Confiança científica: Alto

O mergulhador flutua imóvel dentro de uma tempestade de vidro vivo: em todas as direções, barris de *Salpa thompsoni* pulsam com um ritmo lento de dois batimentos por segundo, tão densamente distribuídos que cada meio metro de linha de visão é interrompido por outro corpo gelatinoso orientado ao acaso, transformando o oceano subantártico numa névoa nacarada e opalescente em que a transparência coletiva não revela o fundo, mas apaga a distância. Cada zoide tem o tamanho aproximado de um punho ou de um antebraço: um cilindro muscular oco e perfeitamente translúcido, cujas bandas de musculatura circular aparecem apenas como um frémito de refração contra a água escura ao fundo, enquanto o intestino cor de âmbar dourado — carregado de fitoplâncton ingerido — brilha como um braseiro interior, e ao lado dele um gónada rosa-coral flutua em suspensão como uma joia à deriva. A centenas destes núcleos quentes dispersos no azul-cinzento polar convertem a coluna de água numa lanterna gigantesca sem limites visíveis, cujo campo distante se dissolve em branco nacarado impenetrável a menos de cinco metros. Cortando esse mundo de gel em queda cadenciada e implacável, peletes fecais verde-acastanhados — cilindros densos e membranosos do tamanho de sementes pequenas — descem em trajetórias quase verticais como balas em câmara lenta, sólidos e saturados contra a palidez translúcida dos salpas, transportando para o abismo cinzento-azulado abaixo o registro comprimido de todo o fitoplâncton que o bloom filtrou, num fluxo de carbono que a superfície produz sem cessar, indiferente e contínua.

Suspenso na escuridão absoluta do oceano aberto, o olhar encontra uma esfera de três centímetros que mal existe — um invólucro de mucopolissacarídeos tão perfeitamente ajustado ao índice de refração da água que se revela apenas como uma distorção leve da treva, um arqueamento quase imperceptível da nada. Quando o feixe oblíquo a encontra, a casa de *Oikopleura dioica* irrompe em iridescência: bandas de interferência em azul ártico, âmbar dourado e verde-espuma reorganizam-se continuamente sobre a membrana curva, produzidas por variações de espessura da mucosa na ordem de micrómetros que transformam toda a superfície num reticulado de difração esférica e vivo. Para além do crescente iluminado, a esfera recua para uma sombra translúcida onde um pulso carmesim difuso — o coração da larvácea batendo por detrás da parede do tronco — brilha com a discreta insistência de uma brasa vista através do nevoeiro. Mais fundo, o notocorda vítreo traça os seus arcos sinusoidais lentos, e com cada batida da cauda as chicanas internas de mucosa estruturada captam o feixe e revelam-se como planos gossamer em suspensão, espalhando uma névoa opalescente que torna brevemente legível a câmara de filtração — uma catedral nanoscópica em gel que funciona sem falha há quinhentos milhões de anos. Grãos de neve marinha derivam pela coluna de água em descida gravitacional, cada um um lampejo breve de branco quente antes de regressar ao abismo, enquanto as bandas iridescentes continuam a reorganizar-se e o coração pulsa dentro da esfera de quase-nada.

Você flutua imóvel a quinze metros de profundidade, olho a olho com uma cadeia viva de *Thalia democratica* que se estende do limite do seu alcance para um infinito cobalto tão saturado que parece menos oceano e mais o interior de uma estrela azul. Cada zoóide do agregado pende na coluna como um barril de vidro soprado — transparente quase ao ponto do inexistente — e da proximidade de poucos centímetros é possível resolver toda a arquitectura interna através da parede corporal: oito aros musculares fantasma encerram cada cilindro como sugestão de estrutura, enquanto no centro geométrico de cada barril um intestino âmbar-dourado brilha como se guardasse um filamento aquecido, fruto de uma manhã inteira de filtração contínua de fitoplâncton. Flanqueando o intestino, gónadas lenticulares cor-de-rosa flutuam como gotas de quartzo rosado, e num polo anterior de cada zoóide um coração carmesim do tamanho de uma cabeça de alfinete pulsa visivelmente a dois ou três batimentos por segundo, projetando cintilações de cor mais profunda através da parede de gel — a totalidade deste maquinário vivo construído em tecido cuja diferença de índice de refração relativamente à água circundante é tão ínfima que os animais existem menos como matéria sólida do que como perturbações ópticas da água que os compõe. A luz solar, reestruturada por quinze metros de oceano em rendilhado geométrico de causticas azul-brancas em deslocamento contínuo, projeta-se através de cada barril e completa os seus padrões no vazio além, transformando a cadeia inteira numa catedral suspensa no espaço negro-cobalto onde não existe substrato, nem fundo, nem fronteira visível — apenas água como espaço puro, e estas arquitecturas de gel quente mantendo âmbar, carmesim e branco-fantasma contra a infinitude fria.

