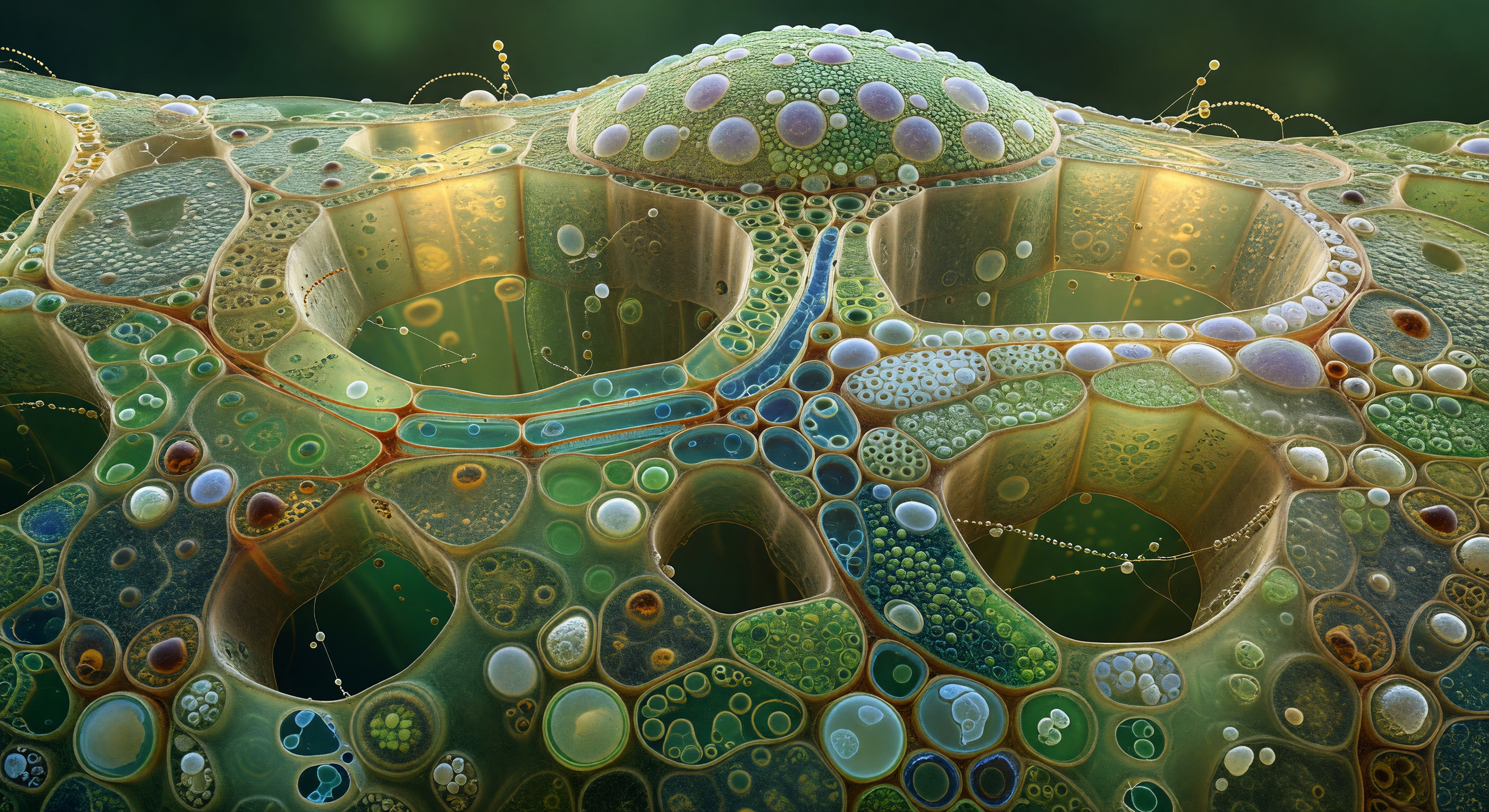
Te encuentras en el interior de un corredor lumenal de 120 micrómetros de diámetro —la cavidad de un vaso de xilema maduro de angiosperma—, una galería de luz ámbar-siena que se extiende hacia el horizonte como la nave de una catedral excavada en resina compacta y madera densa, sus paredes tapizadas por cientos de puntuaciones areoladas dispuestas en filas escalonadas, cada una una depresión circular de seis micrómetros coronada por un labio anular de celulosa lignificada que captura el resplandor difuso, con una membrana fantasma nacarada y casi iridiscente suspendida sobre su apertura, vestigio del gradiente de presión que alguna vez reguló el flujo entre vasos vecinos. El vaso está completamente vacío y bajo tensión hidráulica negativa: no circula agua aquí, sino que la columna cohesiva asciende en algún otro punto hacia la luz del dosel, y el silencio seco y absoluto que resulta es la firma física de ese esfuerzo invisible. Al fondo del corredor, donde la perspectiva comprime el patrón de círculos repetidos en un papel pintado puntillista que se disuelve en la penumbra, una burbuja de embolia aérea ocupa el lumen por completo —una lente de mercurio convexo que refleja toda la galería en miniatura distorsionada, sus bordes anclados a la pared lignificada por un ángulo de contacto preciso, inmóvil en el umbral entre cohesión y cavitación. Es un instante congelado de colapso hidráulico: el vaso ha dejado de conducir, y la burbuja, bella como una grieta en un cristal, es su silenciosa sentencia de muerte.

