Confiança científica: Alto
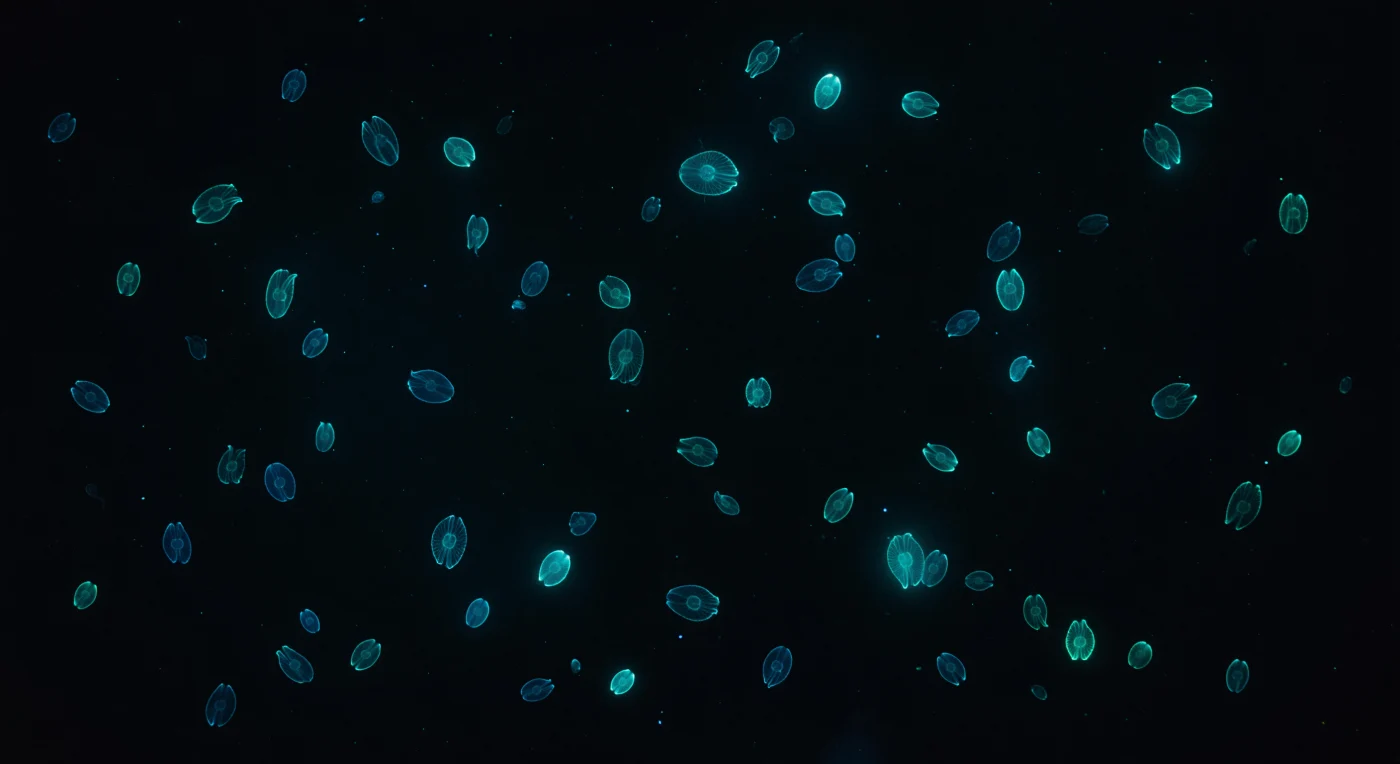
Suspenso em escuridão oceânica absoluta a dois metros abaixo de uma superfície sem lua, o olhar encontra dezenas de *Mnemiopsis leidyi* que derivam em todas as direções, os seus corpos de mesogleia quase completamente invisíveis — apenas oito fitas luminosas de fogo azul-esverdeado frio traçam o contorno oval de cada animal contra o vazio, emitindo precisamente a 490 nanómetros esse tom particular de azul-ciano que não existe em nenhum ambiente terrestre. Ondas de ativação bioluminescente percorrem continuamente cada fileira de ctenas do polo aboral em direção ao oral, um pulso lento e hipnótico como respiração feita de fotões, levando cerca de um segundo a traversar o comprimento do corpo antes de a fileira se apagar e reacender desde o topo — resultado da ação coordenada de fotócitos localizados na mesogleia, onde reações luciferina-luciferase convertem energia química em luz fria sem calor detetável. O animal mais próximo paira a talvez trinta centímetros, a sua geometria reduzida a uma catedral de arame luminoso, os lóbulos orais definidos apenas como espaço negativo onde a luminescência curva e termina, enquanto um ligeiro efeito de lente revela a menisco de refração entre a mesogleia e a água circundante. Ao fundo, quarenta ou cinquenta organismos adicionais dissolvem-se progressivamente em halos ciano difusos — cada fotão disperso pela neve marinha e pelo microzoo planctónico em suspensão, dando à coluna de água negra uma textura volumétrica que transforma este recife de gelatina em nebulosa viva.

