Confiança científica: Alto

Diante de nós, o corredor de solo serpenteia para a direita numa penumbra quase absoluta, iluminado apenas pelo brilho biológico frio que emana da hifa de fungo micorrízico arbuscular — um cilindro vítreo de tecido vivo, sua parede translúcida com um leve reflexo verde como borossilicato sob luz polarizada, ocupando cerca de um quarto da largura total do poro. As paredes do túnel são construídas de plaquetas de esmectita sobrepostas em tons de cinza-prateado, finas como lâminas de navalha, com bordas captando difração de luz em reflexos de peltre e lilás, enquanto blocos maciços de quartzo translúcido — imensos do nosso ponto de vista, centenas de vezes mais largos do que nós — refratam a luminescência ambiente em chispas prismáticas sobre o piso mineral. Películas orgânicas âmbar cobrem cada superfície como verniz antigo, e nas suas reentrâncias se aglomeram colônias de bastonetes bacterianos cinza-bege, cada um apenas um quinto do diâmetro da hifa, formando tapetes de biofilme que abraçam o substrato mineral como uma neblina densa e imóvel. Através da parede da hifa, o citoplasma se move com a deliberação imperceptível de uma maré lenta — glóbulos lipídicos creme-branco e âmbar à deriva numa gel levemente luminoso que transporta carbono, sinais químicos e a continuidade viva de uma rede que, em algum lugar muito acima, conecta raízes de árvores inteiras. Três túneis laterais se abrem no fundo do corredor e recuam para uma escuridão absoluta e impenetrável, emoldurados por plaquetas de argila salientes, lembrando passagens de uma caverna marinha sem cartografia, comprimidas até à largura de uma única célula humana.
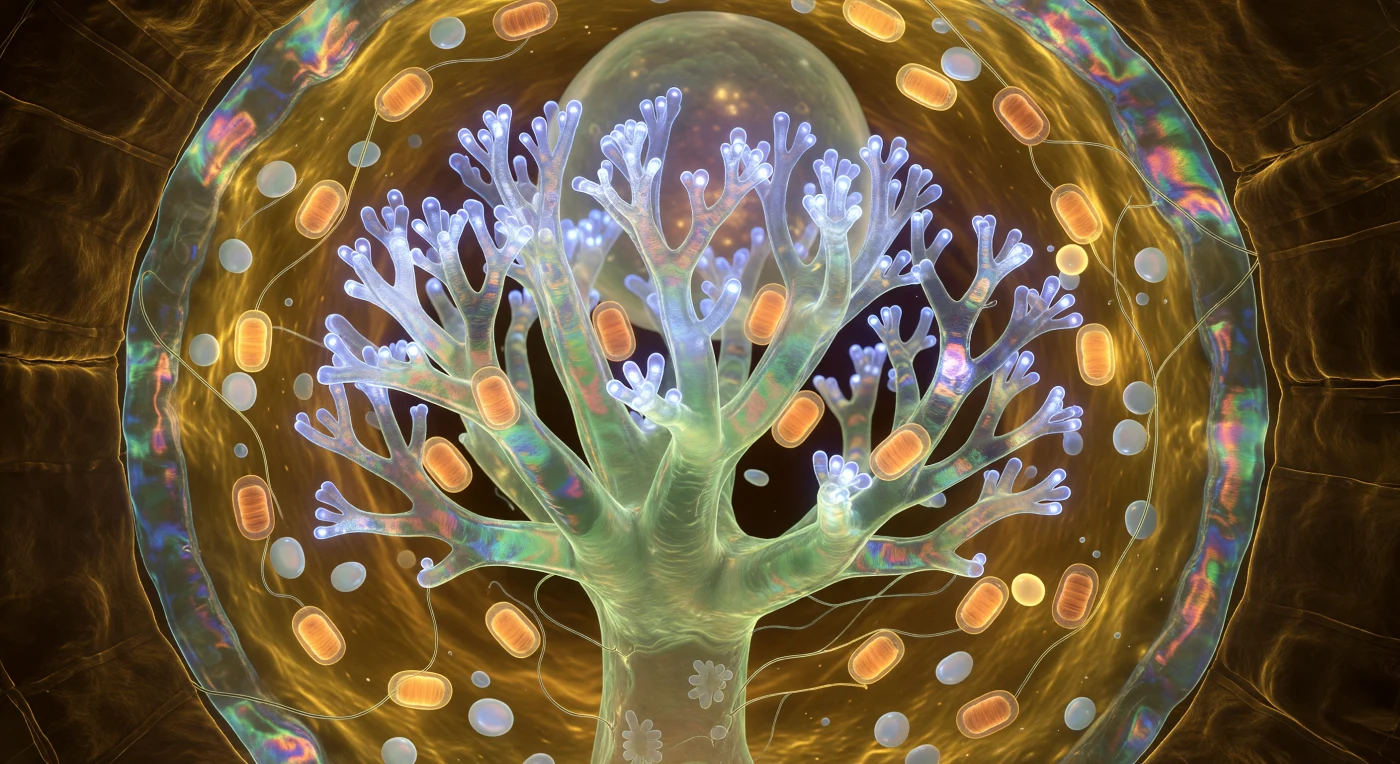
Você flutua suspenso no coração de uma catedral que nunca foi construída, apenas crescida: à sua frente, o arbúsculo eleva-se como uma floresta de coral invertida e congelada em pleno florescimento, seu tronco principal uma coluna translúcida de jade-marfim que se ramifica em dicotomias recursivas até os galhos terminais se dissolverem numa névoa trêmula de luminescência cobalto-branca — não há sol aqui, nenhum fóton do mundo macroscópico penetra esta profundidade de solo, e ainda assim tudo se ilumina pelo fogo metabólico frio da própria troca. O arbúsculo é uma estrutura fúngica que invade a célula cortical da raiz sem jamais romper a membrana hospedeira: a membrana periarbuscular — aquela dupla pele que envolve cada ramo com a lógica orgânica tensa de um filme de sabão — está literalmente cravejada de proteínas transportadoras de fosfato que ciclam suas conformações em ritmo contínuo, bombeando fósforo inorgânico para o citoplasma hospedeiro enquanto açúcares derivados da fotossíntese fluem em sentido contrário, sustentando desta única célula brilhante uma rede micelial que pode se estender por hectares acima de você. As mitocôndrias acumulam-se ao redor do tronco arbuscular como lanternas de âmbar, suas cristas internas visíveis como sulcos concêntricos através de membranas translúcidas, seus halos de respiração quimiosmótica se sobrepondo no citoplasma dourado e viscoelástico que contém amiloplastos perolados à deriva. A parede celular curva-se ao redor de toda a cena como uma muralha de âmbar escuro, suas microfibrilas de celulose em hélices cruzadas capturando o brilho arbuscular em baixos ângulos, revelando uma topografia de relevos e depressões como a face interior de uma abóbada de madeira esculpida — e na interface apoplástica entre os galhos mais externos e a membrana plasmática hospedeira, numa fenda de apenas cem nanômetros de largura, a troca que alimenta florestas inteiras acontece agora, em cascatas invisíveis, cujo resultado é toda esta arquitetura acesa ao redor de você.