De pie en el suelo de la hoja, los pies sobre una losa translúcida de pared celular impregnada de cera, la mirada sube a lo largo de columnas cilíndricas de setenta micrómetros de altura que se alzan como las naves de una catedral de cristal verde: las paredes de cada columna, ámbar-verdosas y semiopacas, están empapeladas por dentro con centenares de cloroplastos biconvexos que se ajustan entre sí como teselas de esmeralda, cada uno irradiando una fría luminiscencia propia que suma sus penumbras jade hasta hacer arder la pared entera con la intensidad de una vidriera viva. Desde el techo de epidermis, a setenta micrómetros de altura, la luz cae difusa y casi paralela, filtrada por la cutícula como a través de vidrio esmerilado, encendiendo de blanco irisado el cielo irregular de polígonos que forman las células epidérmicas, mientras entre las columnas se abren crevices casi negras que sugieren pasadizos hacia las cavernas del mesófilo esponjoso más allá. Cada superficie húmeda luce un destello especular ultrafino, señal del film acuoso que impregna las paredes y garantiza el intercambio gaseoso, y donde dos cloroplastos se aproximan sobre el vacío sus bordes luminosos trazan líneas de neón verde contra la oscuridad absoluta del espacio intercelular. Es la arquitectura de la fotosíntesis operando en todas las direcciones a la vez: estructura pura, pigmento puro, química silenciosa y total.
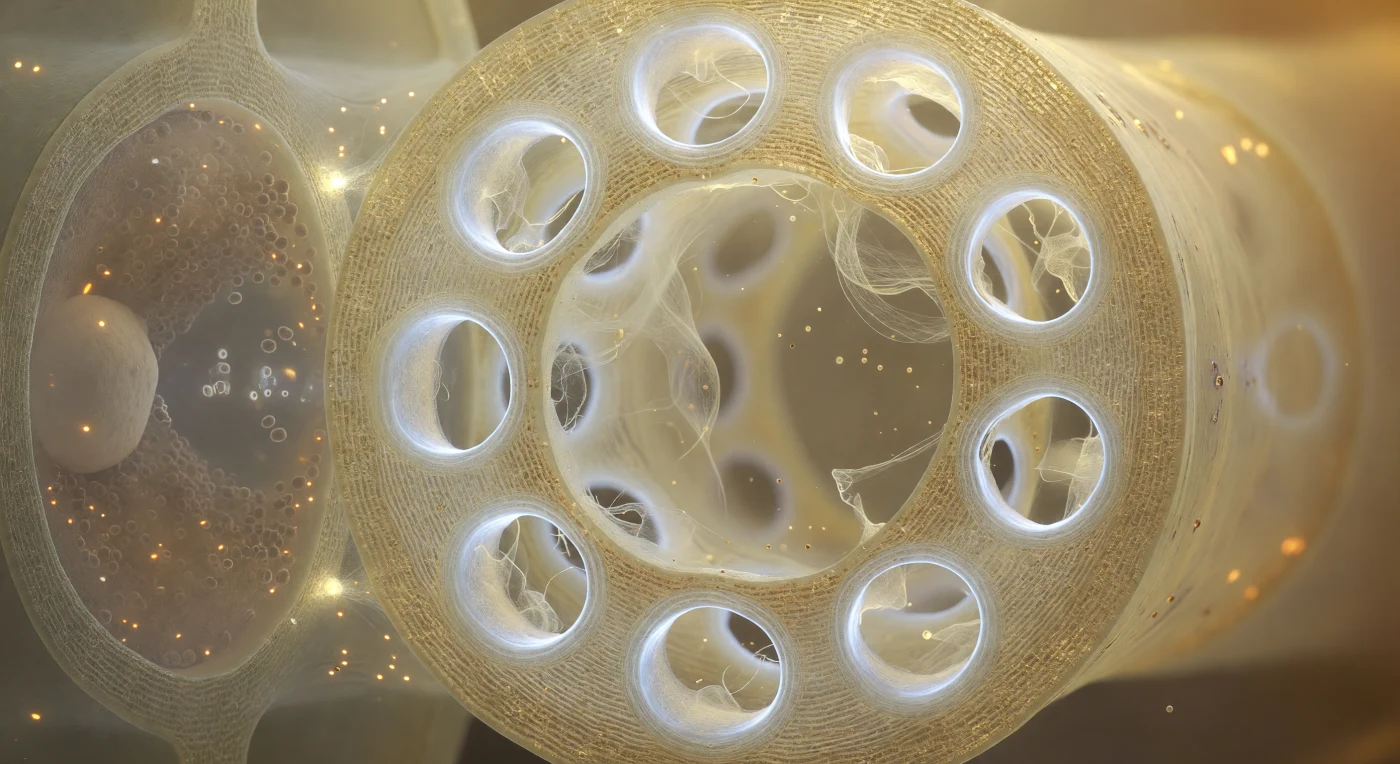
Dentro del lumen del elemento criboso, la placa cribosa cierra el campo de visión como un muro catedralicio de celulosa translúcida, su matriz color ámbar pálido perforada por doce poros dispuestos en constelación irregular, cada uno rodeado por un grueso collar anular de calosa que resplandece con una luminiscencia blanca y fría, ligeramente azulada en el borde interior donde estrecha la garganta hasta reducirla a un tercio de su diámetro original. A través de esos portales brillantes, filamentos de proteína P derivan como una malla iridiscente color crema aguada, algunos cruzando el poro como una cortina diáfana, otros apenas como hilos rotos de tela de araña que oscilan en la corriente casi imperceptible de la savia floemática que pulsa de una cámara a la siguiente a razón de aproximadamente un metro por hora, impulsada por gradientes de presión osmótica generados en las hojas fuente. El lumen que se extiende más allá irradia un halo dorado cálido, salpicado de gránulos proteicos suspendidos en solución coloidal, mientras que hacia la pared lateral compartida la célula acompañante se aprieta como una habitación oscura entrevista tras vidrio esmerilado, su citoplasma casi opaco de mitocondrias, ribosomas y retículo endoplásmico que contrastan con la claridad luminosa del tubo criboso. Las conexiones plasmodesmatales entre ambas células son apenas perceptibles como diminutas perforaciones brillantes en la pared compartida, canales moleculares que regulan el cargamento del floema y coordinan, a escala nanométrica, el destino del azúcar producido por cada hoja hacia cada raíz que crece en la oscuridad del suelo.