Suspenso em escuridão absoluta a quatrocentos metros de profundidade, o observador não existe até que o feixe azul do ROV corte a coluna de água e o mundo se construa num instante — um cone de luz elétrica a 470 nanômetros revelando as formas de *Bathocyroe fosteri*, ctenóforos lobados cujos corpos de mesogleia pigmentada absorvem o azul circundante como tecido de veludo escuro, tornando-se voids negros que só traem a sua presença quando os lobos orais carmesim irrompem contra o nada com a saturação improvável de uma brasa. A mesogleia é um gel viscoelástico, noventa e seis por cento água por massa, com índice de refração calibrado ao da própria água do mar — matéria construída para ser invisível —, mas a pigmentação profunda de vinho tinto que protege estas formas mesopelagicas da bioluminescência dos predadores converte-se aqui numa assinatura inesperadamente luminosa sob iluminação artificial. Ao longo de cada fileira de cílios compostos, as placas ctenas fragmentam o feixe direcional em pulsos iridescentes — violeta, verde-azulado, âmbar fugazes — não bioluminescência, mas difração pura, redes de graticulado vivo refratingindo luz em sequência metacronal como sinais elétricos percorrendo um nervo. Ao redor de tudo isto, neve marinha desce com paciência glacial através do feixe iluminado, flocos de muco e colónias bacterianas entre um e cinco milímetros, fornecendo à cena a sua profundidade espacial enquanto, além da margem da luz, os animais não escurecem mas cessam de existir, dissolvendo-se no que não é cor alguma mas a negação de toda cor.

