Confianza científica: Alto

En lo más profundo del suelo, avanzas por un laberinto de piedra y arcilla que ninguna luz solar ha tocado jamás: las paredes del mesoporo están tapizadas de laminillas de esmectita color plata apiladas como hojas de cuchillo fino, entre las que emergen bóvedas traslúcidas de cuarzo que refractan la única fuente de claridad disponible —el resplandor biológico frío y verdoso de la hifa arbuscular micorrícica que serpentea ante ti, ocupando casi un cuarto del corredor como un tubo de vidrio borosilicato vivo. A través de su pared inmaculada y tensa se adivinan los gránulos del citoplasma —lípidos crema y ámbar derivando en una marea imperceptible— mientras finas películas de agua la anclan a las platelets de arcilla en meniscos curvos que capturan y doblan la escasa luminosidad en arcos brillantes. Las biopelículas bacterianas cubren los rincones minerales como niebla de cilindros grises, cada uno apenas un quinto del diámetro de la hifa, recordándonos que este laberinto es una metrópolis química donde azúcares carbono fluyen desde árboles lejanos y el fósforo es negociado en segundos a través de membranas que ahora podrías casi rozar con la mano. Más adelante, tres túneles ramificados se abren hacia una oscuridad absoluta e impenetrable, sus bocas enmarcadas por costras orgánicas de ámbar oscuro, y la sensación es la de encontrarse ante los umbrales de galerías submarinas sin nombre, comprimidas hasta la anchura de una célula humana.
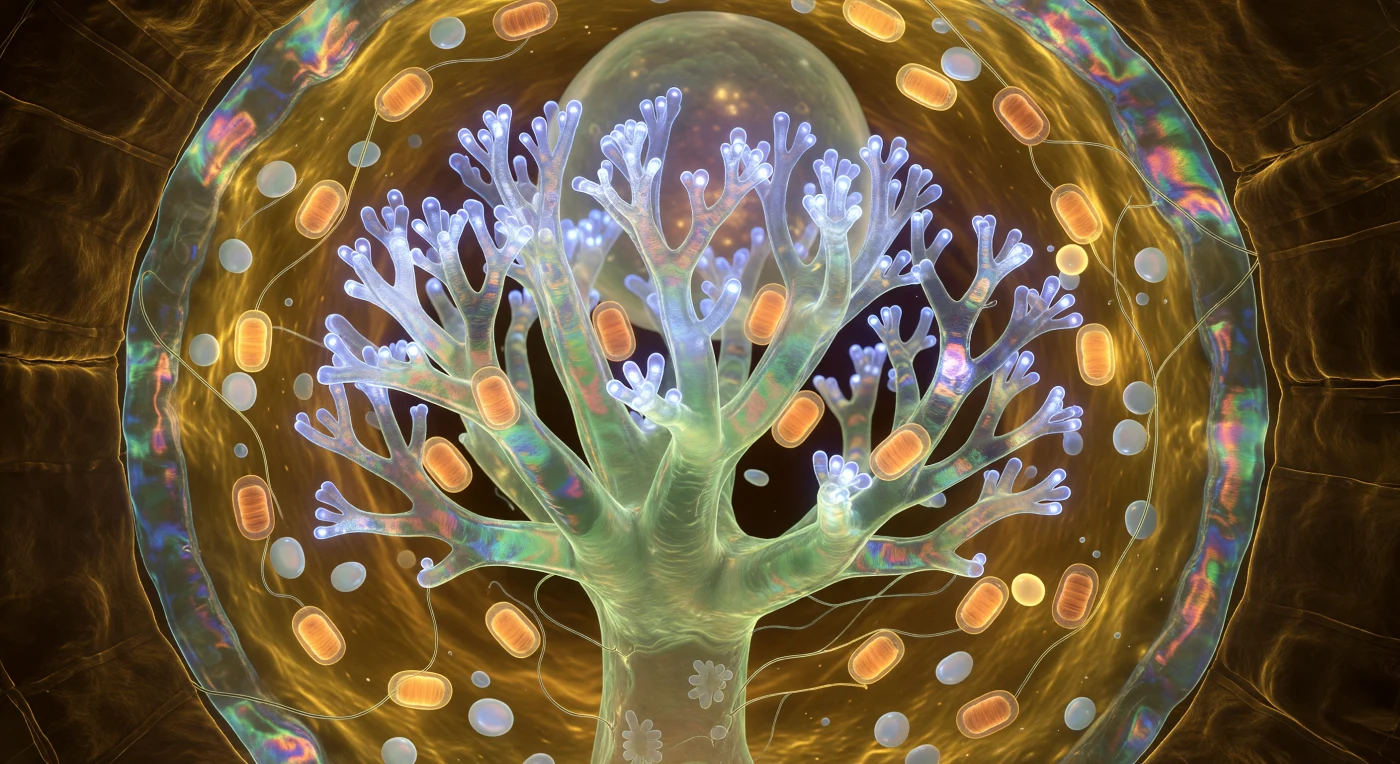
Flotas suspendido en el interior de una catedral que nunca fue construida sino cultivada: ante ti se eleva el arbúsculo como un bosque de coral invertido congelado en plena floración, su tronco principal una columna de jade traslúcido que se ramifica en dicotomías cada vez más finas hasta que las ramificaciones terminales, de apenas medio micrómetro de ancho, se disuelven en una neblina trémula de luminiscencia cobalto-blanca generada por miles de proteínas transportadoras de fosfato ciclando en conformación sincronizada, su actividad química convertida en luz fría que asciende en ondas lentas desde la base hasta las puntas como bioluminiscencia remontando un arrecife nocturno. Toda esta arquitectura ramificada está envuelta en la membrana periarbuscular, una doble piel que iridisce del ámbar intenso al verde azulado y al rosa profundo según su grosor, sin ninguna fuente de luz exterior —no hay fotón del mundo macroscópico que penetre esta profundidad de suelo— y sin embargo ves todo por el fuego metabólico frío del intercambio mismo. Alrededor del tronco arbuscular, mitocondrias en forma de cigarro de dos a tres micrómetros de largo flotan como faroles ámbar en el citoplasma viscoelástico dorado, sus crestas internas visibles como anillos concéntricos a través de membranas translúcidas, sus halos de respiración quimiosmótica superponiéndose en el gel circundante mientras amiloplastos perlados derivan como cantos opalescentes refractando el azul bioluminiscente en destellos blancos y violetas pálidos. La pared celular curva todo el escenario como una muralla de ámbar oscuro profundamente texturada, sus microfibrillas de celulosa en hélices cruzadas captando el resplandor arbuscular en ángulos rasantes para revelar una topografía de crestas y depresiones como el interior de una bóveda tallada; en el intersticio apoplástico de apenas cien nanómetros entre las ramas exteriores del arbúsculo y la membrana plasmática huésped ocurre en este preciso instante el intercambio de fosfato y azúcar que sostiene, desde esta única célula encendida, un bosque de kilómetros de ancho.