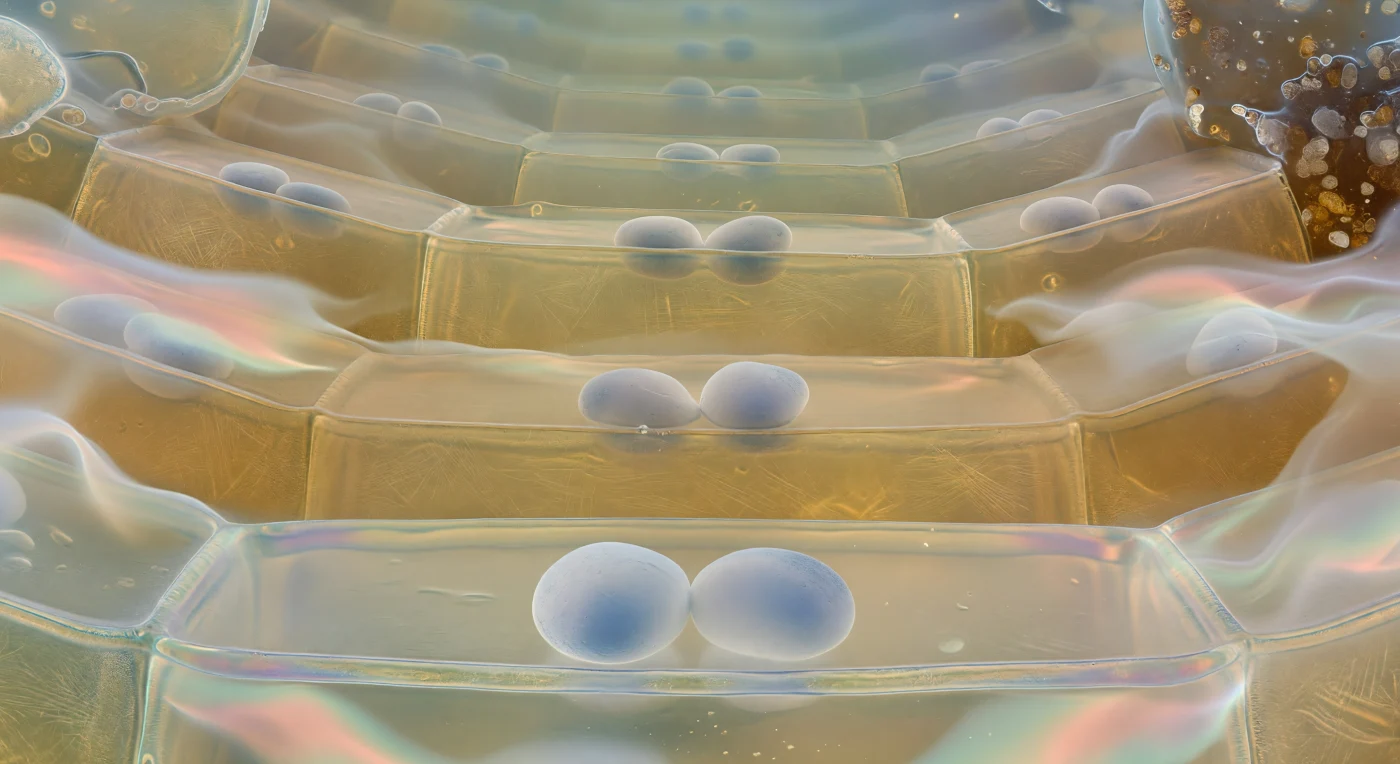
Suspendido justo por encima del suelo de una célula de la columela, la mirada se alza hacia una catedral de arquitectura viva donde enormes amiloplastos estatólitos reposan sobre la membrana plasmática inferior como bolos de piedra pulida sobre un parche de tambor tenso, sus superficies mate de blanco calcáreo sombreándose hacia un gris azulado allí donde la luz los atraviesa en difracción tenue. Las paredes celulares que se elevan a los lados son planos curvos de vidrio ambarino translúcido, y a través del techo semitransparente se percibe el suelo del siguiente nivel celular, con sus propios estatólitos difuminados en medias lunas blancas, repitiéndose el patrón hasta que la perspectiva piramidal se disuelve en un crepúsculo azul-verdoso donde las células más altas apenas son formas fantasmales flotando en la neblina de la zona meristemática. Estos estatólitos son amiloplastos densos en almidón cuya sedimentación gravitacional —renovada en cuestión de minutos cada vez que la raíz cambia de orientación— activa canales mecanosensibles y dispara cascadas de señalización auxínica que reorientan el crecimiento radical hacia el centro de la Tierra. En los márgenes del espacio visible, donde las células de la columela se funden con las células fronterizas periféricas, la arquitectura se deshace en un halo de mucílago iridiscente, ese hidrogel rico en polisacáridos que lubrica el avance de la raíz entre los granos de cuartzo y arcilla del suelo, capturando partículas minerales y colonias bacterianas en una interfaz bioquímica que convierte la frontera entre lo vivo y lo mineral en algo tan fluido y cambiante como el aceite sobre el agua quieta.