Suspensos na coluna d'água mediterrânica, encontramo-nos frente a frente com uma *Bolinopsis infundibulum* cujo corpo de mesogleia — quase indistinguível do mar que o envolve, com índice de refração tão próximo ao da água que apenas uma ténue distorção óptica trai os seus contornos — se revela sobretudo através dos oito fileiras de pentes que ardem em ondas metacrónicas antipleticas: carmim profundo desdobrando-se em âmbar, depois verde-ácido, depois violeta elétrico, cada tonalidade perseguindo a seguinte como aurora lenta sobre um espelho molhado. Estes pentes são redes de difração biológicas, formadas por placas de cílios compostos com algumas centenas de micrómetros de largura que, ao bater em sequência a trinta ciclos por segundo, desviam a luz solar em comprimentos de onda distintos consoante o ângulo de cada pancada — não bioluminescência, mas cor estrutural pura, efêmera e física. Os lobos orais do animal caem em pregas suaves, levemente rosados onde os canais gastrovasculares ramificados na mesogleia se tornam visíveis contra o brilho da superfície, como capilares vistos através de uma mão pressionada contra a luz. Lá em baixo, a luz mediterrânica fragmentada pela ondulação projeta rendas de cáusticas douradas sobre o sedimento ripado a alguns metros de distância, enquanto a coluna de água entre nós e o fundo glows em gradientes de turquesa e cobalto — um espaço de nave de catedral que torna esta criatura de vidro e arco-íris simultaneamente íntima na sua delicadeza e oceânica na sua estranheza.
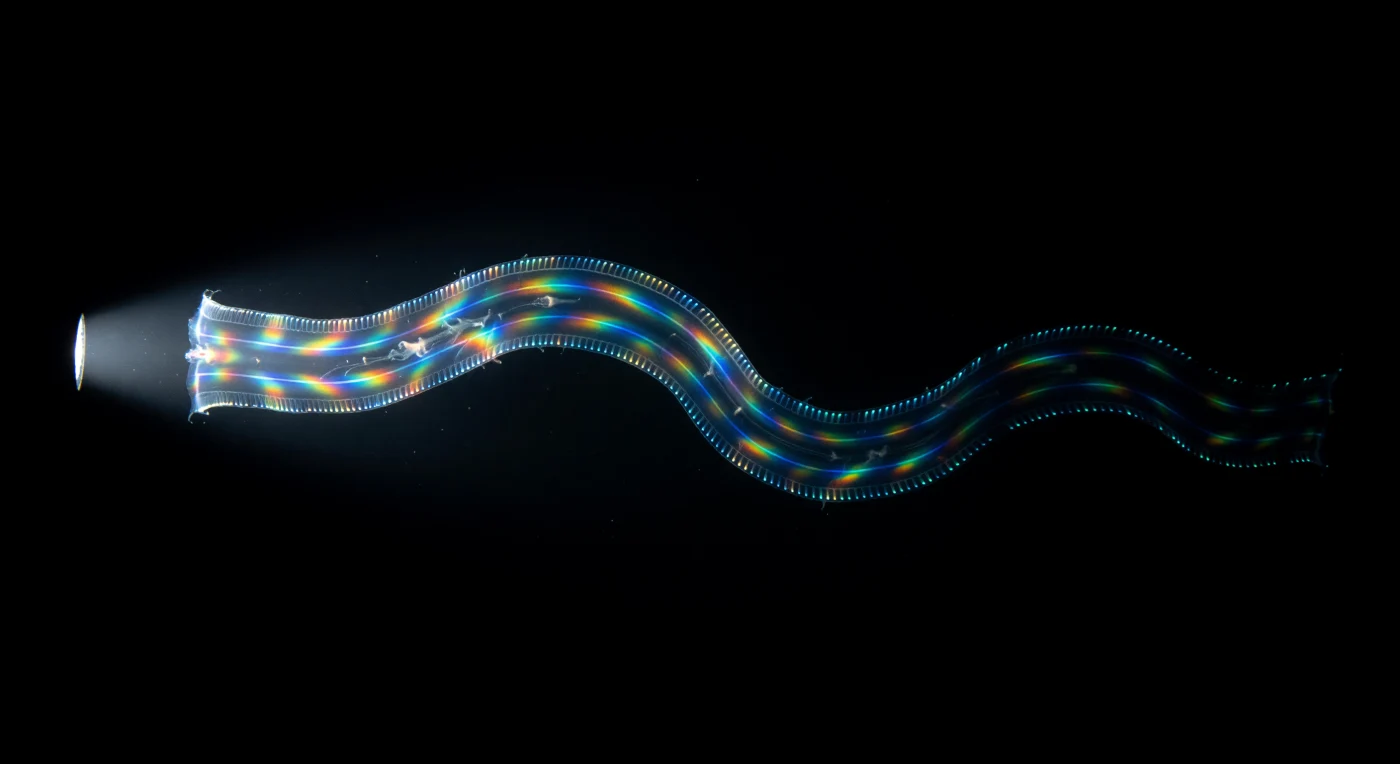
Suspenso a três metros abaixo da superfície do Atlântico tropical, o mergulhador contempla uma fita translúcida de sessenta centímetros que se materializa lentamente da escuridão absoluta — *Cestum veneris*, o cinto de Vénus, um ctenóforo tão opticamente próximo da água salgada que se revela apenas como uma ondulação da escuridão, um leve arqueamento do nada. As suas quatro fileiras de pentes correm ao longo de toda a extensão do corpo como costuras luminosas, cada placa ciliar batendo em ondas metacronais a 15 a 35 Hz, disparando fotócitos no mesogléia subjacente e traçando fios de bioluminescência azul-esverdeada a 490 nanómetros ao longo de cada margem da fita — quatro linhas de foxfire frio que revelam a arquitectura sinusoidal do animal como um diagrama de onda vivo. Quando o feixe da lanterna golpeia o corpo do animal em cheio, as fileiras de pentes transformam-se subitamente em redes de difracção em movimento, e a iridescência estrutural latente nas cílias em batimento explode em fogo espectral — vermelho a sangrar em laranja, em ouro, em cobalto, em violeta, a cascata cromática deslocando-se em onda de uma extremidade à outra como um estandarte de seda que se desenrola e volta a enrolar a cada ciclo. A fita ondula para fora do feixe e regressa à escuridão, os seus bordos bioluminescentes voltando a ser a única prova da sua existência, quatro linhas frias e flexíveis que se dissolvem progressivamente no negro impenetrável do oceano.