En este paisaje de escala milimétrica, te encuentras de pie en una vasta llanura arenosa que se extiende como una meseta desértica al atardecer, donde la luz difusa se filtra desde arriba a través de los estratos minerales como si atravesara grueso vidrio ámbar. Los protagonistas del primer plano son las esporas de hongos micorrícicos arbusculares —Glomus, Gigaspora y Scutellospora—, estructuras reproductivas de resistencia que pueden permanecer viables en el suelo durante décadas, y que aquí aparecen como enormes globos lacados de ámbar profundo, crema rosado y vino tinto, cada una con una arquitectura de pared multilaminada que en microscopía de luz revela capas concéntricas de quitina y glucanos con distinta densidad óptica. Entre ellas se tienden filamentos de hifas casi invisibles —paredes de apenas 1–5 µm de espesor, transparentes como vidrio soplado— que conectan las esporas a través de los huecos abiertos entre granos de cuarzo translúcido y feldespato rosado del tamaño de edificios, siguiendo curvas catenarias dictadas por la tensión superficial de las películas de agua que cubren los poros. Fragmentos de materia orgánica en descomposición, oscuros y planos como adoquines de resina petrificada, alfombran el suelo arenoso entre los granos minerales, representando la fuente principal de carbono y nutrientes que esta red subterránea movilizará hacia las raíces de las plantas. Todo el conjunto transmite la paradoja fundamental del mundo edáfico: una arquitectura biológica de extraordinaria delicadeza —hilos vivos más finos que un cabello humano— sostenida dentro de un paisaje mineral de peso geológico aparentemente eterno.
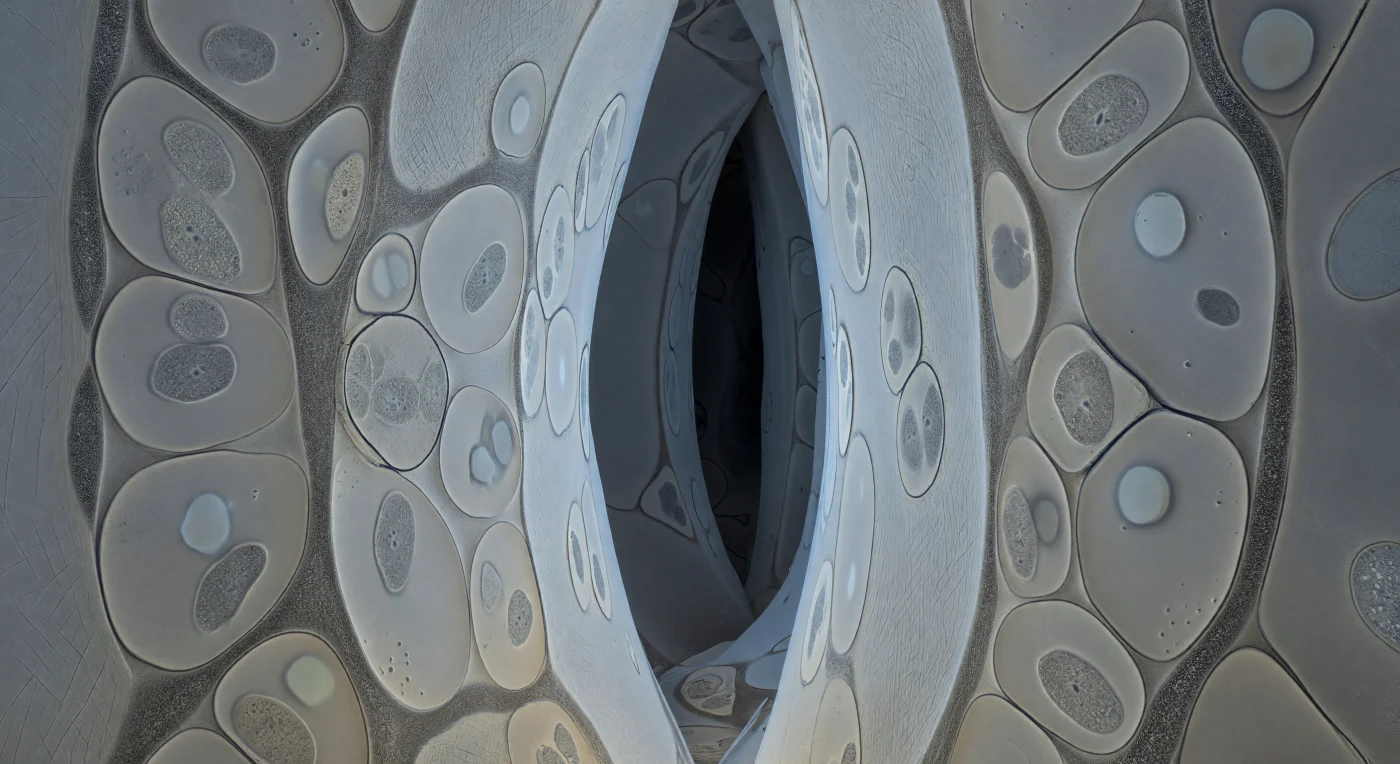
Te encuentras atrapado en el interior de un corredor intercelular tan estrecho que las paredes de celulosa te rozan por todos lados: vastas superficies de marfil pálido se elevan en láminas suavemente onduladas, sus microfibrillas dispuestas en diagonales cruzadas como el grano de un papel antiquísimo comprimido bajo presiones geológicas. Las secciones transversales de las hifas fúngicas llenan cada espacio disponible como lentes oblatas prensadas contra las células vegetales, sus membranas plasmáticas trazadas en grafito oscuro con una tensión que casi se percibe como resistencia física, mientras el citoplasma interior alberga mitocondrias punteadas y vacuolas que brillan como ausencias ópticas en la penumbra. Entre la pared fúngica y la pared celular vegetal corre una franja granular de apenas 15 a 25 nanómetros —la matriz interfacial de glicoproteínas— que en este instante congelado aparece como una textura tibia y heterogénea, ni membrana ni vacío, sino el umbral negociado donde el fosfato abandona el citoplasma fúngico y la sacarosa entra desde la planta. Más allá de esta cámara inmediata, los corredores se pierden en ranuras de pizarra profunda, la arquitectura hexagonal pero orgánica e irregular, un laberinto sin luz exterior cuya dimensionalidad emerge únicamente del contraste estructural extremo: cada interfaz proyecta su propia sombra de borde, y eso es suficiente para que el espacio exista.
