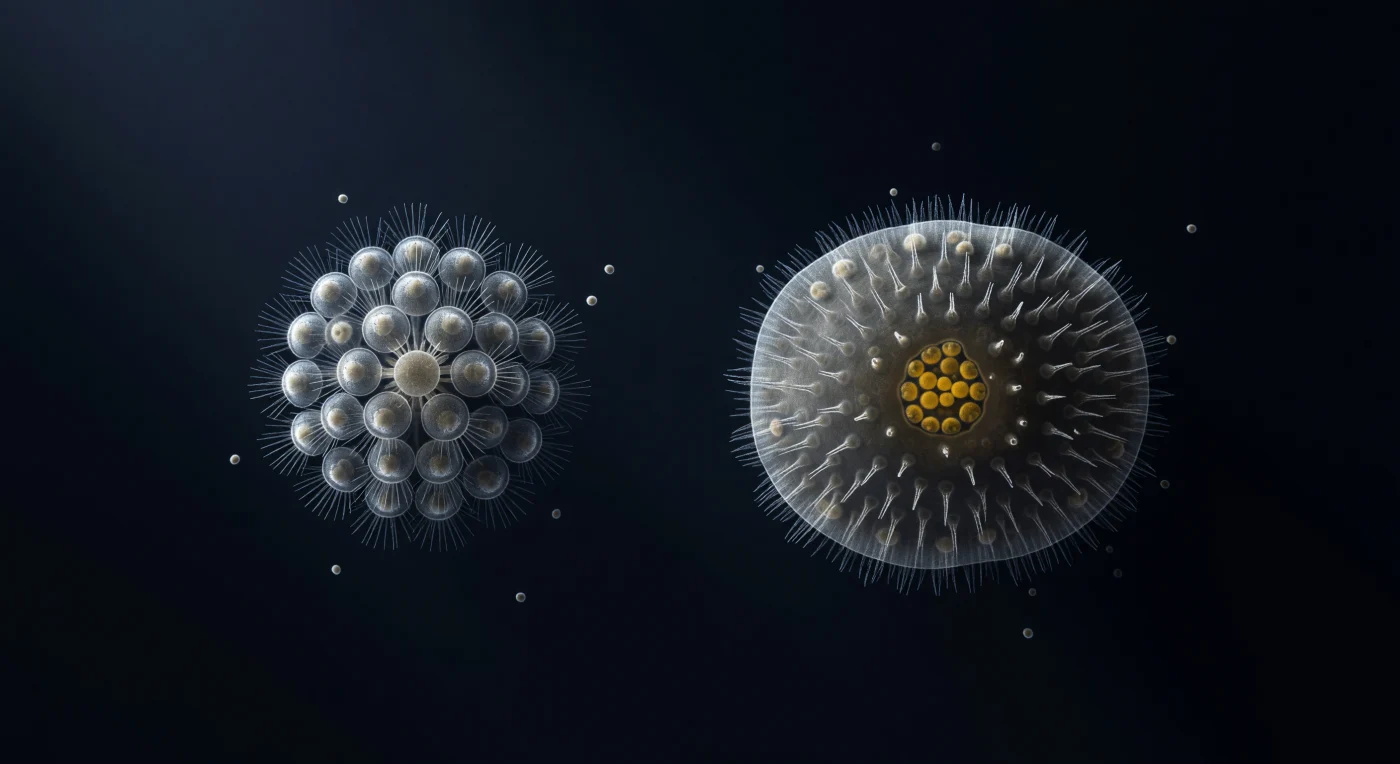
O campo de visão que se abre diante de ti é um vazio de azul-negro profundo, iluminado em contraste interferencial diferencial por uma luz oblíqua que raspa desde o canto superior esquerdo, esculpindo cada membrana celular em alto-relevo de prata e sombra — e dentro deste silêncio pressurizado, dois organismos separam-nos de nós mesmos por setecentos milhões de anos de história evolutiva. À esquerda, a roseta colonial de *Salpingoeca rosetta* ergue-se com precisão quase cristalina: trinta e duas células dispostas em esfera perfeita, cada uma apresentando para o exterior a sua unidade colar-e-flagelo, os microvilos do colar apanhados pela luz oblíqua como agulhas de vidro fino, a arquitectura molecular de pontes intercelulares na base da colónia sendo o esboço mais antigo conhecido de uma corporalidade partilhada. À direita, a larva parênquimela de uma esponja calcárea ocupa o campo com uma densidade orgânica completamente diferente: a sua superfície exterior mantida por uma camada contínua de células ciliadas cujos cílios pousam imóveis como pêlo em fotografia, o interior celular denso e semi-opaco, e nas células posteriores carregadas de vitelo aquece a única cor quente de toda a cena — um âmbar profundo e vivo, reservas lipídicas a arder como brasa num mundo de prata e gelo. O intervalo de cem microns de água aparentemente vazia que separa os dois organismos não contém quase nada — apenas algumas silhuetas bacterianas à deriva — mas carrega o peso implícito da transição mais decisiva da história animal: o momento em que uma célula com colar aprendeu a partilhar o corpo com outras, e o conjunto tornou-se mais do que a soma das suas partes.