Suspenso a quinze metros de profundidade nas águas negras e turvas de um loch escocês em fevereiro, o observador encontra-se rodeado por uma deriva silenciosa de Pleurobrachia pileus — ctenóforos esféricos do tamanho de berlindes de vidro, cada um com apenas dois centímetros de diâmetro, cujos corpos de mesogleia quase inteiramente compostos por água se confundem com o próprio meio que habitam, traindo a sua presença apenas como uma ténue distorção refractiva no verde-cinzento da coluna de água. O interior de vários indivíduos revela um núcleo de avelã-e-salsa luminoso, o conteúdo digestivo visível através de paredes corporais praticamente invisíveis, enquanto atrás de cada esfera se estendem tentáculos gossâmeros de quinze centímetros cobertos de coloblastos — estruturas adesivas únicas deste filo — que desaparecem completamente na suspensão de diatomáceas fragmentadas e matéria orgânica particulada que reduz a visibilidade a escassos metros. Os oito cordões de pentes ciliares do indivíduo mais próximo percorrem a superfície da esfera como meridianos de um mundo minúsculo, cada placa composta por centenas de milhares de cílios fundidos que batem em ondas metacrónicas a dezenas de hertz, gerando a célebre iridescência estrutural que percorre as fileiras em rosas e âmbares apagados — não uma luz emitida, mas cor nascida do movimento, fugaz e quase respiratória. No limite inferior do enquadramento, a margem sombria de uma fronde de alga castanha interrompe este universo de gel e água fria, o único material denso e sólido numa cena construída quase inteiramente de nada.

Suspensos a dois metros abaixo de uma superfície que fragmenta a luz do final da tarde em redes douradas e inquietas, assistimos ao momento mais íntimo da predação pelágica: uma *Beroe cucumis* de rosa-salmão dilata a sua macrostome em oval improvável para engolir mais de metade de uma *Mnemiopsis leidyi* ainda viva, cujas filas de pentes posteriores continuam a bater em defesa mecânica, varrendo o sol oblíquo em cascatas de cor estrutural que deslizam do violeta ao âmbar ao verde em frações de segundo. O corpo da *Beroe* é mesogleia tão transparente que funciona como ar corado — os canais meridionais ramificam-se pelo seu interior como um delta fluvial visto do alto, tributários cor-de-rosa que pulsam suavemente enquanto a digestão começa, e onde os dois corpos se interpenetram o lábio da predadora estica-se fino como membrana, refratando o ouro ambiente numa costura iridescente e trémula. A *Mnemiopsis* que ainda sobressai da boca exibe a sua arquitetura fantasma — mesogleia quase incolor, faringe comprimida, canais gastrovasculares pálidos — enquanto os tentáculos filamentosos da extremidade oral se dissolvem no brilho rosado do interior da predadora. As cáusticas da superfície varrem continuamente ambos os corpos em padrões de interferência dourada, curvando-se à volta da mesogleia curva como se os organismos fossem lentes vivas, e as últimas placas posteriores da *Mnemiopsis* ainda ciclam os seus arco-íris para a água aberta — uma transmissão que ainda não soube que o sinal se perdeu.

Suspendos na coluna d'água negra de um estuário de verão, somos envoltos por três corpos de *Mnemiopsis leidyi* que se elevam como abóbadas de catedral em gel translúcido, as suas paredes de mesogleia tão afinadas em índice de refração à água circundante que os percebemos menos como formas sólidas e mais como distorções suaves no espaço, enquanto as suas oito fileiras de ctenas pulsam em ondas metacrónicas e incendeiam a água com cascatas de azul-cobalto a verde-água a 490 nm, um brilho frio que se difunde pelo interior gelatinoso e ilumina os canais gastrovasculares como rendas escuras dentro de luz diluída. Dos poros gonoporos ao longo dos canais meridionais, exalam-se lentamente plumas leitosas de esperma — não jatos, mas seepagens viscosas que captam a bioluminescência vizinha e se transformam em névoa pérola e prateada à medida que derivam com a microcorrente, transportando sinais químicos que orientam os gâmetas invisivelmente, e onde duas plumas se cruzam formam uma neblina reprodutiva suspensa na água pelágica como cirro a grande altitude. À deriva neste ambiente de urgência reprodutiva surgem os ovos — esferas refratoras de cerca de 120 micrómetros, que à nossa escala assumem o tamanho de pequenas esferas de vidro, as suas superfícies curvadas concentrando a bioluminescência circundante numa caustica interna brilhante rodeada por um halo difuso de cor espectral do violeta ao laranja — dezenas deles faíscam simultaneamente a diferentes profundidades, compondo uma constelação tridimensional de pontos frios que se dissolve gradualmente na escuridão absoluta a dez metros abaixo da superfície da baía às duas da madrugada.