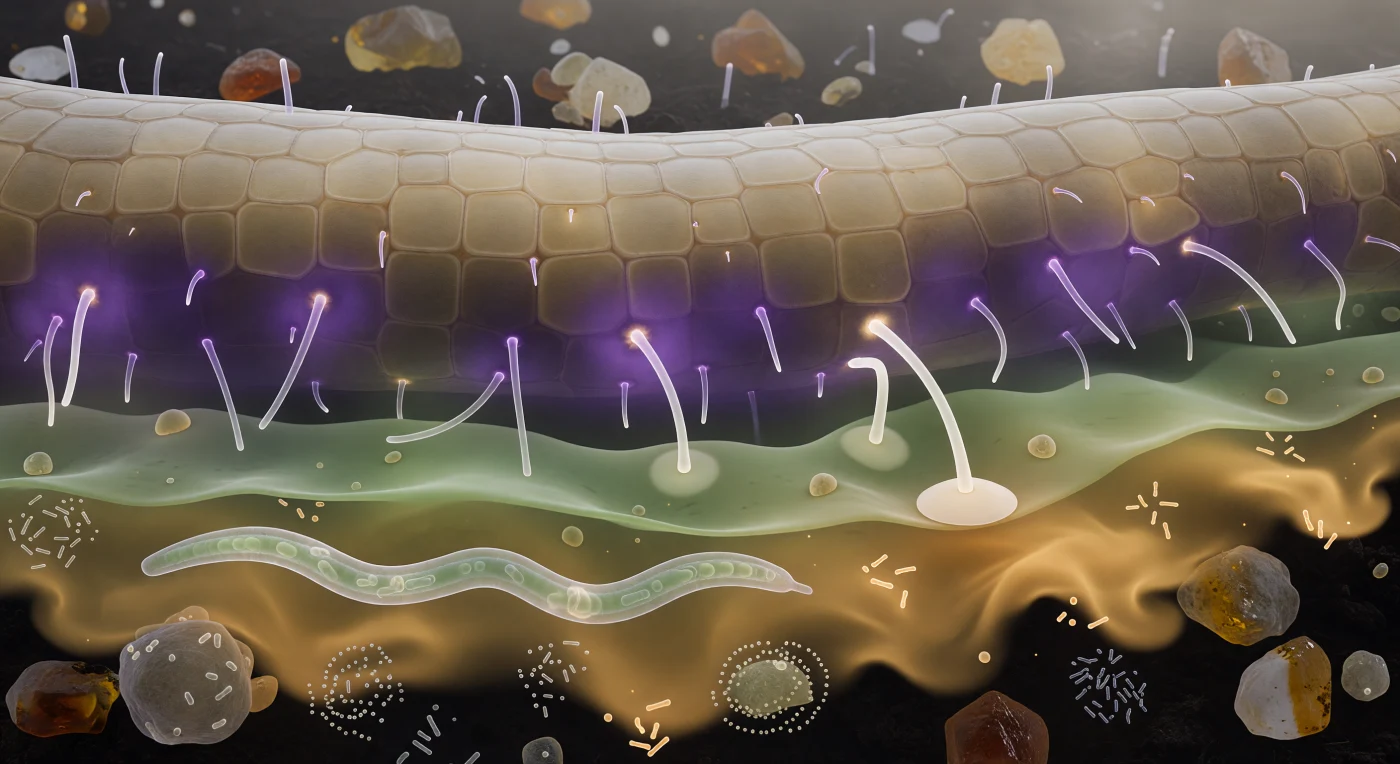
Suspendido frente a la curva de la raíz, el espectador contempla una pared de arenisca pálida que se arquea hacia el horizonte como el flanco de un acantilado, su epidermis compuesta de losas translúcidas color crema separadas por crestas finas como mortero antiguo, mientras pelos radicales de cristal puro proyectan su luminiscencia espectral hacia el espacio intersticial. Entre el observador y esa superficie lo invisible se ha vuelto materia: una corona violeta densa de estrigolactonas y flavonoides abraza los tejidos epidérmicos como niebla termal, de ella emerge hacia afuera un gradiente verde-celadón de aminoácidos que sangra como acuarela en papel húmedo, y más allá una columna ámbar-dorada de exudados azucarados se deshilacha lentamente hacia la oscuridad del suelo bulk con la cadencia de una corriente bioluminiscente. En el tercio inferior del campo visual un nematodo sinusoidal atraviesa la escena como un filamento de cuarzo animado, sus órganos internos apenas visibles a través de la cutícula iridiscente, rodeado de enjambres bacterianos que pulsan en espirales sobre las caras de los agregados minerales. Dos puntas hifales de marfil doblan su trayectoria con precisión quimiotrópica a través de las capas coloreadas, siguiendo el gradiente violeta como navíos que bordean una costa, y una ya ha llegado: su extremo aplanado en disco de apresorio descansa exactamente sobre una baldosa epidérmica convexa, perfectamente adherido, comenzando en silencio enzimático su entrada en la pared que tiene debajo.