Diante de você estende-se uma vasta planície mineral de penumbra, uma fatia de solo arenoso que se abre como um planalto desértico ao entardecer, atravessado por uma luminescência difusa que filtra do alto através dos estratos como luz âmbar a passar por vidro espesso. Em primeiro plano dominam três esporos de *Glomus*, esferas lacadas de âmbar e ocre do tamanho de rochedos à sua escala, com paredes multiestratificadas de caramelo translúcido e mel escuro visíveis como anéis concêntricos, enquanto ao lado um esporo de *Gigaspora* creme-rosado se ergue com a sua superfície verrugosa e ao fundo um esporo vinho-tinto de *Scutellospora* exibe o seu escudo de germinação em crescente, uma placa rígida de marrom escuro cuja aresta capta a luz com um brilho nítido. Entre eles, grãos de quartzo translúcidos e cristais de feldspato rosa — muitas vezes a sua altura — refratam a luz em cáusticas pálidas de branco e azul fantasma, enquanto flocos de matéria orgânica decomposta jazerem como pavimento de resina petrificada sobre o chão arenoso. Cruzando os vazios entre os esporos correm filamentos de hifa quase invisíveis, linhas de prata gossâmere que captam a luz apenas como fios brilhantes contra o escuro intergranular, suspensas em curvas catenárias sob a sua própria tensão superficial — arquitetura viva e delicada suspensa na escuridão permanente e geológica do solo.
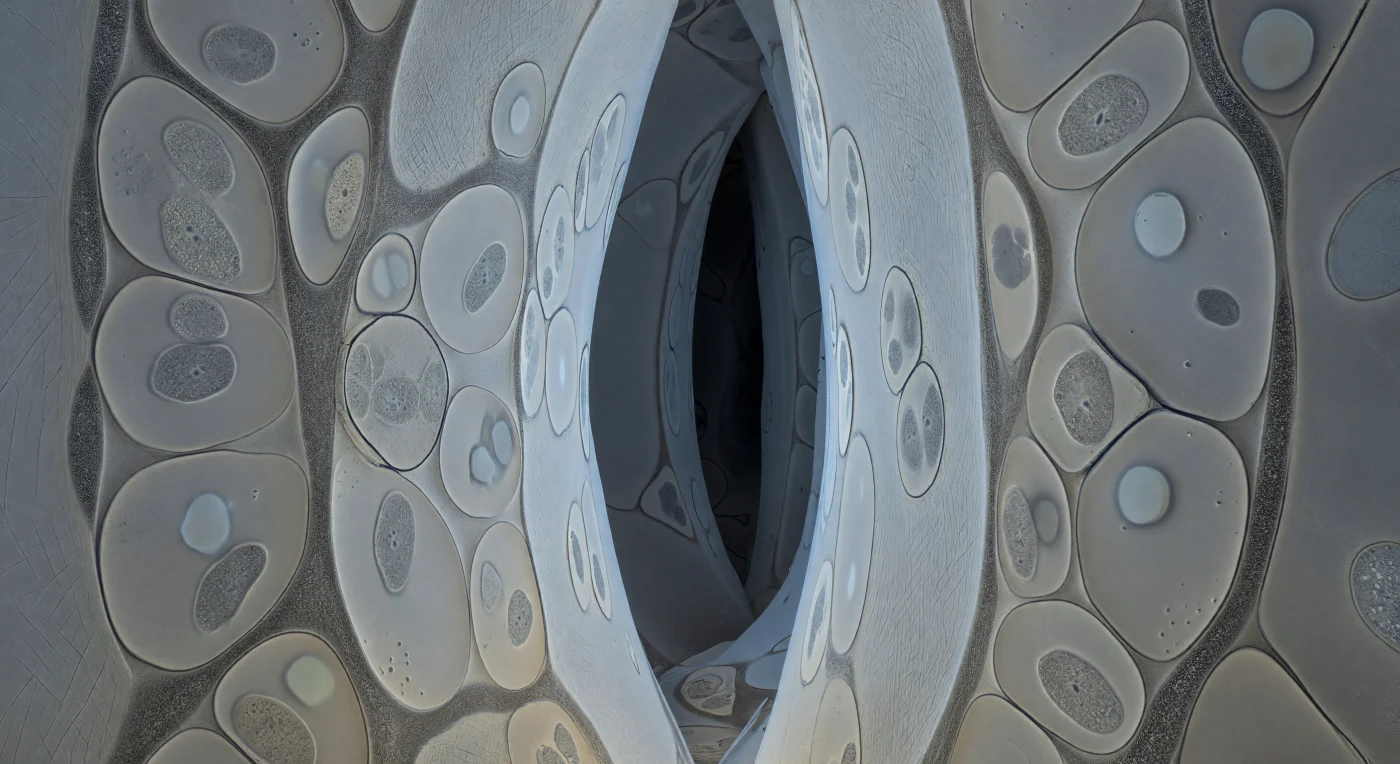
Você está suspenso dentro de um corredor tão estreito que as paredes o roçam por todos os lados — vastas superfícies de celulose cor de creme pálido erguem-se em lâminas levemente onduladas, cada feixe de microfibrilas correndo em diagonais cruzadas como o grão de um papel antigo comprimido, e a geometria global é aproximadamente hexagonal, mas orgânica e irregular, com passagens que se alargam em câmaras sombrias antes de se fecharem novamente em fendas mal mais largas do que o seu próprio corpo. As secções transversais das hifas fúngicas que preenchem esses corredores aparecem como formas lenticulares achatadas contra as paredes das células radiculares, os seus contornos de membrana plasmática traçados numa linha de grafite nítida e tensa, encerrando um citoplasma cinzento-pombo pontuado por mitocôndrias ovóides e vacúolos lúcidos que se lêem como ausências quase ópticas. Entre a parede fúngica e a parede da célula vegetal estende-se uma faixa granular de 15 a 25 nm — a matriz interfacial de glicoproteínas secretada simultaneamente por ambos os lados — que é o teatro invisível de toda a troca simbiótica: iões de fosfato abandonando o citoplasma fúngico, sacarose chegando da planta, num tráfego molecular que a este instante congelado se lê apenas como textura, nem membrana nem vazio, um limiar negociado. A paleta é quase monocromática — marfim frio para as paredes celulósicas, grafite-negro para os contornos de membrana, cinzento médio para o citoplasma, com apenas uma imperceptível nota âmbar onde gotículas lipídicas se agrupam numa secção hiphal — e o contraste estrutural é tão extremo que funciona como a sua própria iluminação, cada interface projectando uma sombra de bordo subtil que confere dimensionalidade a este labirinto interior sem necessidade de qualquer fonte de luz exterior.