Bajo la oscuridad absoluta de los polarizadores cruzados, un cristal de drusas de oxalato de calcio se despliega ante ti como una rosa gótica mineralizada de cincuenta y cinco micrómetros de envergadura, sus cuarenta facetas irradiando desde un núcleo central con la inevitabilidad de la cristalografía, ardiendo en azul cobalto intenso, siena quemado y oro puro según el retardo óptico de cada prisma birrefringente. Cada faceta es un registro de un evento de precipitación iónica: cuando la concentración de calcio y oxalato superó el umbral de solubilidad dentro de la vacuola de esta célula parenquimática viva, los iones se ordenaron en redes cristalinas con una precisión molecular que ninguna ingeniería humana podría replicar a esta escala. Las crestas entre facetas adyacentes actúan como bordes prismáticos que concentran la luz polarizada en halos iridiscentes de color pavo real, mientras que el núcleo central denso —la semilla primordial de toda la estructura— genera una confusión espectral caótica donde los cristalitos interpenetrados superponen sus interferencias en magenta, verde pálido y cobre. En el límite mismo del campo visual, la pared celular de celulosa traza un marco dorado y tenue, recordándote que esta catedral mineral existe sellada dentro de un organismo que respira y crece, presionada desde afuera por el contenido acuoso de la vacuola que los polarizadores han disuelto en negro perfecto.

Suspendido en el aire inmóvil y saturado de humedad que flota sobre la cúpula del meristemo apical caulinar, la vista desciende sobre una bóveda levemente convexa tapizada en un mosaico casi perfecto de células isodiamétricas de unos 12 micrómetros de diámetro, cuyas paredes primarias de celulosa forman una retícula de marfil verdoso que irradia una translucidez suave, como si cada panel hexagonal fuera una lámina de papel de arroz iluminada desde dentro. En el centro de cada célula, visible a través de la pared difusa como una piedra oscura bajo vidrio esmerilado, reposa un núcleo denso y ovalado que delata la naturaleza de este tejido: células densamente cargadas de maquinaria biosintética, con apenas vacuola, repletas de ribosomas y orgánulos en una citoplasma casi opaco que habla de división inminente. Flanqueando el ápice a ambos lados, dos primordios foliares se elevan como contrafuertes de oro antiguo, sus células ya más elongadas y sus paredes ligeramente engrosadas, marcando el inicio de la diferenciación en una zona de transición de apenas dos o tres células de anchura donde el hexágono regular cede paso a una geometría más direccional. La zona axilar entre primordio y cúpula acumula un tinte azul-verdoso de la luz dispersada, acentuando el relieve suave de una arquitectura que, en menos de medio milímetro de diámetro, contiene el programa completo del cuerpo vegetal aún por construir.

Desde el interior de la cavidad subestomática, la mirada asciende hacia una arquitectura que no guarda escala humana pero la evoca con precisión asombrosa: dos células oclusivas se arquean sobre la cabeza como los doveles de un pórtico catedralicio, sus paredes internas engrosadas visibles como laminaciones de marfil pálido que resisten la presión osmótica y mantienen el poro abierto —siete micrómetros de cielo luminoso enmarcados por tensión diferencial de celulosa. A lo largo de cada célula oclusiva, doce cloroplastos se alinean como farolillos de jade eléctrico, presionados contra la pared interna por la misma presión de turgencia que infla la célula en su característica forma reniforme, sus membranas apiladas en grana captando la luz fría que desciende desde el exterior y devolviéndola como una luminiscencia propia. Los rebordes cuticulares sobresalen sobre el poro como labios de una cisterna translúcida, refractando la luz en un fleco iridiscente donde la cutícula se adelgaza hasta casi la escala molecular, mientras el vacío oscuro del poro se resuelve, al fijarse en él, en una columna de aire iluminada por el cielo abierto. Las células epidérmicas que rodean el estoma parecen, por contraste, losas grises y vacuas, sus citoplasmas apenas visibles, amplificando por comparación el resplandor casi incandescente de las células oclusivas que controlan, con la precisión de una válvula biológica, el intercambio de gases entre la planta y la atmósfera.