La superficie sobre la que te encuentras de pie es un tejido vivo: una plecténquima apretada de células fúngicas alargadas, dispuestas como adoquines cerámicos pulidos a mano, cada una de entre cinco y quince micrómetros de anchura, con paredes ligeramente translúcidas de color crema en su núcleo y bordes teñidos de amarillo azufre pálido donde las membranas adyacentes se presionan entre sí. Esta envoltura protectora —el manto ectomicorrícico de *Suillus*— forma una capa continua y ceñida alrededor de la punta radicular, una interfaz biológica donde el hongo y la raíz negocian el intercambio de carbono fotosintético por fósforo y agua minerales, todo ello a través de la red del Hartig que se infiltra invisiblemente entre las células epidérmicas de la raíz, apenas perceptible como un leve resplandor ambarino que asciende desde dentro del tejido como luz de lámpara tras papel de arroz. Detrás de ti se alza la pared curva e inmensa de la propia raíz —tan suave y tan vasta como el casco de un barco varado, su curvatura disolviéndose en la penumbra antes de completar su arco—, mientras que fragmentos angulosos de cuarzo del tamaño de bloques y láminas de arcilla se acumulan como sedimento en las grietas del manto, recubiertos de una película orgánica de color ámbar oscuro. Hacia el borde deshilachado del manto, docenas de hifas extraradicales de una sola célula de grosor se lanzan hacia la oscuridad absoluta del suelo no colonizado como filamentos de fibra óptica: tubos transparentes y refractivos de tres a seis micrómetros de diámetro, con el citoplasma barely visible en su interior, cada uno rodeado de un vacío tan negro y tan denso que parece materia sólida, y cuyo avance lento e incesante —guiado por señales químicas imperceptibles— constituye la vanguardia viva de una red que puede extenderse durante metros a través del bosque.
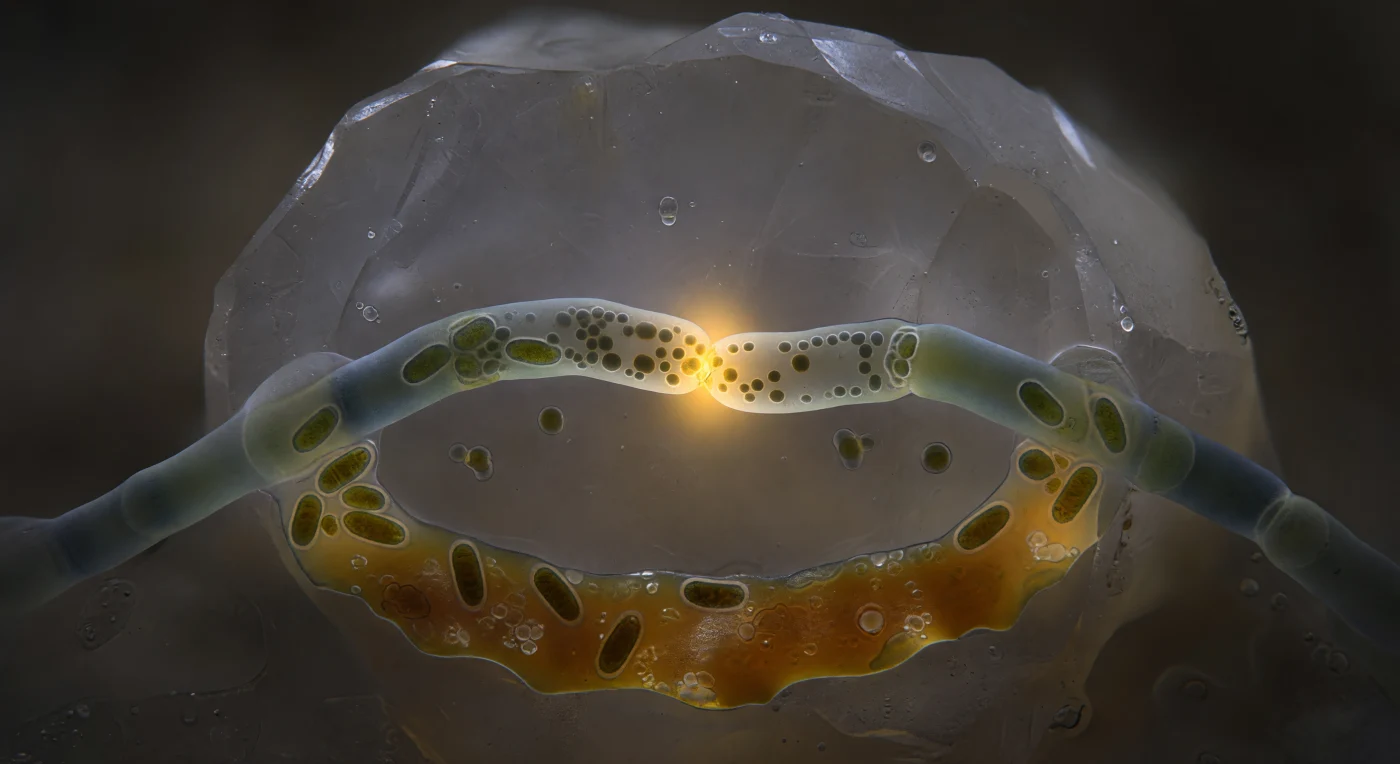
En el interior de un poro de suelo completamente oscuro, dos hifas de hongos micorrícicos arbusculares se curvan la una hacia la otra como cables de suspensión guiados por una tensión química invisible, sus paredes de quitina-glucano translúcidas brillando con un tenue tono de vidrio marino en el único resplandor disponible: la luz ámbar-dorada que emana del punto de fusión mismo. En esa zona de contacto, apenas un disco de un micrómetro y medio de diámetro, las paredes celulares se han disuelto entre sí, borrando la frontera entre dos organismos distintos en un proceso llamado anastomosis hifal, mediante el cual las redes fúngicas se interconectan para redistribuir nutrientes, señales y orgánulos a través de la miceliosa colonia. Las mitocondrias se agolpan en el margen del poro como cuerpos de color ámbar cobrizo, sus membranas internas apenas visibles como estrías más oscuras, mientras gránulos lipídicos y citoplasma fluyen en una corriente viscosa desde la hifa izquierda hacia la derecha, una transferencia de materia viva en tiempo real. Detrás de este punto de confluencia, el grano de feldespato asciende como un acantilado pulido por un glaciar, difuminándose hacia un gris plateado en los bordes del campo de visión, y la película de agua que recubre cada superficie dispersa el resplandor ámbar en un suave halo que convierte este poro de cuarenta micrómetros en algo que se siente, a la vez, íntimo e inconmensurablemente vasto.