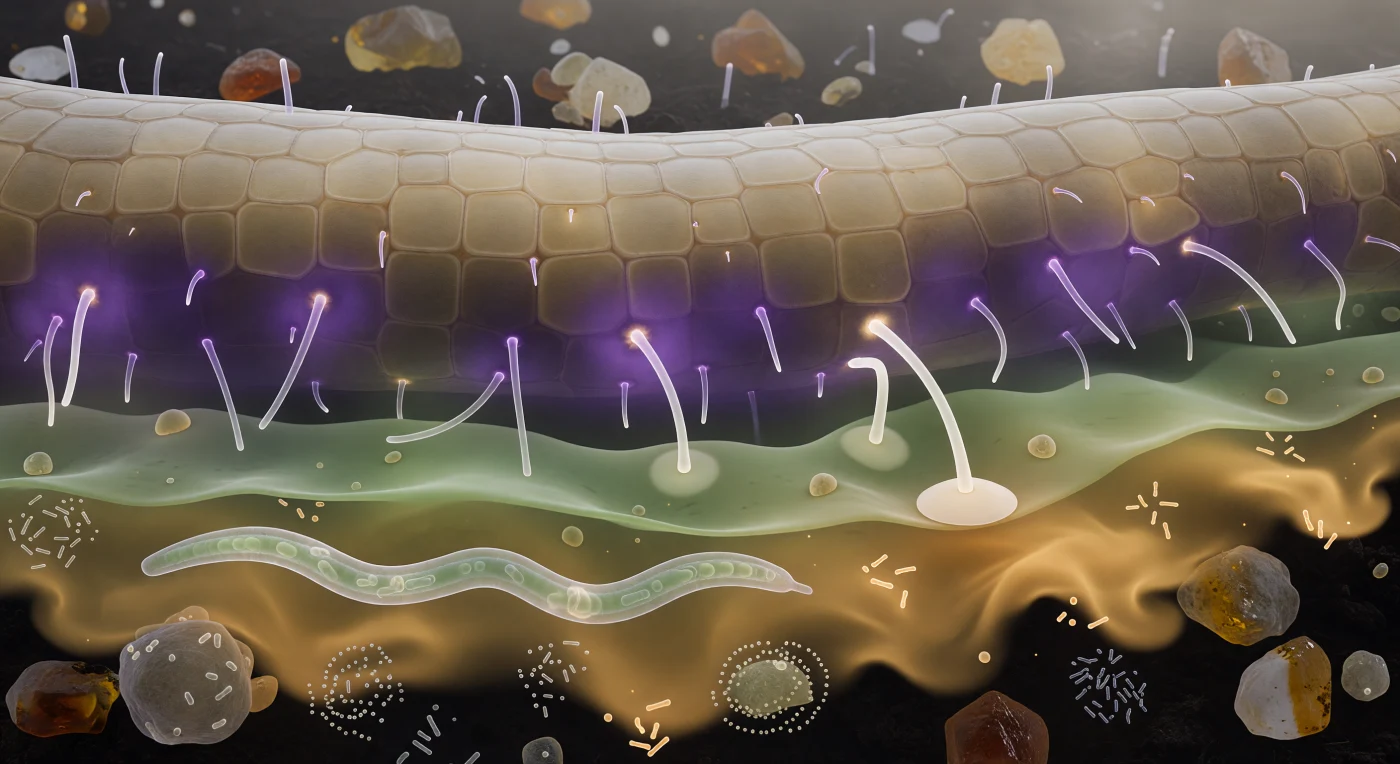
Você paira a meio caminho entre o solo mineral e a parede da raiz, e o que se vê não é escuridão, mas uma atmosfera química estratificada: uma coroa violeta densa de estrigolactonas e flavonoides abraça as células epidérmicas como névoa sobre lava, depois abre-se para um gradiente verde-acinzentado de aminoácidos que sangra em bandas de sálvia e espuma-do-mar pelo filme de água intersticial, e mais além uma pluma âmbar de exsudatos açucarados deriva até quase à sua face com a fluidez de uma corrente abissal luminescente. A raiz em si enche o campo visual superior como um penhasco de arenito pálido, cada célula epidérmica uma telha convexa de translúcido creme, os pelos radiculares projetando-se como cilindros de vidro que captam o brilho químico em interferência violeta ao longo dos seus eixos — estruturas especializadas em maximizar a superfície de absorção e em emitir os próprios sinais moleculares que atraem os fungos simbiontes. Na região mais sombria do enquadramento, uma ponta hifal de paredes marfim já arqueou em resposta ao gradiente e achatou a sua extremidade contra uma célula da epiderme numa estrutura discoide — o apressório —, o momento imediatamente anterior à penetração enzimática que iniciará a simbiose micorrízica; enquanto isso, um nemátodo translúcido atravessa a cena em ondulação sinusoidal, e enxames bacterianos em espiral pulsam sobre as faces dos agregados minerais, cada bactéria uma fração do diâmetro das hifas, lembrando que esta zona da rizosfera é simultaneamente sinal, alimento, campo de batalha e ponto de encontro evolutivo.