O olhar dirigido para cima encontra uma estrutura que mal parece ter existido — uma casa de larváceo abandonada há minutos, descendo lentamente pelos duzentos metros da zona mesopelagica onde a luz do dia se estreita num único tom de índigo profundo, como o último instante antes do anoitecer definitivo. A membrana de muco, que horas atrás era uma esfera perfeita de quase quarenta centímetros, dobra-se sobre si mesma de forma assimétrica, um hemisfério cedendo em pregas elegantes enquanto o outro resiste ainda à forma de cúpula, e onde a superfície permanece tensa ela fragmenta a luz em bandas iridescentes de violeta, ouro e azul que percorrem o tecido à medida que a estrutura roda em câmara lenta. Esta "casa" é inteiramente feita de mucopolissacarídeos secretados por células glandulares do animal que a habitou — um filtro biológico de poros entre 0,2 e 2 micrómetros, capaz de reter bactérias e picofitoplâncton demasiado pequenos para a maioria dos predadores — e o que agora parece névoa leitosa no interior da membrana é precisamente o acúmulo dessas partículas capturadas, carbono orgânico embalado num invólucro que o transportará, silenciosamente, para o fundo. Na grelha de entrada ainda visível como um rendilhado geométrico de fios de muco, copépodes de um milímetro — olhos cor de âmbar quente, corpos tão transparentes que a água azul os atravessa — exploram a malha entupida com apêndices demasiado finos para serem resolvidos pela distância, testemunhas minúsculas de uma transferência de matéria que, multiplicada por oceanos inteiros, move mais carbono para as profundezas do que qualquer outro mecanismo biológico conhecido.