Olhando para cima a partir de doze metros de profundidade nas águas frias do Hardangerfjord, centenas de *Bolinopsis infundibulum* suspendem-se em toda a coluna d'água como ornamentos de vidro num candelabro invisível — corpos ovais de oito a doze centímetros, compostos quase inteiramente de mesogle ia com índice de refração tão próximo ao da água do mar que se revelam apenas como distorções leves, lentes biológicas que traem a sua presença quando um feixe de luz nórdica de verão os atravessa em ângulo certeiro. Ao longo de cada corpo correm oito fileiras de ctenas meridionais, bandas de cílios fundidos que actuam como redes de difração vivas, decompondo a luz em vermelho, laranja, amarelo, verde e violeta numa sequência metacronal contínua e assíncrona entre indivíduos, de modo que o campo acima fervilha com um fogo policromático lento e rolante — um lustre de fibras ópticas animadas cujas listras espectrais nunca se alinham todas ao mesmo tempo. Vários dos organismos mais próximos exibem um rubor âmbar-alaranjado difuso nos seus canais gastrovasculares, o vestígio translúcido de copépodes recentemente ingeridos visível através da mesogleia incolor como uma gema suspensa em gelatina. A superfície prateada do fjord flutua longe acima como um tecto de espelho cravejado de redes causticas rastejantes, enquanto colunas volumétricas de luz azul-branca descem obliquamente através da névoa de fitoplâncton de fim de verão, intersectando o candelabro vivo e espalhando-se em leques prismáticos sobrepostos que enchem a água entre os corpos com uma bruma cromática difusa e silenciosa.

Diante de você, suspenso na escuridão absoluta a mil metros de profundidade no Canhão de Monterey, aparece algo que parece impossível: *Lampocteis cruentiventer*, uma ctenófora lobada da cor do sangue arterial, iluminada pelo cone azul dos LEDs do ROV como uma joia cirúrgica contra o nada. A mesogleia escarlate — um gel viscoelástico composto sobretudo de água e fibras de colagénio, cuja pigmentação absorve todos os comprimentos de onda abaixo dos 600 nm — devolve apenas o seu próprio carmim profundo, criando a ilusão de que o animal irradia luz própria num espectro que a câmara não consegue fornecer. Os amplos lobos orais pairam abertos e relaxados como pétalas de veludo, as suas margens superiores captando reflexos ciano nítidos que traçam a curvatura exata do tecido antes de se perderem na sombra cor de vinho, enquanto oito filas de pentes percorrem o corpo como costuras topográficas subtis, as suas placas de cílios compostos demasiado finas para se resolverem a esta distância mas coletivamente impondo uma textura estriada que o feixe rasante azul transforma em micro-sombras. A neve marinha deriva continuamente através do cone de iluminação — partículas brancas de tamanho variável, umas em foco nítido, outras desfocadas em halos suaves — envolvendo o animal numa queda de neve lenta que dá profundidade mensurável à água negra e transforma esta coluna de escuridão fria e salgada num espaço habitado e vivo.

O olhar paira suspenso sobre o polo aboral de um *Mnemiopsis leidyi* vivo, dominado por uma cúpula hemisférica de cerca de duzentos micrómetros de diâmetro — uma estrutura de vidro biológico tão opticamente homogénea que apenas se revela pela fina crescente de luz azul-branca que a sua curvatura projeta, dobrando a iluminação oceânica transmitida através de colagénio e glicoproteínas. No interior dessa cúpula suspende-se o estatólito, um agregado compacto de grânulos de carbonato de cálcio de tom marfim suave, pousado em aparente imobilidade sobre quatro leques de cílios equilibradores — as células ciliadas balanceadoras cujo batimento a frequências de quinze a trinta e cinco hertz as dissolve em véus de luminescência perlada, demasiado rápidas para se fixarem em qualquer instante. Quatro sulcos ciliados irradiam da base da cúpula como os braços de uma rosa dos ventos, as suas superfícies cobertas de cílios mais finos que produzem um leve cintilo iridescente em verde e prata quando a luz rasante os encontra em ângulo, prolongando-se até às primeiras filas de ctenas que emergem ao longe como ribbons luminosos de azul-esverdeado. A mesoglea que serve de paisagem a esta cena — um gel viscoelástico de profundidade transparente, composto de noventa e sete por cento de água com fibras de colagénio que apenas se denunciam por um trémulo refrativo ao menor desvio do ângulo de visão — dissolve-se na distância numa névoa de azul cobalto, enquanto além da parede curva do animal o oceano aberto se apresenta como um vazio luminoso e absoluto, conferindo à cúpula cristalina diante de nós a qualidade simultânea do íntimo e do infinitamente suspenso.