O observador encontra-se sobre uma superfície viva e tecida com precisão quase artesanal: uma manta plectênquimatosa de células fúngicas alongadas que se encaixam como cerâmica disposta à mão, creme nas suas zonas centrais e orladas de amarelo-enxofre pálido onde as membranas adjacentes se comprimem umas contra as outras, tudo isso revestido de um brilho pérola frio que emerge das próprias paredes celulósicas sem fonte de luz aparente. Por trás, curvando-se em ambas as direções e preenchendo o campo visual como o casco de um navio encalhado, a coluna radicular ergue-se como uma muralha de tecido castanho-tostado de superfície lisa e estriada, tão vasta na sua curvatura que a extremidade distante se dissolve em neblina antes de desaparecer completamente — é a epiderme da raiz, e a manta adere a ela com intimidade molecular, enquanto no interior invisível a rede de Hartig tece a sua labiríntica penetração intercelular. Aqui, a gravidade perde sentido: as forças de adesão e tensão superficial que colam o observador à superfície fúngica superam em ordens de magnitude qualquer tração descendente. Na franja da manta, a apenas alguns passos à frente, o tecido se desfaz em hifas individuais que partem para a escuridão absoluta do solo não colonizado como filamentos de fibra ótica — tubos transparentes de três a seis micrómetros de diâmetro, refrativos como vidro, com o citoplasma visível como um fio ténue de movimento interno onde organelos e vesículas transitam em direção ao ápice em avanço —, cada um deles contrastando com o negro total do vazio poroso do solo numa imagem de profundidade sem luz, percebida apenas pelo modo como esse brilho pérola se extingue ao longe entre as silhuetas rugosas dos agregados minerais.
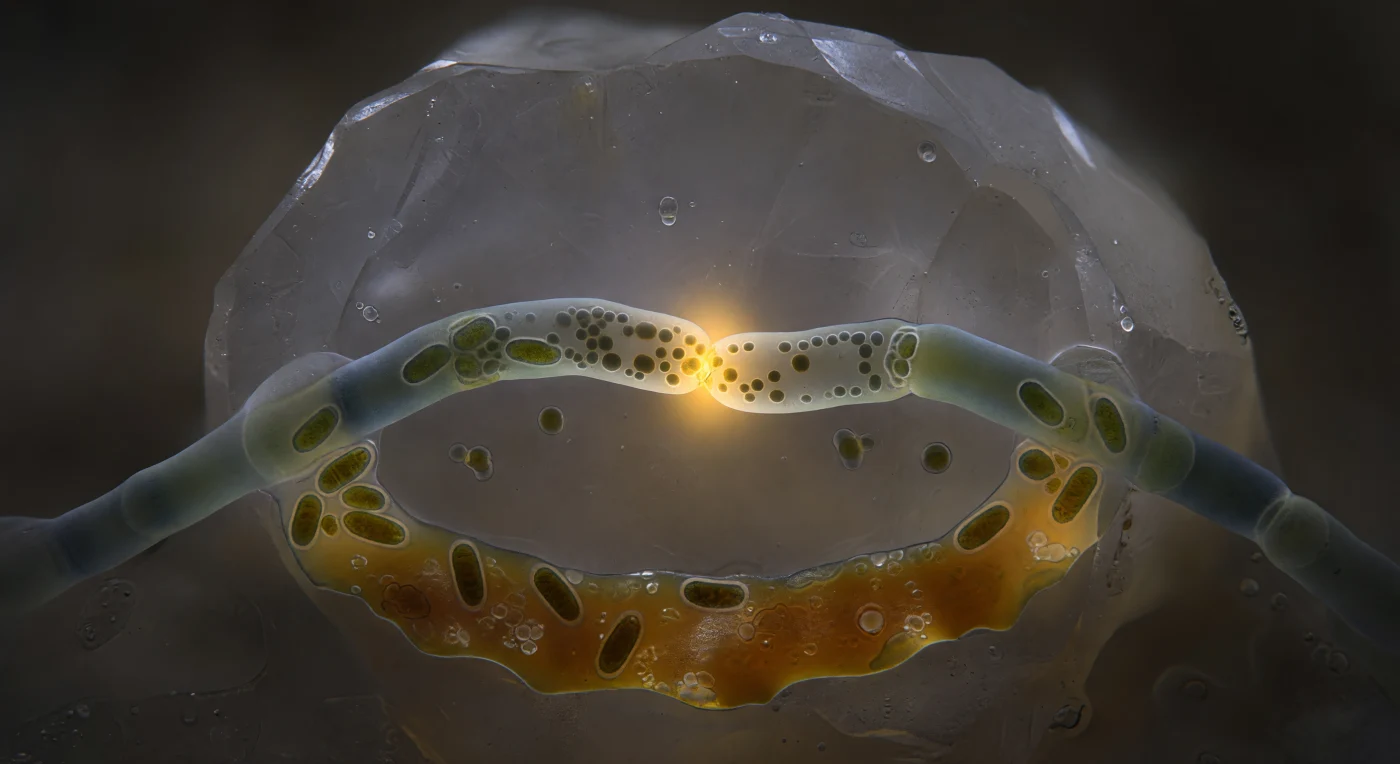
Diante dos seus olhos materializa-se um momento de fusão celular raramente testemunhado: duas hifas de fungo micorrízico arbuscular arqueiam-se uma em direção à outra como cabos de suspensão atraídos por uma tensão invisível, suas paredes de quitina-glucano translúcidas brilhando em verde-azulado pálido contra a escuridão absoluta de um poro de solo. No único ponto de contacto — um disco de apenas um micrómetro e meio — as paredes das duas células dissolveram-se mutuamente, apagando a fronteira entre dois organismos distintos num processo chamado anastomose hifal, pelo qual as redes fúngicas trocam núcleos, organelos e citoplasma para coordenar a colonização do solo e a partilha de recursos com raízes de plantas hospedeiras. O poro de fusão pulsa em âmbar-dourado intenso, alimentado por mitocôndrias densamente acumuladas nas suas margens, enquanto grânulos lipídicos e substância citoplasmática fluem em torrente viscosa da hifa esquerda para a direita, um transporte de nutrientes e sinais que poderá, em horas, influenciar uma rede que se estende por metros de solo florestal. Ao fundo, o grão de feldspato ergue-se como uma falésia polida por glaciares, e o filme de água do solo dispersa o brilho âmbar num halo difuso, fazendo deste poro escuro e minúsculo um centro metabólico de extraordinária intensidade.