Você está suspenso dentro de uma catedral de âmbar vivo, envolto em todas as direções por uma malha ortogonal de fibras de gel que se estende até um horizonte dissolto em névoa dourada — a rede mucosa da cesta faríngea de uma salpa, secretada pelo endostilo e esticada em filamentos cilíndricos de mucopolissacarídeos com pouco mais de um micrómetro de diâmetro. Cada fio capta a luz quente que filtra através do corpo translúcido do animal e a redistribui como vidro de mel fundido, os pontos de cruzamento formando janelas retangulares de cerca de um micrómetro de abertura — aberturas perfeitamente dimensionadas para aprisionar as menores células fotossintéticas do oceano. Numa dessas junções próximas, uma esfera de *Prochlorococcus* encravou-se entre dois fios como um meteorito borgonha-escuro, os seus 0,6 micrómetros suficientes para a tornar proporcional a um bloco de pedra neste cosmos miniaturizado, enquanto uma haste bacteriana pálida se adere ao fio adjacente através de interações eletrostáticas de superfície. Uma corrente invisível, movida pelos cílios do cesto faríngeo que batem a dezenas de vezes por segundo, arrasta água rica em picoplâncton através desta geometria paciente e repetitiva, transformando a própria estrutura do gel numa armadilha biológica que alimenta a salpa e, ao depositar fezes densas no fundo oceânico, bombeia carbono atmosférico para as profundezas.
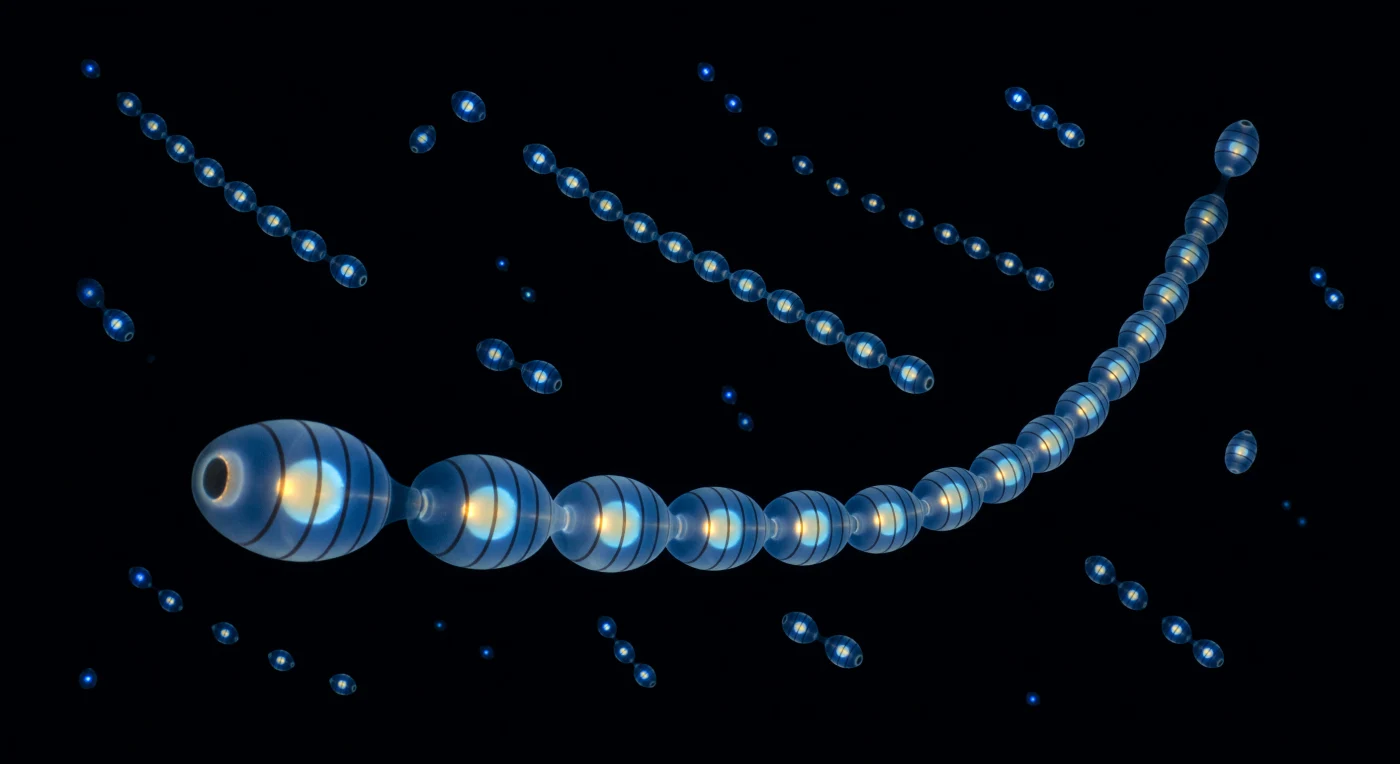
A corrente de salpas estende-se diante de você como um lustre vivo suspenso no vazio absoluto, cada zoóide um barril de gel do tamanho de um polegar que acende em sequência numa onda azul-branca fria propagando-se de anterior a posterior a 476 nanómetros — uma cor tão pura e cirúrgica que parece menos visível do que sentida. Dentro de cada barril iluminado, a arquitetura do organismo revela-se por transparência: oito bandas musculares circulares projetam-se como halos escuros contra o brilho interno, as suas sombras pressionando para dentro como as costelas de uma lanterna de vidro, enquanto o conteúdo intestinal flutua como uma silhueta âmbar aquecida — ouro queimado contra gelo azul — no centro de um animal que é em noventa e cinco por cento água do mar e que emite luz a partir da sua própria substância como uma estrela fria e contida. Ao redor e além da cadeia mais próxima, outras cadeias pulsam em dispersão tridimensional solta, as mais próximas legíveis como fios de contas, as intermédias como pontos de brasa, as mais distantes como faíscas estrelares indistinguíveis do cosmos, até que a água absorve os fotões antes que possam regressar e entre elas existe apenas o negro perfeito e irreversível do oceano de meia-noite. Esta explosão bioluminescente foi desencadeada por perturbação mecânica — a passagem de algo maior, uma onda de pressão, o eco de movimento — e a colónia responde com uma linguagem de luz sequencial cuja função permanece debatida: dissuasão de predadores, resposta de alarme em cascata, ou simplesmente a física inevitável de fotoproteínas excitadas em gel vivo.