En esta imagen, un único macroagregado de suelo de tres milímetros ocupa el encuadre como si fuera un mundo completo: su superficie de materia orgánica humificada y restos fúngicos impregnados de melanina forma un paisaje oscuro salpicado de inclusiones minerales —granos de cuarzo y feldespato— que emergen como témpanos pálidos de una matriz casi negra. Una película de glomalina, esa glicoproteína secretada por hongos micorrícicos arbusculares, recubre cada cara expuesta del agregado con un barniz ámbaro-dorado que le confiere una ligera hidrofobicidad, evidente en la gota de agua que reposa sobre la superficie con un ángulo de contacto pronunciado, reflejando el mundo oscuro en su interior como un espejo convexo. Desde las fisuras ecuatoriales del agregado brotan hilos hifales blancos en arcos paralelos, finos como hilo de seda, que actúan literalmente como suturas biológicas: son las estructuras que cosen y estabilizan el agregado, atrapando carbono orgánico en una arquitectura que puede persistir décadas. En el ángulo inferior derecho, la iluminación ultravioleta transforma la escena por completo: la oscuridad ambiente se profundiza hasta parecer espacio exterior y la glomalina estalla en una fluorescencia amarillo-verdosa incandescente que traza con precisión cada cresta y grieta del agregado, revelando la geografía invisible de un suelo que los hongos llevan construyendo cuatrocientos millones de años.
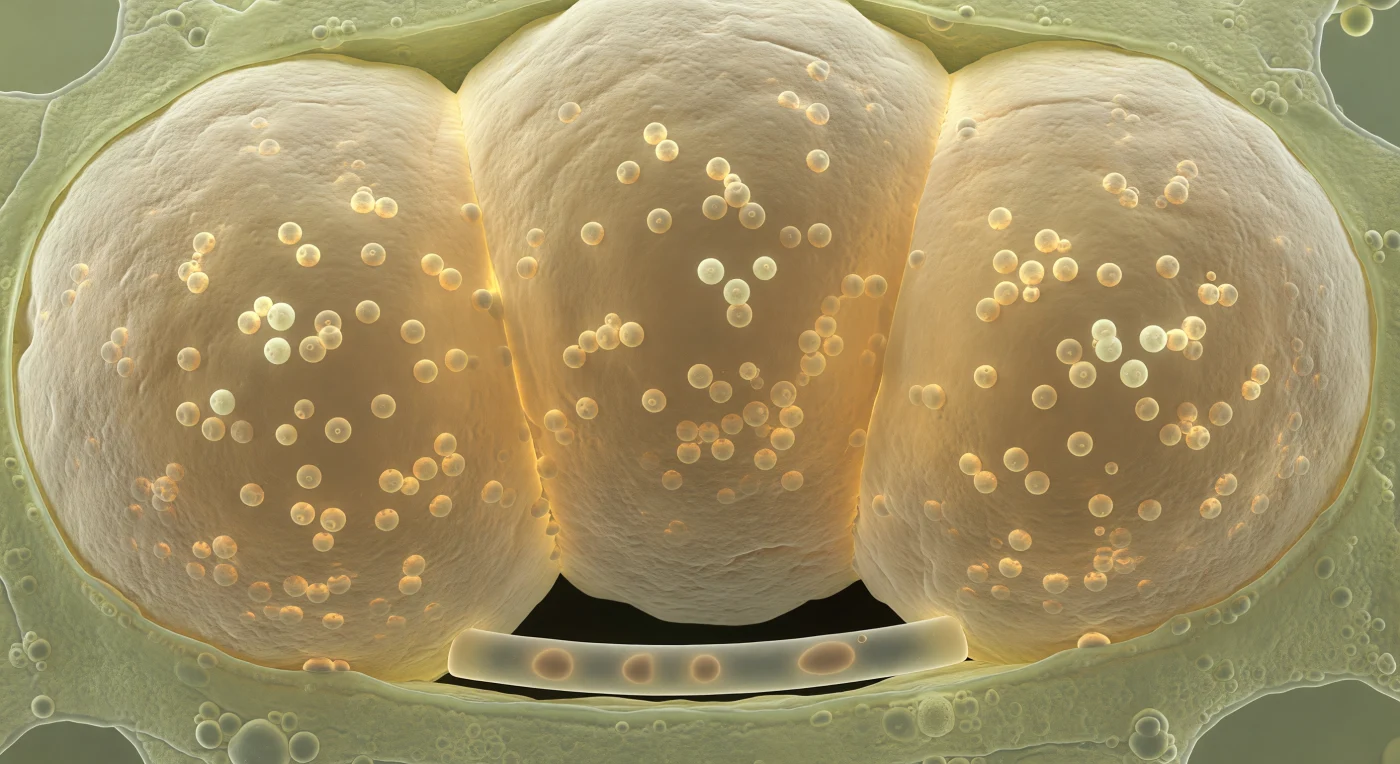
Te encuentras suspendido en el interior de una célula cortical de raíz, aprisionado entre los flancos de tres vesículas colosales que llenan el volumen como globos de ámbar pálido inflados hasta el límite: estas estructuras son vesículas arbusculares micorrícicas, órganos de almacenamiento formados por el hongo simbionte *Rhizophagus irregularis* dentro de los tejidos de la planta hospedadora, cuyas paredes laminadas de quitina y glucano —rugosas y estriadas en la superficie exterior, lisas y vítricas hacia el interior— encierran decenas de glóbulos lipídicos de cinco a diez micrómetros de diámetro que flotan suspendidos en un medio viscoso y translúcido, refractando la luz ambiental en destellos dorados como gotas de aceite atrapadas en resina. Allí donde las tres vesículas se presionan mutuamente y contra la pared celular, la geometría cede en zonas de contacto aplanadas, y el citoplasma verde pálido de la célula hospedadora ha sido comprimido hasta los márgenes como cera líquida, reducido a una película granular y translúcida cargada de ribosomas y orgánulos. En el límite inferior de tu campo visual, casi imperceptible, una sola hifa intraradical de cinco micrómetros de anchura —lisa, incolora, cubierta por una fina película de agua— serpentea entre dos de las vesículas y desaparece en la oscuridad absoluta entre las paredes celulares, único filamento que conecta este depósito cargado de lípidos con la red exterior invisible. No hay fuente de luz externa: toda la iluminación proviene del interior, la autofluorescencia difusa de las membranas comprimidas y las interfaces lipídicas que irradian como si la química de este espacio fuera cálida.