Te encuentras suspendido en el interior de una de las membranas generatrices más delgadas del reino vegetal: el cambium vascular en plena actividad primaveral, donde cada célula fusiforme se extiende ante ti como la nave de una catedral, treinta veces tu longitud de extremo a extremo, sus paredes tan finas que apenas merecen llamarse superficies. La citoplasma de color amarillo paja te envuelve como la luz dentro de un farol de pergamino, y el núcleo más cercano flota suspendido en ese halo ámbar como una esfera opalescente en la que el cromatín se enturbia suavemente, recordándote que aquí la vida aún no ha elegido su destino. A tu izquierda, las traqueidas xilémicas ya han tomado su decisión: depósitos concéntricos de pared secundaria se acumulan en bandas de ocre y óxido, la lignina sellando para siempre el interior en una necrópolis de tubos perfectamente diseñados que se pierden en la penumbra cobriza. A tu derecha, los elementos cribosos del floema se deshacen voluntariamente de sus núcleos y adelgazan su citoplasma hasta un gel traslúcido, sus placas cribosas abriéndose como halos de calosa luminiscente, entregándose al flujo. Entre esos dos imperios que solidifican o se disuelven, esta membrana casi transparente sostiene en silencio la entera arquitectura del árbol.

Ante ti se eleva la cabeza secretora de un tricoma glandular de tomate, sus cuatro células dispuestas en una corona aplanada de cúpulas translúcidas de color verde-dorado, cuyo citoplasma denso en orgánulos presiona suavemente contra la cara interna de la cutícula como huellas dactilares en vidrio tibio. Por encima de ellas, la cutícula ha sido empujada hacia afuera formando una ampolla tensa y reluciente —una lente de aceite esencial acumulado, teñida de ámbar y sutilmente refractiva— cuya superficie captura la luz oblicua en un destello líquido y lento que recorre el ecuador de la vesícula como el terminador sobre una pequeña luna; en su interior, gradientes de densidad y espirales pálidas traicionan la secreción estratificada de distintos compuestos terpenoides retenidos bajo la cutícula en una tensión creciente, en el instante preciso antes de la ruptura. Más abajo, el tallo celular de seis niveles desciende en escalones decrecientes —sus paredes compartidas visibles como líneas oscuras de pared primaria cementada con pectina— hasta anclarse en una epidermis de células con paredes anticlinas onduladas características del patrón de rompecabezas de las dicotiledóneas, sobre cuya superficie se cristaliza un escaso bosque de microcristales de cera cuticular que proyectan sombras duras y diminutas bajo la luz rasante. La luz que filtra a través del mesófilo superior tiñe las sombras de jade profundo y confiere al aceite ámbar un cálido resplandor secundario, como si ardiera iluminado desde dentro por su propia química, mientras la ampolla suspendida sobre la corona secretora brilla como el único objeto luminoso en un paisaje por lo demás mate, texturado y translúcido.

Ante ti se extiende una catedral viviente de luz acuosa, donde células de doscientos cincuenta micrómetros de altura se apilan extremo con extremo formando un corredor que se pierde en una infinidad cerúlea y translúcida. Cada célula está dominada casi por completo por una vacuola central de geometría perfecta —una cámara de fluido incoloro bajo una presión de turgencia tan colosal que las paredes de celulosa se curvan ligeramente hacia afuera, transmitiendo la luz como cristal marino esmerilado en un resplandor azul-blanco que se profundiza en aguamarina hacia las lejanías del tejido. El citoplasma vivo ha sido relegado a un film parietal apenas perceptible, una pátina verde espuma de mar aplastada contra la cara interior de cada pared, interrumpida aquí y allá por un núcleo de diez micrómetros —una esfera ambarino-pálida suspendida como una linterna al borde de un océano— cuya presencia revela de golpe la escala vertiginosa del espacio que lo rodea. Esta zona de elongación es donde la raíz conquista el suelo sin dividirse: las células absorben agua mediante aquaporinas, la vacuola se expande irreversiblemente empujada por el potencial hídrico, y las microfibrillas de celulosa se reorientan transversalmente para canalizar el crecimiento en el único eje que importa. Al fondo del corredor, a quizás cincuenta longitudes celulares de distancia, el cilindro vascular central emerge como una columna de índigo oscuro —pared lignificada, luz absorbida en lugar de transmitida— un eje vascular que ya transporta el agua que estas mismas vacuolas cedieron al apoplasto, clausurando el ciclo silencioso e hidráulico del crecimiento radical.

Dentro del grosor radial de la piel de un árbol, el mundo se construye en su totalidad con cámaras rectangulares encajadas con precisión milimétrica: células de corcho muertas cuyos tabiques de suberina irradian un ámbar cálido y oscuro, semejante a coñac viejo sostenido frente a una vela, mientras que sus lúmenes permanecen como vacíos absolutos, sellados en sus seis caras, sin fluido, sin orgánulo, sin ningún rastro de vida. La estructura es la del periderma, la cubierta protectora secundaria del árbol, donde el felógeno —una capa meristemática de división lenta— produce hacia afuera estas hileras de células suberificadas muertas que impermeabilizan y blindan al organismo frente a la desecación y los patógenos. De pronto, el amber se interrumpe en un umbral luminoso: el felógeno vivo presenta células traslúcidas de paredes delgadas como papel de arroz húmedo, y más adentro el felodermo muestra un verde difuso de cloroplastos, todo ello recordando que esta fortaleza mineral envuelve un interior metabólicamente activo. A un lado, la geometría perfecta se desmorona en un lenticelo, una zona de células de complemento incompletamente suberificadas que abren canales irregulares hacia el exterior, permitiendo el intercambio gaseoso que el propio impermeabilizante de suberin haría imposible en cualquier otro punto del muro. Estás de pie dentro de un cilindro que abraza un árbol entero, y cada pared de ámbar que te supera en altura es apenas una fracción de milímetro en el mundo de afuera.