Diante de você, a trezentos metros de profundidade onde nenhuma luz solar jamais penetra, flutua uma esfera de sessenta centímetros que parece ter sido soprada em vidro vivo — a casa de *Bathochordaeus charon*, uma estrutura secretada inteiramente por um único animal em poucas horas, construída de mucopolissacarídeos tão finos e tão próximos do índice de refração da água que sua existência só se revela pela iridescência tênue que percorre sua superfície sob a iluminação lateral do ROV. O interior organiza-se em camadas concêntricas de filtros de muco — bafles geométricos de malha progressivamente mais fina, capazes de reter partículas de apenas dois micrómetros, capturando bactérias e picofitoplâncton que escapariam a qualquer outro filtrador do oceano — enquanto ao centro o próprio larváceo pulsa em âmbar-laranja, sua cauda em chicotada lateral lenta bombeando correntes contínuas através de toda essa arquitetura mucosa. Pequenos copépodes de olhos carmesins pairam imóveis junto à grade de entrada anterior, atraídos pela corrente gerada mas impedidos por ela mesma de recuar, num equilíbrio tenso entre predação e fuga. Quando esta casa for abandonada — em poucas horas, entupida pelo próprio sucesso da filtração — afundará como um pacote compacto de carbono orgânico em direção ao fundo abissal, carregando consigo tudo o que capturou e tornando este organismo efémero num dos mais eficazes vetores de exportação de carbono do oceano profundo.

Você está deitado sobre a planície abissal a 3 000 metros de profundidade, o rosto voltado para cima em direção a uma coluna d'água negra e sem forma, enquanto uma procissão lenta e esparsa de pellets fecais de salpa — cilindros oliva-escuros de meio a dois milímetros revestidos de biofilme bacteriano como veludo estipulado — desce em espirais glaciais em sua direção, cada um captando a única luz disponível como um filete de brilho azul-cerúleo na borda curva de sua membrana. Entre eles flutuam carcaças de salpas em colapso, fantasmas de papel de seda translúcido cujos arcos musculares se dissolveram em véus de gel, cada corpo rodeado por uma auréola difusa de matéria orgânica dissolvida que turva a água ao redor como respiração sobre vidro frio. Abaixo, a lama biogênica cinza-bege — construída ao longo de milênios a partir de carapaças de foraminíferos, esqueletos de radiolários e neve marinha compactada — registra a chegada de cada pellet em minúsculas crateras de impacto, os terminais físicos de uma viagem de semanas desde a zona fótica iluminada pelo sol. Às margens do campo visual, silhuetas pálidas de holotúrias jazem semi-submersas no sedimento, imóveis como ossos antigos, enquanto as halos de matéria orgânica se expandem à velocidade da difusão molecular nesta câmara terminal da bomba biológica — um mundo sem cor além desta única frequência fria de azul, sem turbulência, sem estações, apenas a chuva escura e paciente de carbono fixado descendo para um assoalho quieto e receptivo.

O observador flutua suspenso numa camada de água azul-safira tão pura que a transparência em si parece uma substância — não o vazio entre as coisas, mas um meio luminoso com espessura e peso próprios. Neste campo imóvel, esferas rubras de *Prochlorococcus* distribuem-se em todas as direções com uma regularidade quase cristalina, cada uma emitindo uma cálida fluorescência vermelha em resposta ao azul frio da excitação solar filtrada por dezenas de metros de coluna de água; a sua escassez aparente é enganosa, pois o olho acompanha a sua repetição por camadas e camadas até ao infinito azul-negro, percebendo então que este vazio é habitado com uma densidade que não tem equivalente terrestre. Entre elas, cilindros ligeiramente maiores de *Synechococcus* derivam em ângulos suaves, o seu interior aquecido por pigmentos ficobilissómicos num tom âmbar-alaranjado, enquanto bastonetes bacterianos quase invisíveis se revelam apenas como um frémito prismático ao longo das membranas. Ocasionalmente, um picoeucariota de dois a três micrómetros emerge como uma lanterna mais complexa — as suas membranas internas de cloroplasto formando câmaras concêntricas de ouro esverdeado — lembrando que, neste cosmos de luz biológica, a arquitetura celular é também uma forma de luminosidade.