Suspensos a milímetros abaixo da superfície do oceano ao amanhecer, contemplamos uma larva cydippid de dois milímetros que flutua como uma esfera de vidro soprado — tão opticamente transparente que se revela menos como um corpo sólido e mais como uma perturbação suave na luz, uma lente viva que curva os raios de ouro pálido e azul frio que filtram através da microcamada superficial. Ao longo do seu corpo esférico, oito fileiras de cílios rudimentares traçam meridianos fantasmais, emitindo lampejos prismáticos de rosa a violeta enquanto batem em ondas metacrónicas — não bioluminescência neste amanhecer, mas cor estrutural pura, a assinatura efémera de cílios em movimento sobre fotões de aurora. À volta da larva, o que parece ser um universo denso e habitado: uma cadeia de diatomáceas *Chaetoceros* deriva como um comboio de lanças de vidro dourado-âmbar, cada célula de sílica com o tamanho de um melão à nossa escala reduzida, os seus espinhos hialinos dispersando a luz em agulhas prismáticas; e mais além, um nauplius de copépode roda sobre si mesmo como uma nave de exploração alienígena, o seu carapaça em cobre-alaranjado a captar o brilho difuso da superfície com o olho único e obsidiano fixo num horizonte aquático invisível. A neve marinha — agregados de muco, fragmentos de detritos, células vivas e carapaças vazias em âmbar, verde-oliva e ouro pálido — deriva em todas as direções, transformando a coluna de água numa catedral suspensa de partículas luminosas onde esta larva quase inexistente já estende dois tentáculos de seda para provar, silenciosamente, o mundo que a rodeia.

Suspenso a quarenta metros de profundidade, o observador paira numa coluna de água que transita imperceptivelmente do azul-cerúleo acima para um verde frio e denso abaixo, e precisamente ao nível dos olhos a termoclina impõe-se como um espelho horizontal e trémulo — uma lente contínua de distorção refractiva que torce os contornos distantes tal como o calor de verão ondula sobre o asfalto, marcando o encontro entre duas massas de água com densidades, temperaturas e composições ópticas distintas. Os feixes de luz da tarde filtram-se desde a superfície em cones de ouro pálido que se atenuam e dissipam antes de atravessar essa fronteira, deixando a água mais fria mergulhada num crepúsculo esverdeado e particulado onde copépodes alaranjados cintilam como brasas em suspensão, concentrados na interface de densidade onde o gradiente de temperatura retém os nutrientes e a biomassa. Três *Mnemiopsis leidyi* — ovais achatados de transparência quase perfeita, cada um com cinco a oito centímetros de mesogleia cujo índice de refracção quase iguala o da água do mar — flutuam imóveis precisamente na linha da termoclina, a metade superior banhada em luz dourada filtrada, a metade inferior imersa no verde-escuro frio, a fronteira refractiva cortando cada corpo como uma régua traçada através de vidro. Ao longo dos flancos de cada animal, as oito fileiras de pentes irrompem em vagas lentas de cor estrutural — rubis profundos deslizando para âmbar, âmbar para verde-ácido, verde para índigo e índigo de volta a violeta — enquanto as placas de cílios batem em ondas metacrónicas que percorrem cada fileira em dezenas de milissegundos, impulsionando a água e criando redemoinhos invisíveis que perturbam levemente os flocos de neve marinha à deriva na camada de transição. Os lobos orais, abertos e estendidos para a zona mais fria e rica em presas, tremem com cílios microscópicos que aspiram a água carregada de copépodes para cima, através da fronteira da termoclina, num acto de predação tão silencioso e preciso que o único indício da sua violência são as pequenas chamas laranja que, uma a uma, desaparecem do crepúsculo verde abaixo.