O que domina o campo visual é um único macroa-gregado de solo de 3 milímetros, apresentado à distância de centímetros como se fosse um corpo planetário — a sua superfície negra-espresso, mosqueada de ocre quente, é o resultado de séculos de humificação, melanina fúngica e silicatos minerais fundidos numa geologia comprimida e densa. Sobre cada face exposta estende-se uma fina película de glomalina, proteína glicosilada excretada por fungos micorrízicos arbusculares, que confere à superfície um brilho âmbar-dourado ligeiramente hidrófugo — atestado pela gota de água que assenta perfeita e convexa num dos planos frontais, o seu ângulo de contacto alto e resoluto, reflectindo o mundo escuro do agregado como um espelho miniaturizado. Das fissuras que cortam a cintura equatorial da estrutura emergem filamentos hifais brancos em arcos paralelos, delgados como fio de seda, suturas vivas que mantêm coesos os grãos de quartzo e feldspato visíveis como inclusões pálidas semelhantes a blocos de gelo meio submersos na matriz orgânica. No canto inferior direito, sob iluminação ultravioleta, a glomalina abandona o seu dourado discreto e explode em fluorescência amarelo-verde incandescente, mapeando em relevo luminoso toda a arquitectura de carbono bloqueado nesta estrutura que os fungos do solo têm vindo a armar e a reparar há quatrocentos milhões de anos.
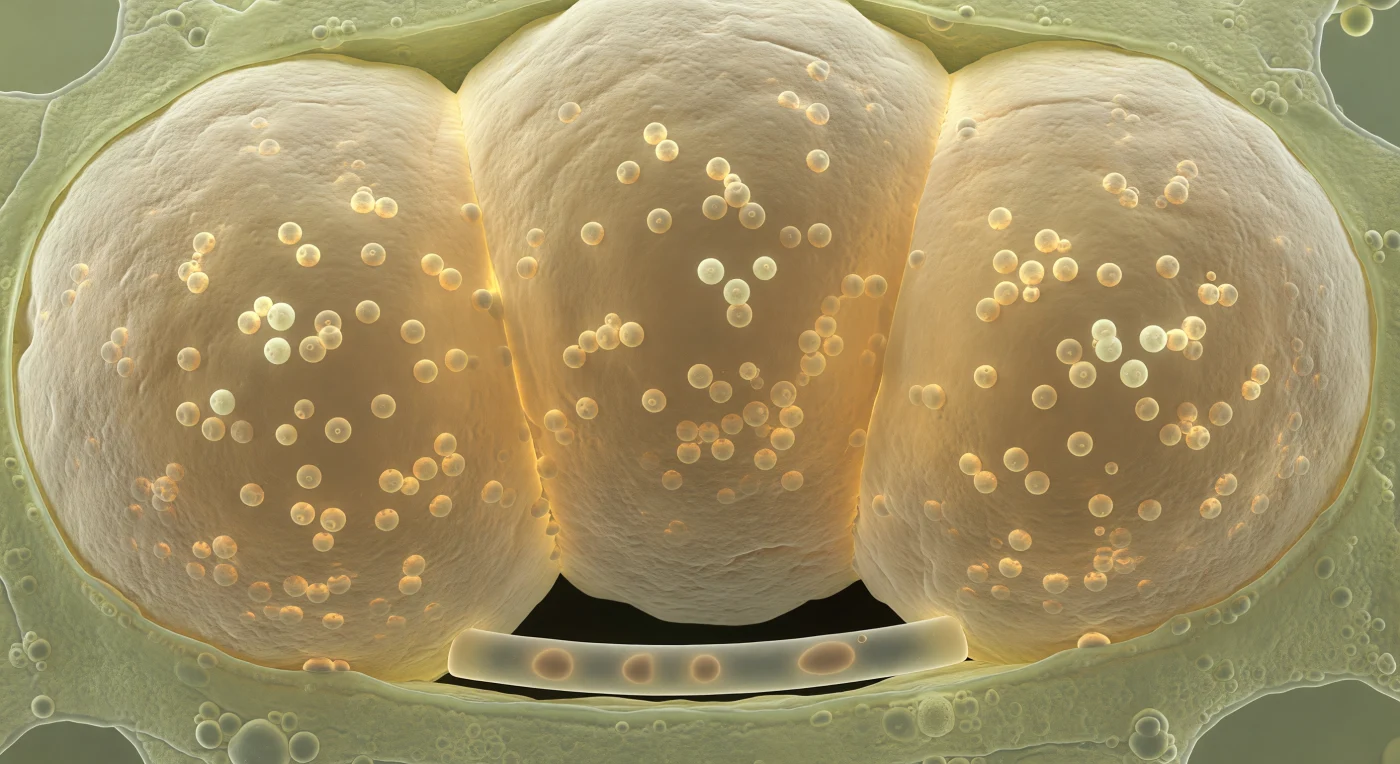
O interior desta célula cortical da raiz é uma câmara sufocante dominada por três vesículas de fungo micorrízico arbuscular que enchem o espaço como balões âmbar inflados até ao limite — as suas paredes multicamadas de quitina-glucano, rugosas como marfim gasto na superfície exterior e vítreas e luminosas por dentro, comprimem-se mutuamente ao longo de zonas de contacto achatadas, enquanto o citoplasma da célula hospedeira é espremido para as margens numa película translúcida e granular de ribossomas e organelos. Dentro de cada vesícula, dezenas de glóbulos lipídicos esféricos de cinco a dez micrómetros flutuam em suspensão imóvel, cada um refractando a luminescência âmbar ambiente num brilho interno dourado-pálido, como gotas de óleo aprisionadas em resina — reservatórios de carbono e lípidos acumulados a partir da fotossíntese da planta hospedeira, aguardando mobilização pelo fungo. Esta estrutura de armazenamento é central na simbiose arbuscular: as vesículas funcionam como depósitos de energia do fungo, acumulando triglicerídeos e outros lípidos que alimentam o crescimento da rede micelial externa invisível no solo. Um único hifa intraradical, não mais largo do que um pulso e transparente, filtra-se entre as vesículas na periferia inferior da câmara, o seu citoplasma internamente âmbar a prolongar-se para além das paredes celulares até à escuridão absoluta do exterior — o único fio que conecta este cofre repleto de lípidos a tudo o que existe do outro lado.