Suspenso no interior líquido da casa de muco de *Oikopleura dioica*, o observador confronta uma única estrutura translúcida que ocupa todo o campo de visão: a cauda da larvácea, congelada estroboscopicamente a meio batimento numa curva em S de precisão orgânica perfeita, com apenas três milímetros de largura mas imensurável na sua complexidade óptica. Ao longo do eixo central dessa fita de vidro vivo, o notocorda revela-se como uma coluna de células vacuoladas em forma de disco biconvexo, cada uma funcionando como uma lente individual de citoplasma comprimido, empilhadas em sequência e concentrando a luz oceânica difusa azul-esverdeada numa linha luminosa fria e nítida que percorre toda a extensão da curva. As lamelas musculares estriadas que flanqueiam este eixo central apresentam um bandado transversal de densidade óptica discretamente superior, testemunho da ordem molecular dos sarcómeros — estruturas proteicas contrácteis organizadas em repetição regular que geram o batimento assimétrico responsável pela propulsão e pela filtração dentro da casa mucopolissacarídica. Nas margens externas da cauda, as membranas das barbatanas laterais existem apenas como franjas únicas de interferência difractiva, faixas iridescentes de violeta frio a prata que tocam o limite absoluto da resolução óptica da luz visível. No espaço aquoso circundante, micro-partículas e colóides alimentares descrevem trajectórias circulares residuais, registos fantasma das estruturas de vórtice deixadas pelo batimento mais recente — o fluido ainda a relaxar para a imobilidade, devolvendo ao silêncio um movimento que existiu apenas milissegundos antes.