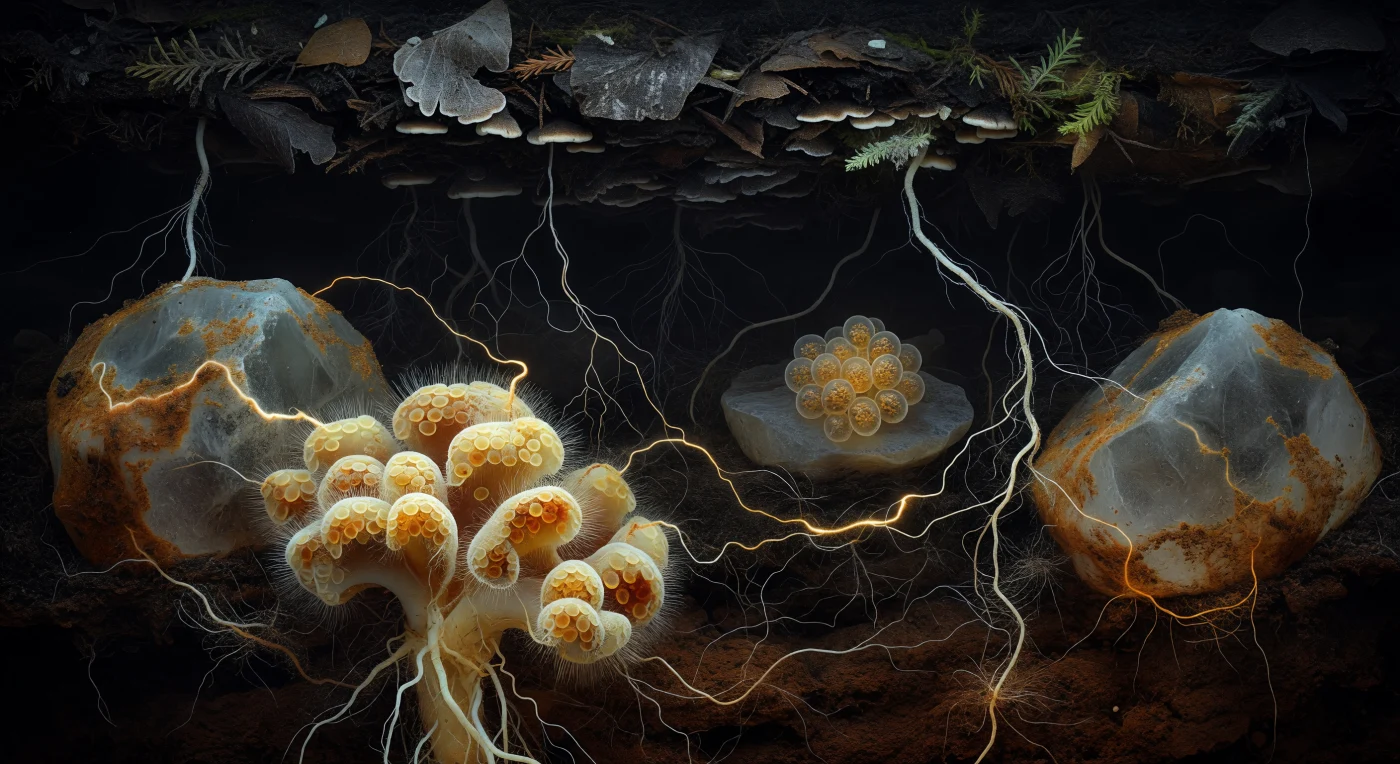
Te encuentras suspendido en la oscuridad absoluta de un poro del suelo antiguo, a la deriva entre dos masas minerales de cuarzo cuyas caras están cubiertas por una pátina ámbar de materia orgánica que parece guardar calor propio como resina vieja. Lo que ves no es luz en ningún sentido convencional, sino una representación de gradientes químicos y flujos biológicos hecha visible: directamente ante ti, un racimo de puntas radiculares ectomicorrícicas domina el primer plano como una formación coralina barroca, con su manto pseudoparenquimatoso en capas de crema, amarillo mantequilla y óxido ocre, cada faceta celular reluciendo levemente con su película de humedad, mientras hifas extraradicales se irradian hacia afuera como cabellos en una corriente imperceptible. Más allá, la Red Micorrícica Común —la llamada Wood Wide Web— se revela como la textura definitoria de este mundo subterráneo: hilos blancos traslúcidos, cada uno apenas tan grueso como una línea trazada con pluma fina, tejen una malla tridimensional que conecta raíces de picea, haya y roble a través de decenas de centímetros de oscuridad mineral, algunos filamentos irradiando un cálido resplandor ámbar que delata el flujo citoplásmico de azúcares fotosintéticos viajando de árbol en árbol. En el plano medio, un racimo de esporas translúcidas reposa contra una cara de feldespato como huevos de vidrio marino, sus paredes estratificadas encierran glóbulos lipídicos que refractan la escasa luminosidad en tonos de oro y ámbar profundo, mientras el gradiente del horizonte A —desde el humus casi negro de la superficie hasta el suelo mineral rojizo-marrón de las profundidades— absorbe progresivamente la red, los hilos se vuelven más tenues y finalmente desaparecen en la arcilla sin luz del fondo.