Você existe à escala de uma bactéria, suspenso no interior da parede bissetada de uma gêmula de *Spongilla*, olhando para dentro de uma câmara seccionada que se abre diante de você como a nave de uma catedral partida por uma falha geológica. À sua esquerda ergue-se a muralha tripartida: uma membrana externa âmbar e levemente iridescente, seguida pela paliçada de espículas anfidisco — minúsculos halteres de sílica cristalina dispostos em fila cerrada no cimento de espongina cor de creme, os seus discos rotulares divergindo como rondelas de vidro fosco e lançando reflexos prismáticos em branco gelo, azul espectral e rosa pálido — e, mais internamente, uma camada lisa de espongina dourada, quase lacada, que delimita o espaço interior com cerca de sessenta micrómetros de espessura total. Para além dessa fronteira, o interior da gêmula irradia um calor âmbar e alaranjado: os arqueócitos estão embalados ombro a ombro, cada célula globosa inchada de gotículas lipídicas em açafrão, tangerina e amarelo cádmio, que brilham como lamparinas em suspensão animada, reservas metabolizáveis seladas quimicamente contra o inverno. À direita, o tubo do micropilo perfura a armadura em secção transversal — uma passagem cilíndrica actualmente bloqueada por uma coluna compacta de células pálidas encostadas umas às outras como rolhas numa garrafa, a sua abertura circular fitando o exterior como uma escotilha voltada para águas frias e escuras, enquanto o contraste entre esse mundo exterior morno e cinzento-oliváceo e o fulgor interior incandescente confere à cena o peso visual de um geodo partido em pleno inverno.

Você paira a centímetros acima de uma madeira submersa e encharcada, o seu campo de visão inteiramente dominado por uma crosta viva que se estende como uma tundra de veludo verde-elétrico — a esponja de água doce *Spongilla lacustris*, cujo tom vibrante nasce de uma aliança antiga com algas fotossintéticas que habitam o seu tecido vivo, convertendo cada feixe de luz que atravessa a superfície ondulada do ribeiro numa incandescência quase interior. A textura da crosta é enganosa na sua aparente macieza: sob o veludo verde escondem-se espículas de sílica que se projectam como agulhas de vidro em cada milímetro da superfície, apanhando a luz nas suas pontas como cristais de geada, enquanto a transição do verde luminoso para o creme amarelado nas zonas de sombra traça uma topografia fotossintética em gradientes de milímetros. Encravadas na crosta, esferas de mogno escuro afloram como paralelepípedos sepultados — os gêmulos, cápsulas de sobrevivência de meio milímetro de diâmetro ornamentadas com uma arquitectura radial de espículas, pacientes e inertes entre as células vivas que bombeiam incessantemente água a um ritmo de vinte mil vezes o volume do corpo por dia. Ostracodes rolam pela superfície como pérolas de semente sobre veludo, e planárias translúcidas deslizam em arcos lentos, os seus corpos deixando entrever o verde da esponja através do ventre, tudo isto sob uma catedral de água fresca e clara onde colunas de luz volumétrica descem da superfície prateada e varrem a crosta em ondas suaves de claridade e sombra. Este organismo filtrante, que não alterou o seu plano fundamental em seiscentos milhões de anos, continua a desempenhar a mesma função ancestral que tornou possível a transição para a multicelularidade animal — um arquivo vivo da biologia que nos originou.

O que se vê é uma planície alaranjada e brilhante que se estende em todas as direções como um lamaçal alienígena fotografado por um drone rasante, a superfície externa de uma esponja demosponge tropical revelando-se como um mosaico de células poligonais achatadas, cujas bordas elevadas capturam a luz filtrada do recife e projetam sombras finas nos sulcos entre elas, o pigmento carotenóide tão concentrado que parece irradiar calor de dentro para fora como terracota aquecida. Distribuídos por essa paisagem em intervalos irregulares, os óstios perfuram a superfície como caldeiras vulcânicas — alguns completamente dilatados, revelando a escuridão absoluta dos canais internos que conduzem a água pelo sistema de filtragem da esponja, outros contraídos em fendas quase fechadas pelo músculo circular do porocito, com linhas de tensão radiais no revestimento de mucopolissacarídeos denunciando a sucção contínua do corrente inalante. Sobre toda essa extensão, uma comunidade microbiana tridimensional de bactérias bastonetes, cocos e cianobactérias filamentosas habita a fina camada de muco que reveste as células, formando uma penugem viva de apenas alguns micrómetros de espessura que confere à superfície uma textura levemente fosca. No primeiro plano, um pequeno poliqueta emerge de um dos óstios mais largos, seus palpos ciliados varrendo a camada limite em movimentos lentos e deliberados, enquanto no canto superior direito um copépode aterra brevemente — seu exoesqueleto transparente refratando a luz azul do recife como uma lente de vidro — antes de ser defletido pelo gradiente de pressão invisível gerado por um ósculo próximo, a negociação constante entre filtração, predação e vida microbiana inscrita em cada milímetro quadrado desta pele viva.