Suspenso a dois centímetros da face anterior de uma *Pegea confoederata*, o observador contempla uma das arquitecturas biológicas mais transparentes do oceano: um barril de gel puro, oito centímetros de comprimento, cuja parede de túnica se trai apenas como uma ténue curvatura do azul-verde que a envolve, índice de refracção tão próximo da água do mar que o limite entre organismo e oceano existe mais como conceito do que como superfície. A luz de meio-dia tropical, filtrada por quarenta metros de coluna de água e chegando de todas as direcções do hemisfério superior em simultâneo, atravessa o corpo inteiro e transforma-o numa lanterna viva, revelando a anatomia completa como uma natureza morta luminosa: o cesto faríngeo arde em âmbar dourado, a sua rede de muco capturando e dispersando a luz em iridescência aquecida; o endóstilo traça uma risca ventral de resina iluminada ao longo de todo o comprimento do barril; o lobo gonadal alaranjado contém oócitos individuais visíveis como esferas pálidas suspensas em tecido cor de âmbar. No centro dorsal, o coração — apenas três milímetros de tecido carmesim — contrai em ondas peristálticas lentas a cada meio segundo, cada sístole enviando uma onda visível de cor através do gel circundante, lembrando que este cristal flutuante e silencioso é um animal vivo, a filtrar bactérias picoplânctónicas do vazio azul e a fabricar, pellet a pellet, uma das correntes de carbono mais eficientes que o oceano produz.
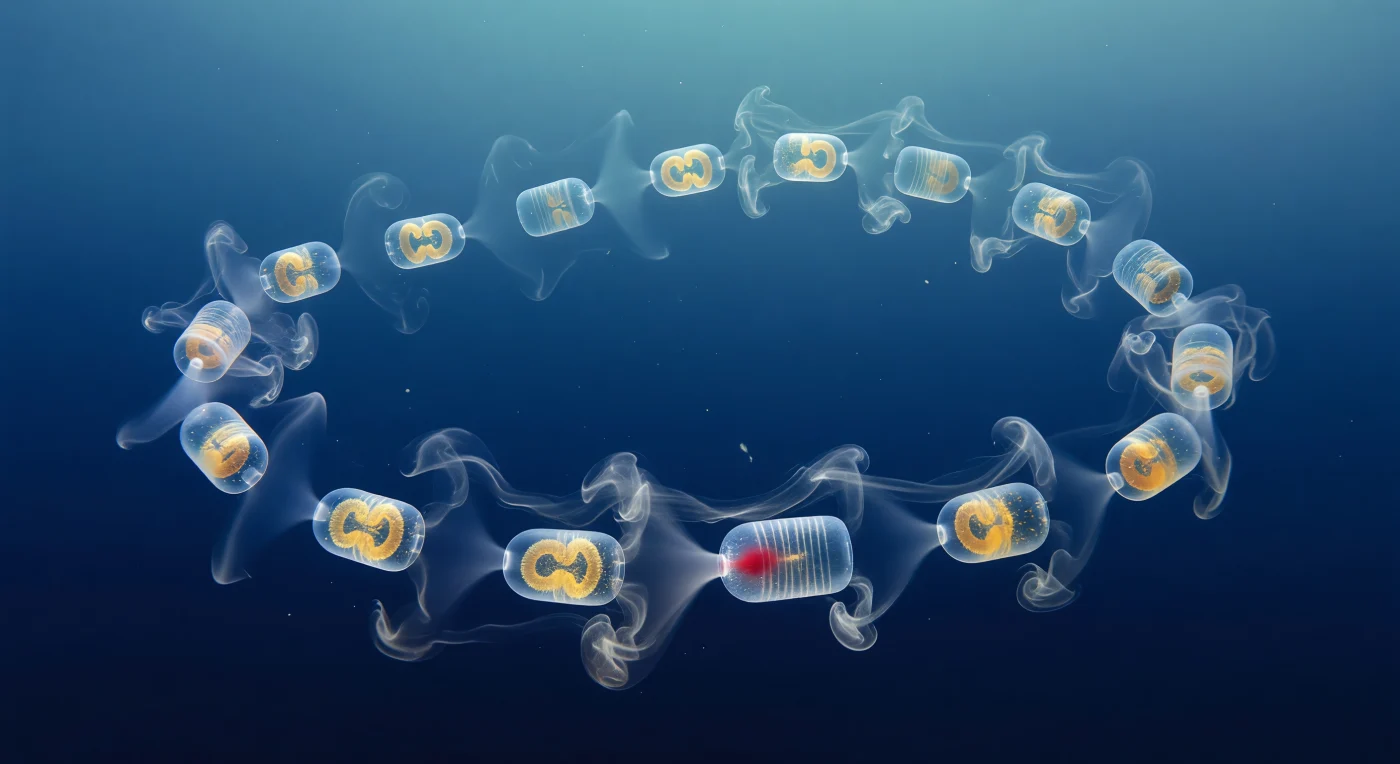
Você flutua a meia água diante de uma corrente viva de *Cyclosalpa affinis*, uma guirlanda âmbar de zoides cilíndricos cujas paredes tão transparentes que o azul cobalto do oceano atravessa cada barril quase sem obstáculo, traído apenas pelo halo prismático ténue onde a túnica curva dobra a luz e pelo brilho dourado-laranja do intestino repleto de fitoplâncton, suspenso como brasa dentro de um frasco de cristal. A óptica Schlieren transforma a própria coluna de água num meio visível: de cada sifão atrial emerge um penacho contínuo de água exalada ligeiramente mais quente, rendido pelo gradiente de densidade como fumo branco-prateado que se expande, se desfaz em filamentos laminares e se funde com o jato do zoide vizinho, tecendo uma esteira composta de fios refráteis entrelaçados que captam a luz descendente em breves cintilações iridescentes. Nos sifões orais opostos, a óptica revela o lado inverso do processo: concavidades de sombra subtis onde a água carregada de partículas é aspirada para dentro da câmara filtrante, formando um cone de influxo opticamente distorcido que se projeta para a frente como um penacho invertido e lento. A cadeia curva-se em arco helicoidal pelo enquadramento, os zoides próximos nítidos como vidro lapidado e os distantes dissolvidos em formas fantasmagóricas pelo azul-índigo da coluna de água, enquanto flocos de neve marinha derivam no plano médio, um deles desviando-se visivelmente à volta da rede de muco da cesta faríngea — estrutura quase inexistente, visível apenas como uma grelha de sombra de quase-nada.