Suspenso no interior de uma larva cydippid de *Mnemiopsis leidyi* com apenas 2 milímetros, o observador flutua numa escuridão absoluta atravessada por uma teia não hierárquica de filamentos magenta — a rede nervosa marcada com FMRFamide — que irradia em todas as direções sem centro discernível, cada fibra pulsando com o calor de coral saturado e projectando halos suaves de rosa difuso no vazio circundante. Oito arcos de verde-ácido curvam-se sobre a cabeça como as nervuras de uma abóbada de catedral, as bandas serotonina-positivas das fileiras de pentes traçando meridianos paralelos que impõem uma gramática espacial à anarquia da rede filamentosa abaixo, as duas fluorescências fundindo-se em intersecções âmbar-branco onde se cruzam. No zénite, o órgão apical — o estatocisto sensorial que governa a orientação gravitacional do animal — arde como uma auréola circular perfeita de magenta e verde em co-localização, o anel de brilhância quase branca rodeando um centro levemente mais escuro como o olho de uma tempestade e banhando em violeta-rosa difuso os filamentos nervosos mais próximos. Pontos azuis de DAPI — cada núcleo celular uma esfera cerúlea discreta — derivam a profundidades irregulares pela mesogleia como um campo estelar disperso dentro de um gel viscoelástico que é 97% água e quase indistinguível opticamente do próprio oceano. O interior de um animal menor do que uma semente de romã revela-se, visto assim, com a escala sentida de uma nebulosa à deriva.

Suspenso no interior do kreisel, iluminado por colunas de luz LED azul-branca que sobem do fundo do tanque como um sol artificial, o *Mnemiopsis leidyi* preenche todo o campo visual como uma escultura de vidro vivo em suspensão perfeita — seis centímetros de mesogleia, esse gel viscoelástico de colagénio e glicoproteínas com índice de refração quase idêntico ao da água do mar, tornando o corpo da criatura numa transparência que dobra a luz em caustics suaves e mutáveis. As oito fileiras de ctenas arqueiam-se em sequência ao longo do corpo como nervuras luminosas, cada prato composto por dezenas de milhares de cílios fundidos em leques franjados que batem em ondas metacrónicas antipleticas a dezenas de hertz, e a geometria pura dessas estruturas transforma a luz branca em fogo espectral — vermelho a sangrar em âmbar, verde em azul-esverdeado elétrico, violeta a desaparecer de novo em azul — não por pigmento algum, mas por difração pura. Os lobos orais contraem-se e expandem-se em ritmo peristáltico lento, tecido gelatinoso translúcido com margens franjadas que capturam a luz ascendente e a aureolam em lavanda pálida, enquanto dois cordões gonádicos cor de coral e âmbar correm em paralelo ao longo do estomodeu, únicos volumes opacos neste organismo que é noventa e cinco por cento água. A parede escura e curva do tanque desenha no fundo um horizonte negro e circular, e uma ponta de pipeta desfocada na margem da imagem — enormíssima, planetária nesta escala — lembra que este universo de vidro vivo cabe na palma de uma mão.

Você flutua ao lado de um tentículo de *Pleurobrachia* no escuro mesopelagial, suspenso numa escala onde um fio de vidro biológico preenche todo o seu campo visual — o tentículo lateral, não mais espesso que um cabelo de cristal, vibra em meia-retração como uma mola de relojoeiro prestes a disparar. Ao longo do seu comprimento alinham-se os coloblastos: estruturas adesivas em forma de cúpula, cada uma com seu filamento espiral enrolado no núcleo do fio e sua cabeça repleta de grânulos refrativos que captam a luz azul-negra ambiente e devolvem clarões frios de branco-dourado — alguns ainda intactos e esféricos, outros achatados em discos de contato irreversível contra a quitina do copépodo. A antena de *Calanus* irrompe pelo campo visual como uma viga de âmbar enferrujado, suas cristas transversais e setas marginadas pelo brilho lateral da coluna de luz difusa que desce da superfície distante a cerca de 480 nanômetros, enquanto dois ou três coloblastos esmagados contra a cutícula do crustáceo liberam halos adesivos quase invisíveis — a bioquímica da captura congelada no instante exato da adesão. A pata natatória do copépodo, translúcida e emplumada, empurra inutilmente contra o fio, e a leve curvatura do tentículo registra a força desse esforço desesperado antes que a musculatura do ctenóforo retome o encurtamento — violência íntima e silenciosa, legível apenas na geometria da tensão entre os dois corpos.