Te encuentras suspendido en el interior de un canal de aerénquima del pecíolo de un nenúfar, un corredor cilíndrico de 800 micrómetros de diámetro cuya pared continua de células clorenquimáticas hexagonales irradia un verde esmeralda profundo y moteado, cada cámara celular separada de sus vecinas por tabiques de celulosa tan finos como pergamino húmedo. Este tejido lacunar no es un accidente estructural sino una arquitectura funcional precisa: los grandes espacios de aire interconectados del aerénquima permiten la difusión de oxígeno desde las hojas flotantes hasta los órganos sumergidos en condiciones de escasa aireación, convirtiendo el pecíolo en un sistema de ventilación interno. Flotando ante ti, el diafragma —una pantalla estelar de células en forma de brazo que irradian desde un núcleo central sin llegar a tocarse en sus extremos— atraviesa la totalidad del canal como un medallaje de encaje translúcido, celadón pálido en la base donde se concentran los cloroplastos y casi vítreo en las puntas, con sus vanos poligonales abiertos enmarcando la profundidad del corredor más allá. Una luz difusa retrocede desde el fondo del canal y transforma la pantalla estelar en una silueta de bordes dorados, mientras finas membranas de vapor iridiscente tiemblan en algunos de los vanos, proyectando interferencias de violeta y cobre sobre las paredes verdes: la escala de lo invisible hecha arquitectura viviente.
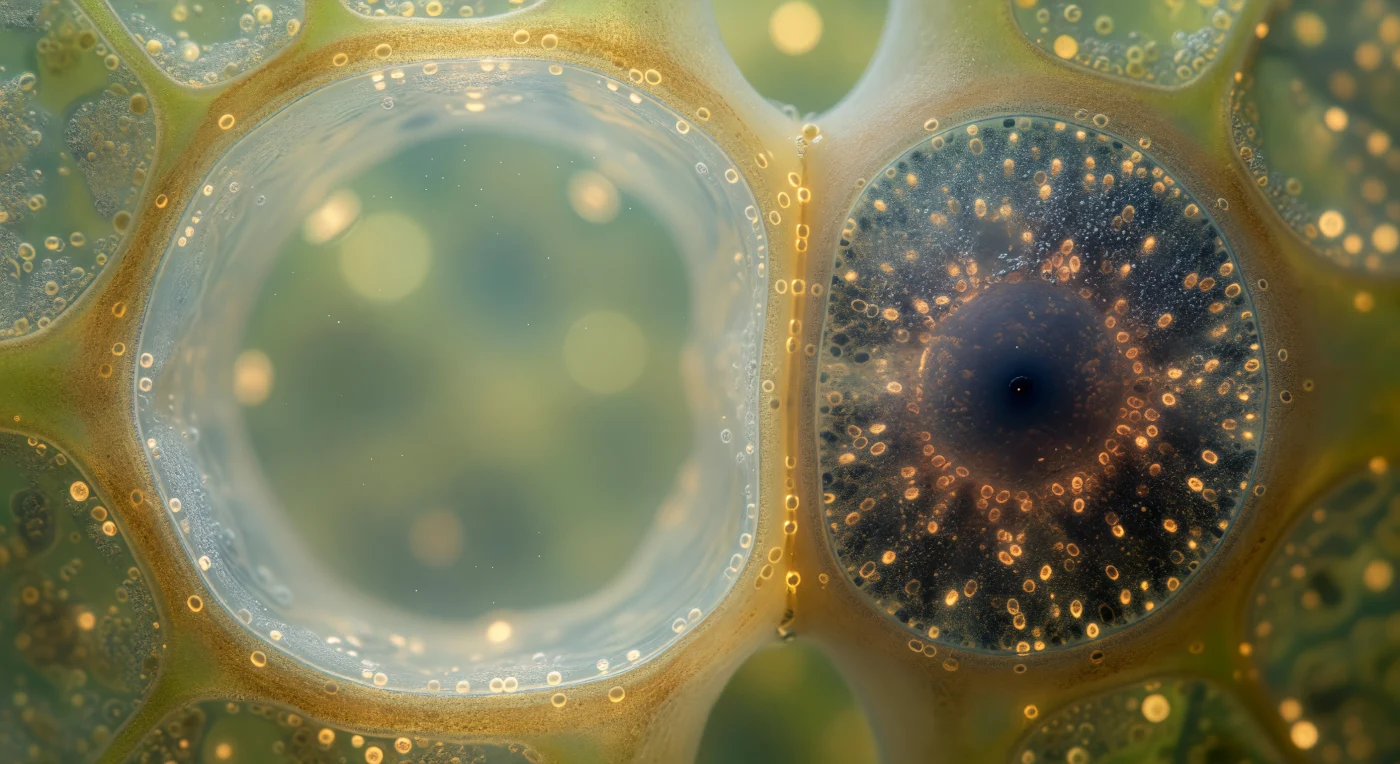
En este corredor vascular de un tallo vivo, la vista queda atrapada entre dos mundos celulares de contraste casi violento: a la izquierda, el elemento del tubo criboso se abre como una catedral de paredes ámbar translúcidas, su lumen casi vacío ocupado por un citosol acuoso que fluye en silencio, transportando azúcares disueltos desde las hojas hacia los órganos en crecimiento a una velocidad de cerca de un metro por hora, toda esa vastedad luminosa sostenida por apenas un filme parietal de citoplasma que se aferra a la pared como un susurro. A la derecha, la célula acompañante se aprieta contra esa pared como una forja en miniatura: su citoplasma oscuro y denso está repleto de mitocondrias —gránulos ovalados que irradian energía metabólica— y una escarcha granular de ribosomas que sintetizan sin pausa las proteínas necesarias para cargar y mantener el flujo del floema, todo organizado en torno a un núcleo redondo y autoritario que gobierna esta actividad frenética. La pared compartida entre ambas células —de apenas doscientos nanómetros de grosor en los puntos de contacto— está perforada por plasmodesmos que aparecen como una constelación puntada de oro cálido, nanoestructuras tubulares que comunican a la célula-río serena con su servidora metabólica en un diálogo molecular incesante. Esta pareja de luz y sombra encarna uno de los principios más elegantes de la biología vegetal: la división del trabajo entre una célula especializada en el transporte y otra que sacrifica toda su arquitectura interna al servicio de esa función.