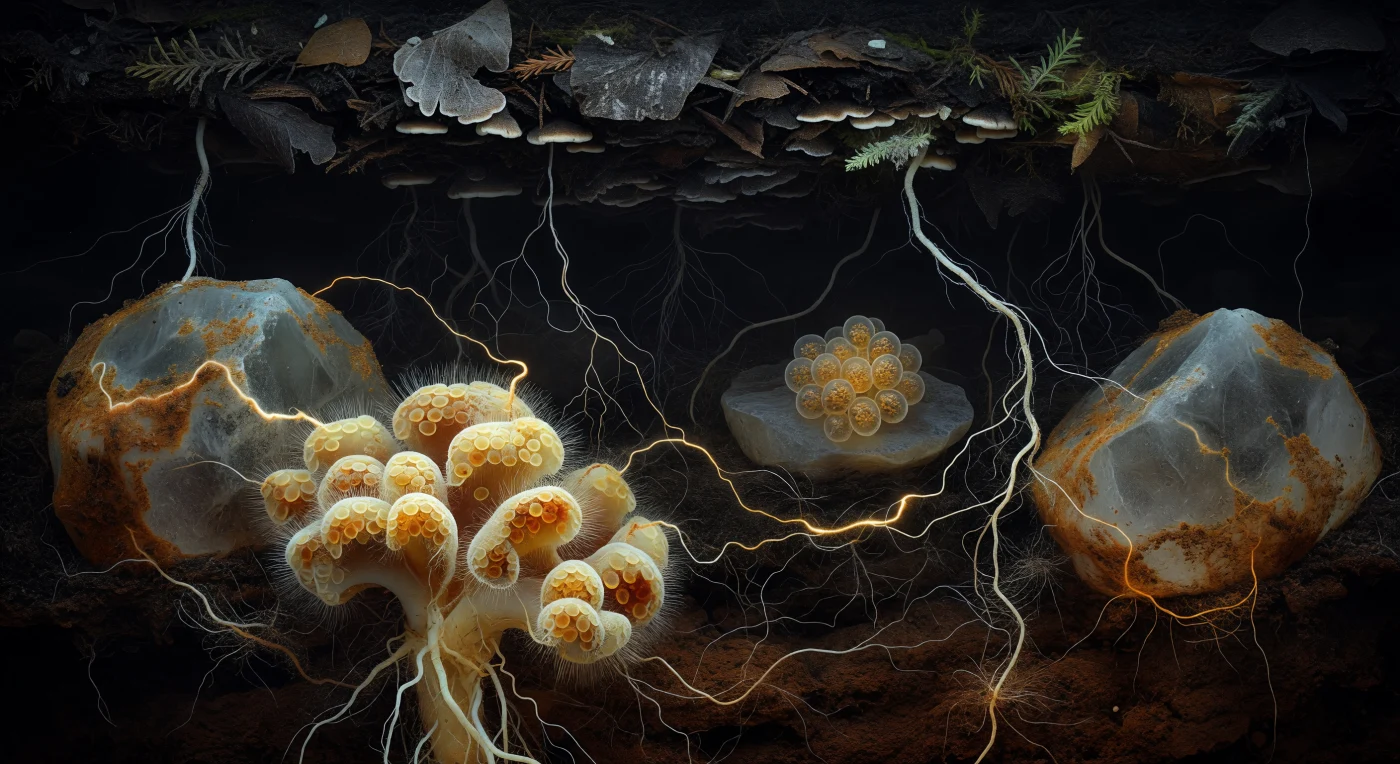
Suspenso na escuridão absoluta de um poro entre grãos de quartzo, o observador encontra-se imerso numa catedral subterrânea onde a luminosidade não é luz, mas química tornada visível — gradientes de carbono e fósforo que revelam, como numa imagem confocal onírica, uma das estruturas biológicas mais complexas do planeta. Em primeiro plano, um cluster ectomicorrízico domina a cena como uma formação coralínea barroca: o manto fúngico, com as suas camadas de creme, amarelo-manteiga e ocre-ferrugem de pseudoparênquima, envolve a ponta radicular numa fusão íntima onde o fungo e a raíz partilham trocas contínuas de açúcares fotossintéticos por fósforo e azoto mineral, uma simbiose mutuamente obrigatória que sustenta a maioria das árvores da Terra. A meia distância, a Rede Micorrízica Comum — o chamado *Wood Wide Web* — manifesta-se como uma malha tridimensional de fios brancos gossâmeros, cada hifa com 1 a 10 µm de diâmetro, tecendo ligações entre raízes de faia, pinheiro e carvalho separadas por dezenas de centímetros de solo mineral, alguns filamentos percorridos por uma luminescência âmbar ténue que trai o fluxo citoplasmático de fotossintatos em movimento. Um cacho de esporos repousa contra uma face de feldspato como ovos de vidro translúcido, cada um com 200 a 400 µm e paredes estratificadas que refratam a escassíssima luz em dourado profundo, enquanto acima, a transição do húmus quase negro para o solo mineral acastanhado-avermelhado compõe um horizonte que se perde na escuridão argilosa, engolindo os últimos fios da rede numa ausência total de luz.