En la oscuridad absoluta del suelo, lo que se despliega ante ti es la cara expuesta de un rizomorfo ectomicorrícico cortado en sección transversal, un cilindro de apenas millón y medio de micrómetros de diámetro que, a esta distancia, se alza como el pilar fundacional de una ciudad enterrada. La corteza exterior forma un anillo de células melanizadas tan densamente empaquetadas que se han fundido en algo parecido a corteza carbonizada u obsidiana, su melanina granulosa visible dentro de las paredes engrosadas como una armadura biológica capaz de atrapar y extinguir cualquier fotón errante; inmediatamente después de esa frontera nítida, la médula se abre en una catedral de células-tubo pálidas, de paredes delgadas como papel de arroz húmedo, cuyos lúmenes amplios y vacuolas translúcidas emiten una fosforescencia fría, azul-blanca, que se difunde hacia afuera como una química invisible haciéndose visible. En el eje más profundo del rizomorfo se abre un canal hueco, un túnel circular cuya superficie interior ligeramente húmeda capta ese resplandor interno como un brillo acuoso a lo largo de su curvatura, mientras las células medulares circundantes se disponen como las cámaras de un nautilo visto desde arriba, algunas con sus membranas plegadas hacia adentro en suaves acordeones. Fuera del perímetro externo, hifas satélite de apenas unos pocos micrómetros de anchura se irradian hacia la matriz del suelo comprimido, filamentos casi vítreos que se pierden entre granos de cuarzo presionados contra el rizomorfo como adoquines mortados contra un cable enterrado, sus facetas angulares captando la luz difusa como ladrillos de vidrio esmerilado, la estructura entera legible como un cable biológico antiguo y preciso, diseñado para transportar carbono, agua y fósforo a través de una red que, a escalas mayores, sostiene bosques enteros.

En el horizonte de este mundo comprimido, una franja de puntas hifales avanza como un delta fluvial vivo, cada filamento irradiando una luminiscencia azul-blanca espectral que perfora la oscuridad absoluta del suelo mineral. Las puntas más jóvenes son cilindros opacos y densos, ciegos como gusanos que presionan hacia adelante, mientras que los segmentos más maduros detrás de ellas muestran vacuolas claras que les dan el aspecto de collares de perlas nubladas, sus vientres ligeramente amarillentos donde se concentran reservas lipídicas. Estas hifas de hongos micorrícicos arbusculares —estructuras vivas de entre uno y diez micrómetros de diámetro— secretan activamente ácido oxálico en contacto con granos de calcita, disolviendo la superficie mineral en una aureola de química cálida color azufre que contrasta con el frío biológico de la luminiscencia fúngica. Donde los filamentos atraviesan poros angostos entre monolitos de cuarzo color ámbar oxidado, el citoplasma se deforma levemente en sección oval, como si la vida misma se exprumiera hacia adelante con urgencia. En el fondo apenas legible a través de la turbiedad orgánica, la pared curva de un pelo radical —enorme como un acantilado de celulosa— organiza silenciosamente toda la dirección del paisaje, destino invisible que orienta cada punta en crecimiento con la precisión de una brújula bioquímica.
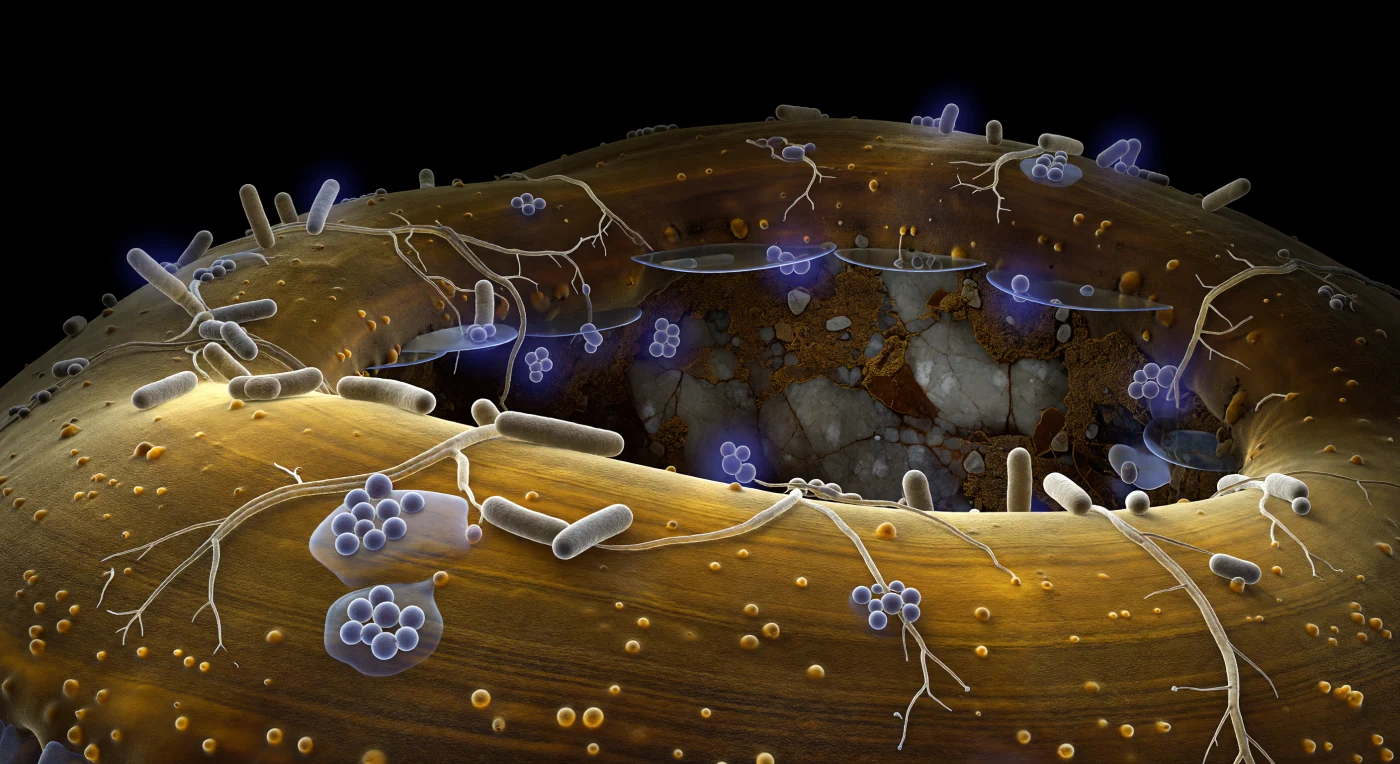
La superficie que se extiende ante el observador es la pared de una hifa ectomicorrícica, una arquitectura de quitina y glucoproteínas que a esta distancia se curva como el flanco de un acantilado de arenisca vidriada, su ámbar cálido iluminado desde adentro por la dispersión fría del citoplasma subyacente, como luz filtrada a través de carey pulido. Adheridas a esta topografía granular, bacterias Bacillus en forma de cilindros gris-beige se distribuyen aisladas o agrupadas en pequeñas colonias donde el gel de exopolisacáridos se acumula entre ellas como glicerina derramada, refractando el resplandor ambiental en halos prismáticos apenas perceptibles; filamentos ramificados de Streptomyces trazan redes irregulares sobre la superficie hiphal, anclados por tenues amarras adhesivas que solo se revelan como destellos