En el umbral entre dos mundos, el observador se encuentra suspendido dentro de la raíz viva de una planta joven, mirando a lo largo de la fila endodérmica como si recorriera la nave de una catedral viva que se curva suavemente en ambas direcciones. Las células endodérmicas se alzan como columnas prismáticas de unos 30 micrómetros de anchura, sus paredes translúcidas y faintly verdosas como vidrio marino helado, encerrando citoplasmas granulares salpicados de inclusiones de almidón que centellan como especas de plata en un fluido ámbar lento. Lo que detiene la mirada es la banda de Caspary: un cinturón continuo de suberina y lignina depositado con precisión geométrica en cada pared radial compartida entre células vecinas, que resplandece con un ámbar anaranjado profundo —el color del cobre al rojo vivo o del ámbar iluminado desde dentro— formando un collar de fuego ininterrumpido que rodea completamente el estele sin un solo eslabón roto. Esta barrera no es ornamental sino funcional y absoluta: impide que el agua y los solutos atraviesen el apoplasto entre la corteza y el cilindro vascular, obligando a todo flujo mineral a cruzar la membrana plasmática de las propias células endodérmicas, donde proteínas transportadoras ejercen un control selectivo sobre lo que entra al xilema. Detrás, la corteza se abre en una arquitectura aireada de parénquima laxo con espacios intercelulares neblinosos en azul frío; adelante, el estele se condensa en una masa compacta de células procambiales de un intenso violeta-índigo, creando un contraste cromático tan dramático como el de un pasillo de forja que desemboca en una bóveda iluminada por la luna.

En esta latitud ecuatorial de un mundo construido en ámbar, te encuentras de pie sobre la superficie externa de un grano de polen de rosa, donde la exina —compuesta de esporopolenina, uno de los biopolímeros más resistentes que produce la vida vegetal— se ha organizado en una red reticulada de crestas continuas que definen celdas poligonales de unos 2,5 µm de anchura, cada una separando lúmenes en penumbra que descienden cerca de 1,5 µm hacia el interior de la pared del grano. Esta arquitectura no es aleatoria: las variaciones en la forma y densidad de las celdas reflejan los gradientes de deposición controlados por el tapeto del esporangio durante la microsporogénesis, cuando las proteínas precursoras fueron secretadas y polimerizadas en capas concéntricas alrededor del microsporocito. Hacia el horizonte, el paisaje se interrumpe en una franja pálida y translúcida —el colpo—, una de las tres aperturas germinales que atraviesan la exina de polo a polo, zonas donde la pared se adelgaza hasta casi desaparecer para permitir la emergencia futura del tubo polínico cuando el grano alcance el estigma. Todo el campo vibra con una luminiscencia dorada propia del material, la autofluorescencia característica de la esporopolenina bajo iluminación oblicua, convirtiendo cada cresta en una muralla iluminada y cada lumen en un pozo de sombra cálida, recordando que esta estructura microscópica ha evolucionado durante más de cuatrocientos millones de años como la primera armadura biológica capaz de resistir la desecación, la luz ultravioleta y la digestión enzimática.