Diante de você ergue-se a face cortada de um rizomorfo ectomicorrízico, exposta como a secção de um cabo enterrado há milénios — uma estrutura de 1,5 mm de diâmetro que, à escala em que pairamos, equivale à fachada de um edifício inteiro. O anel externo é uma armadura biológica de células melanizadas fundidas em algo semelhante ao vidro de obsidiana ou à casca carbonizada, os seus grânulos de melanina absorvendo cada fotão errante enquanto os cristais de quartzo do solo circundante se encostam ao rindo como paralelepípedos pressionados contra uma fundação antiga. A transição para a medula é abrupta e total: do lado de lá dessa fronteira, células-tubo pálidas e de paredes finas como papel de arroz molhado abrem-se em lumens vastos e fantasmagóricos, as suas vacúolos ocupando quase todo o interior celular como ovais cinzentos e translúcidos que difundem uma fosforescência fria para o escuro absoluto do solo. No eixo central, um canal oco perfura a estrutura inteira — a superfície interior ligeiramente húmida, com gotículas de água condensada a captar o brilho difuso do interior como um reflexo em túnel profundo. Para além do rindo, hifas-satélite finas como fibras ópticas partem para a matriz do solo comprimido, desaparecendo entre agregados de argila, películas orgânicas e grãos minerais numa química invisível à luz mas absoluta nas suas consequências — o transporte silencioso de carbono, fósforo e sinais químicos entre árvores que, à superfície, distam dezenas de metros.

À escala de um grão de poeira suspenso sobre o solo, o que se desdobra diante dos olhos é uma paisagem de canhões minerais e filamentos vivos que avança com uma urgência silenciosa e implacável: a frente micelial de uma micorriza arbuscular estende-se em leque a partir de uma hifa condutora central, cada ponta arredondada e densa de citoplasma irradiando uma luminescência azul-branca fria, como fibras óticas premidas através de obsidiana molhada. Os segmentos mais antigos por detrás da frente exibem grandes vacúolos translúcidos que se sucedem em intervalos regulares, conferindo às hifas o aspeto de cadeias de pérolas de vidro fosco, enquanto os grãos de quartzo que os rodeiam se erguem como monólitos de âmbar translúcido revestidos de filmes de óxido de ferro cor de ferrugem escura. Aqui e ali, grãos de calcite de superfície acremeada mostram halos de dissolução laranja-amarelados nos pontos de contacto hiphal — a secreção de ácido oxálico corrói lentamente a estrutura cristalina, deixando uma micro-topografia fosca e pejada de química quente contra o azul-frio dos filamentos vivos. As hifas navegam por entre os poros do solo com uma determinação que parece geológica na sua escala mas é inteiramente biológica na sua lógica: algumas espremem-se em câmaras de oito micrómetros de largura, deformando-se em secções ovais enquanto o citoplasma empurra para a frente; outras atravessam macroporos abertos como cabos de suspensão entre falésias. Ao fundo, mal discernível através das camadas de partículas e filmes orgânicos, uma parede curva cor de creme — a epiderme de um pelo radicular — organiza toda a paisagem numa só direção: cada ponta cega que avança na escuridão está orientada para aquele destino sem que nenhuma seta precise de ser desenhada.
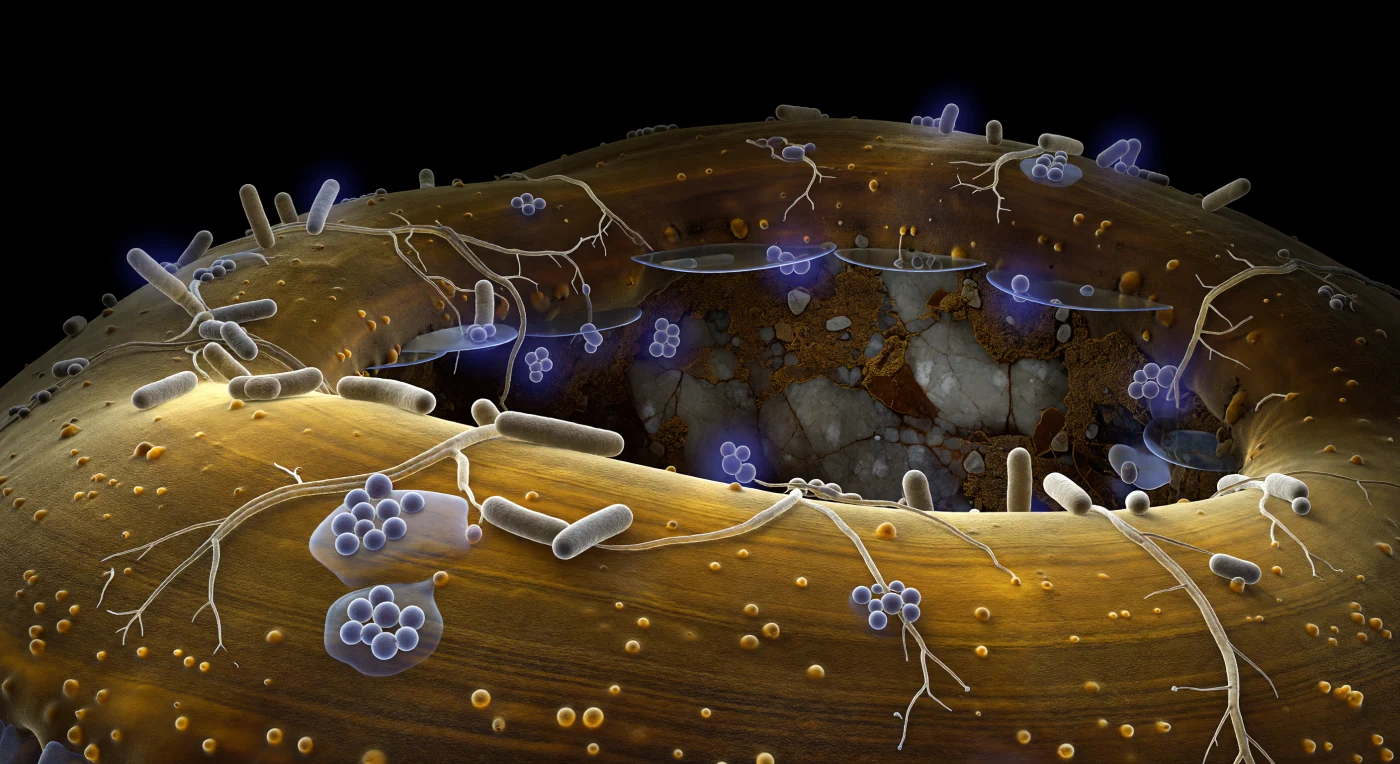
A superfície curva da hifa ectomicorrízica estende-se pelo campo de visão como o flanco de um penhasco de arenito envernizado, a parede quitinosa âmbar-dourada emitindo um brilho frio e translúcido a partir do citoplasma interior — a única iluminação neste subsolo sem luz é química, emanando de dentro como luz filtrada através de resina polida. A textura da parede revela bandas laminares de âmbar mais claro e mais escuro, salpicadas de protrusões glicoproteicas que emergem como bolhas arredondadas, e por toda esta topografia clivada se distribuem colónias de bactérias auxiliares da micorriza: bacilos Bacillus cinzento-bege, três vezes mais altos do que a escala humana imaginada aqui, alguns isolados e erectos, outros agrupados em pequenas colónias onde géis de exopolissacarídeo se acumulam entre as células como glicerina derramada, refractando o brilho ambiente em halos prismáticos subtis. Filamentos ramificados de Streptomyces traçam redes irregulares pela superfície da hifa, ancorados por ligações adesivas