translúcidos cuando el ángulo de luz los roza. Algunos individuos bacterianos emiten halos violetas en falso color —moléculas señalizadoras difundiéndose en gradientes esféricos a través de la película acuosa que rodea la hifa— convirtiendo a esas células en pequeñas linternas apagadas pegadas contra el acantilado de ámbar. En el fondo distante, la pared de un grano mineral de feldespato fragmentado emerge apenas desde la oscuridad absoluta del poro de suelo, su superficie gris-blanca con destellos micáceos y recubierta de materia orgánica seca color toffee oscuro, mientras finas láminas de agua en menisco tienden puentes invisibles entre la hifa y esa pared remota, reflejando el azul frío del citoplasma a través del vacío subterráneo permanente.

En la oscuridad absoluta de una grieta de suelo bajo un prado templado, una semilla de orquídea *Dactylorhiza* de apenas 300 micrómetros flota como un dirigible translúcido contra paredes de granito negro empapado: su cubierta seminal, una membrana de una sola célula de grosor, refracta la química bioluminiscente del agua intersticial en plata fría y verde-azulado tenue, mientras que a través de ella el embrión se revela como un puñado de células esféricas sin color, cada una con el brillo apagado de una linterna de papel encendida muy despacio. Desde la izquierda inferior avanza una hifa de *Rhizoctonia*, un cilindro dorado-ocre cuya pared de quitina texturada como bambú lacado resulta inmensa a esta escala, y ya ha penetrado la cubierta seminal formando dentro de una célula embrionaria un pelotón —un resorte de relojería de hifas enrolladas sobre sí mismas decenas de veces, ámbar y marfil pálido, que empuja la pared celular hasta deformarla en una elipse perceptible. Las células adyacentes al pelotón han comenzado ya a diferenciarse: sus membranas se tensan y su luminosidad interior vira del blanco frío hacia un crema dorado cálido, los primeros pasos moleculares de un organismo que hasta hace instantes permanecía detenido en el umbral de la posibilidad. Todo lo demás es materia oscura: granos angulosos de basalto y cuarzo cubiertos de películas húmicas color caramelo, meniscos de agua perfectamente quietos curvándose bajo la tensión superficial, y más allá, una negrura tan densa que se percibe como una presencia física, un frío mineral que se extiende en todas direcciones interrumpido apenas por colonias de bacterias —cilindros pálidos pegados a superficies de cuarzo como percebes en un acantilado— increíblemente pequeñas incluso a esta escala.

Suspendido en la oscuridad absoluta de un poro del suelo, el observador se encuentra rodeado por torres de cuarzo recubiertas de una resina ámbar translúcida que difunde el único resplandor disponible: la química lenta y fría de la descomposición. Sobre esta escena se abre la arquitectura derrumbada de la corteza radicular del árbol muerto, sus paredes celulares disgregándose en cintas fibrosas desde las que caen en cámara lenta esporas de hongos micorrícicos arbusculares —esferas y elipsoides del tamaño de peñascos o pequeñas casas a esta escala, de paredes gruesas que brillan en ámbar, rojo vino y ocre, algunas ya resquebrajadas y rezumando fluido lipídico hacia la película de agua que todo lo recubre. A media distancia, los cables plateados de la red miceliar común se extienden hacia los horizontes minerales en ambas direcciones, portando todavía una translucidez interna que delata el flujo citoplasmático residual hacia los árboles vecinos que siguen vivos; mientras tanto, un micelio saprótrofo de hifas blancas y opacas —casi el doble de gruesas que los filamentos micorrícicos— coloniza las ruinas celulares con una urgencia arquitectónica y sin ambages. Desde la izquierda avanza un frente de hifas de un amarillo pálido como un maremoto de hilo dorado, y desde la derecha una red crema más densa converge sobre las esporas liberadas, los dos frentes aproximándose lentamente en esta zona de máxima riqueza de nutrientes donde la muerte de un gigante se convierte, en silencio y en tinieblas, en el sustento de los que permanecen.