Você está suspenso a poucos milímetros do tronco gelatinoso de uma *Oikopleura longicauda*, um organismo menor que um grão de arroz que flutua no vazio cerúleo da coluna d'água como um berlinde de vidro vivo, seus órgãos internos — o intestino âmbar, o rudimento rosado de gônada — visíveis através de um corpo que é, na prática, água organizada em forma. Da superfície do tronco, em tempo real, células glandulares oikoplásticas extrude filamentos de muco branco-prateado mais finos do que teia de aranha, cada um captando a luz difusa azul-catedral como uma linha brilhante antes de absorver água do mar e se expandir imediatamente em membrana semitransparente, construindo uma nova casa de substituição três semanas depois do abandono da anterior — um ciclo que este animal repete durante toda a sua vida. A casa incompleta, já com cerca de três milímetros, apresenta nas regiões onde duas camadas de muco se fundiram e afinaram à espessura óptica precisa uma iridescência de interferência de filme fino: ouro pálido que desliza para um verde-azulado fantasma e depois para um violeta residual nas bordas, cores que se reorganizam enquanto a membrana respira e flexiona exatamente como petróleo em superfície calma. Na fronteira de crescimento ativo, o material ainda é francamente espumoso — uma renda de microbolhas de muco não fundido que captam a luz azul ambiente como pontos especulares suaves — e além dessa franja, uma névoa opalescente leve difunde-se pela água circundante, a assinatura visível de mucopolissacarídeos dissolvendo-se em gradientes brownianos lentos, tornando o volume imediato em torno do animal levemente fosco, como se o oceano aberto estivesse participando silenciosamente da construção.

O campo visual é dominado por uma bolsa de gel pálido murcha sobre o sedimento abissal — a carcaça de uma salpa achatada como um lenço de vidro amassado, coberta por manchas de biofilme bacteriano branco-giz que contrastam com a membrana translúcida ainda brilhante por baixo, e cujas estrias musculares circulares sobrevivem como linhas paralelas prata-acinzentadas, vestígios fantasmáticos da arquitetura corporal outrora viva. Três poliquetas marfim percorrem a superfície da carcaça com os seus palpos tentaculares em leque, varrendo ritmicamente o biofilme, enquanto cinco anfípodes translúcidos — os seus olhos compostos vermelhos a arder como garnets minúsculos num mundo quase monocromático — raspam material nas margens; um único braço de ofiuro avança desde a borda do sedimento, os ossículos calcários articulando-se em creme-marfim até que os pés ambulacrais âmbar tocam o gel. A luz ambiente é uma luminescência biológica azul-esverdeada difusa, sem fonte identificável, que torna a carcaça o objecto mais brilhante da cena e mergulha o lodo foraminífero circundante — repleto de testas esféricas e planispiraladas de calcário branco e âmbar pálido — numa penumbra de teal profundo que se dissolve rapidamente em escuridão absoluta. Este banquete silencioso no fundo do oceano representa o culminar de semanas de exportação de carbono: a carcaça de gel, quase invisível na coluna de água iluminada pelo sol, torna-se aqui, a 1 500 metros de profundidade, um pulso localizado de energia orgânica que sustenta toda uma comunidade de oportunistas abissais especializados em capitalizar a imprevisível chuva de detritos gelatinosos.

Suspenso a cinquenta metros de profundidade num oceano subtropical estratificado, o observador flutua dentro de uma coluna de água que transita do jade translúcido acima para um azul-noite tão saturado que se torna sua própria escuridão abaixo — e entre esses dois registros, organismos de vidro habitam um espaço que a luz solar ainda alcança, mas já não ilumina com nitidez, chegando apenas como cones difusos de luminosidade sem fonte, quase âmbar nas margens do campo visual. À frente, a casa de uma larvácea lê-se quase como ausência: uma geometria elipsoidal de salas e funis de entrada que só se revela pelas cores de interferência nas suas superfícies curvas — violeta pálido, ouro fugaz, um fantasma de azul-esverdeado —, enquanto o corpo do animal no interior pulsa a dois ou três ciclos por segundo, dirigindo uma corrente imperceptível de picoplâncton através de malhas de muco com poros medidos em frações de micrómetro, filtrando cianobactérias que nenhum outro zooplâncton consegue capturar. Ligeiramente mais fundo e à direita, uma salpa solitária de cinco centímetros liberta dois peletes fecais densos e envoltos em membrana que já iniciam a sua descida gravitacional em direção à escuridão — pacotes comprimidos de carbono orgânico que podem levar semanas a atingir o fundo abissal, exportando para as profundezas a energia fixada por células de meio micrómetro à superfície. E do azul-índigo abaixo emerge, lenta e inevitável, a silhueta prateada de um *Mola mola* de trezentos quilos com a boca aberta dirigida para cima, tornando visível numa única cena a ponte trófica mais improvável do oceano: de bactérias fotossintéticas invisíveis a estas lanternas de gel a um dos maiores peixes ósseos do mundo.