quase invisíveis que apenas se revelam como breves cintilações translúcidas onde a luz toca o ângulo de contacto certo, enquanto alguns indivíduos irradiam halos difusos de moléculas sinalizadoras em falsa cor violeta — gradientes esféricos que se esvanecem a poucos comprimentos bacterianos na película aquosa circundante, dando a esses organismos a aparência de lanternas fracas encostadas à parede âmbar. No fundo distante, uma superfície de feldspato fracturado dissolve-se em geometria angular mal iluminada, as suas películas orgânicas secas cor de caramelo escuro separadas da hifa por lençóis de água em menisco plano que reflectem o brilho citoplasmático através do poro do solo, neste mundo sem cima nem baixo, medido inteiramente em corpos vivos.

Você existe à escala de uma ponta hifal suspensa na escuridão fria e úmida de uma fissura de solo sob um prado temperado, e o que ocupa todo o seu campo visual é uma semente de orquídea *Dactylorhiza* — uma espécie de dirigível translúcido fundeado contra uma falésia de granito negro, o tegumento reduzido a uma única membrana iridescente tão fina quanto uma película de sabão, refratando a química imperceptível da água intersticial em pratas e verdes-azulados fantasmais. No interior dessa membrana, o embrião existe como um agrupamento frouxo de células quase esféricas e incolores, cada uma com uma luminosidade interna suave, a fosforescência metabólica de um organismo pausado na fronteira da possibilidade — porque as sementes de orquídea são radicalmente dependentes: desprovidas de endosperma, não germinam sem a invasão de um fungo do género *Rhizoctonia* que lhes forneça os açúcares e nutrientes necessários para o desenvolvimento inicial. A hifa que se aproxima pela esquerda lê-se, a esta escala, como um cilindro maciço de ouro-âmbar, a sua parede de quitina texturada como bambu laqueado, o citoplasma movendo-se em rios lentos de organelos — e onde já penetrou o tegumento, no interior de uma célula embrionária, o pelotão que formou preenche o espaço celular inteiro como uma mola de relógio enrolada sobre si mesma dezenas de vezes, estrutura densa de ouro e marfim pressionando a membrana da célula anfitriã até a deformar numa elipse perceptível, enquanto as células adjacentes já iniciam a diferenciação, o seu interior aquecendo de prata para um creme-dourado que assinala os primeiros passos moleculares da transformação — tudo isso acontecendo no silêncio químico e absoluto de uma terra negra de mineral húmido, onde o espaço recua em todas as direções como uma espessura tangível de frio e ausência total de luz.

Suspenso no interior de um poro do solo à escala de uma hifa — cerca de cinco micrómetros de altura —, o observador encontra-se numa catedral de escuridão geológica iluminada apenas pelo brilho âmbar da química da decomposição: grãos de quartzo erguem-se como arranha-céus revestidos por uma película de glomalina translúcida, cor de mel cru, e da cortex radicular desintegrada acima tombam em câmara lenta esporos de fungos micorrízicos arbusculares — corpos esféricos e elipsoidais do tamanho de rochas e pequenas casas à esta escala, com paredes espessas que irradiam tons âmbar, vermelho-vinho e ocre-amarelo. Estendendo-se horizontalmente por todo o campo de visão, os filamentos remanescentes da rede miceliana comum percorrem a escuridão como cabos prateados de diâmetros variáveis, transportando ainda um fluxo citoplasmático visível que vai depressurizando à medida que a árvore morta se dissolve, enquanto o micélio saprotrófico coloniza as ruínas celulares com hifas de quase o dobro do diâmetro, branco-creme e opacas, agarrando o substrato em decomposição com urgência anastomótica. Da esquerda, uma frente micorrízica amarelo-pálida avança como uma maré de fio de ouro sobre a turfa escura; da direita, uma rede creme-branca de maior densidade já contacta os primeiros esporos libertados, enquanto entre ambas as redes convergentes o solo se apresenta como uma paisagem de agregados húmicos laqueados em âmbar e meniscos de água que captam toda a luz disponível em superfícies de espelho curvas. Este instante congela uma catástrofe silenciosa de consequências biológicas imensuráveis: a morte de uma árvore não é um fim, mas uma redistribuição feroz de capital nutricional por entre os sobreviventes da floresta.