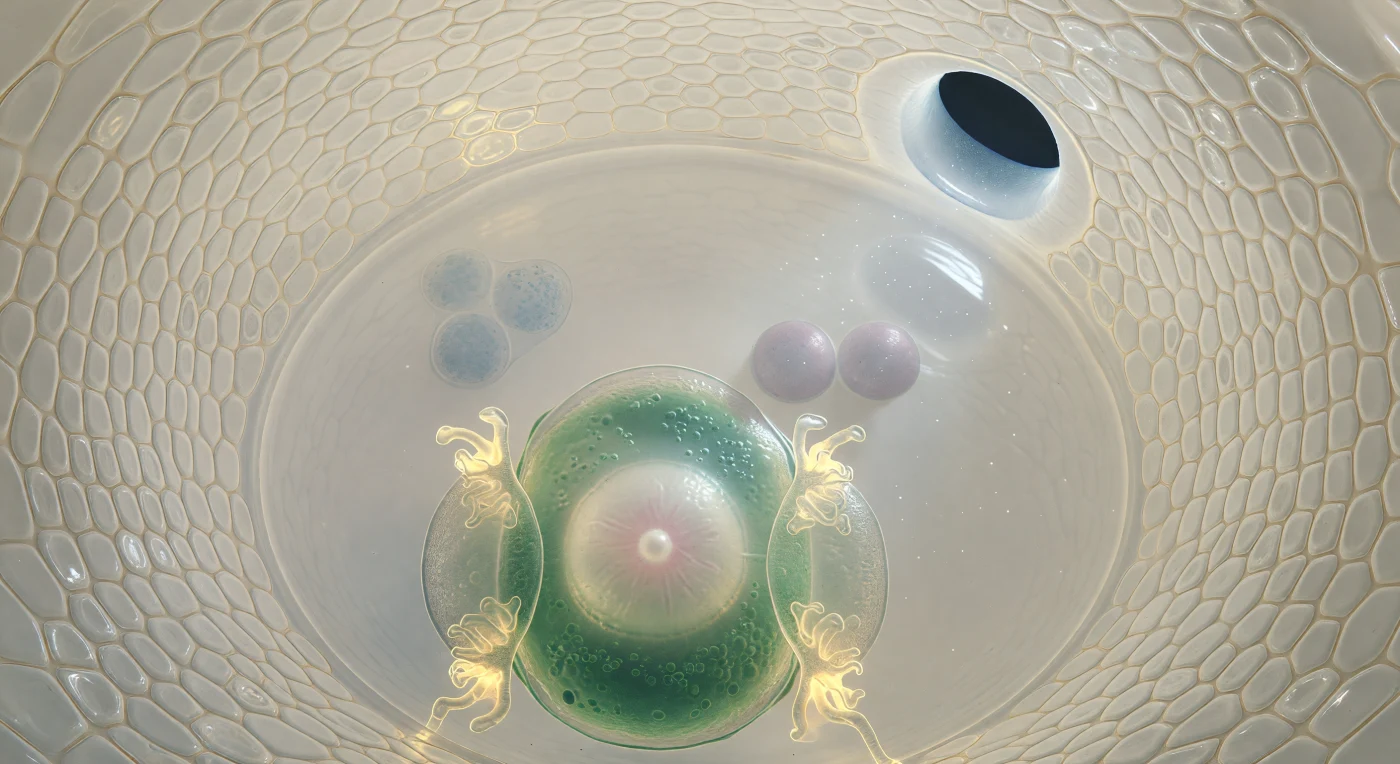
Te encuentras suspendido en el interior de un saco embrionario casi maduro, a apenas cuarenta micrómetros de la superficie convexa de la célula huevo, rodeado por las dos capas de integumentos que se curvan a tu alrededor como las bóvedas de una gruta de marfil vivo, sus paredes una mosaico translúcido de células apretadas cuyos límites se revelan como finas costuras ámbar bañadas en una luminosidad difusa y sin origen aparente. En el extremo superior derecho, la micrópilaforma un pasaje oval de apenas quince micrómetros, por el que se cuela un haz de luz exterior que ilumina partículas suspendidas en el fluido apoplástico y arroja un cono suave sobre la pared más próxima. La célula huevo domina el primer plano: una masa piriforme de citoplasma jade oscuro en su periferia organelar que se aclara hacia un núcleo central, una esfera albastrina surcada de un levísimo rosa con el nucléolo brillando como una perla quieta, flanqueada por las dos sinérgidas cuyos rizos filiformes relucen en ámbar dorado concentrado, como brasas diminutas suspendidas en el espacio. Más allá, la célula central se abre en una cámara inmensa de citoplasma tan transparente que parece agua quieta, donde dos núcleos polares flotan como lunas lilas suspendidas en un océano inmóvil, y en el polo calazal más remoto, tres células antípodas se agrupan difuminadas como piedras vistas a través de agua profunda y clara. Todo este universo encapsulado, escenario inminente de la doble fecundación angiosperma, tiene el tamaño de un grano de polvo invisible al ojo desnudo, y sin embargo contiene la totalidad de la arquitectura celular necesaria para dar origen a una nueva planta.

Estás suspendido en el umbral exacto donde el roble registra el paso de un año entero: a tu izquierda, los vasos del xilema temprano se abren como túneles de catedral de 250 µm de diámetro, sus lúmenes absolutamente negros rodeados por halos ámbar donde las paredes lignificadas con punteaduras areoladas capturan la luz polarizada como molduras de arquitectura petrificada. A tu derecha, el límite cae como un acantilado sin transición: las fibras libriformes de la madera tardía se apiñan en una masa casi sólida donde los lúmenes se reducen a rendijas de 15 µm, y bajo la luz polarizada sus paredes secundarias estallan en azul cobalto eléctrico y oro bruñido, un mosaico de esmalte bizantino que irradia energía mecánica almacenada. Cruzando ambos mundos en horizontal, las cintas de parénquima radial avanzan como estratos geológicos de miel pálida, sus células en forma de ladrillo articuladas con precisión, indiferentes a la frontera que atraviesan, llevando una calidez mate que ancla el caos iridiscente a su alrededor. Todo el conjunto arde como una vidriera orgánica, cristalino y monumental, el registro silencioso de una estación entera codificado en la arquitectura celulósica del árbol.
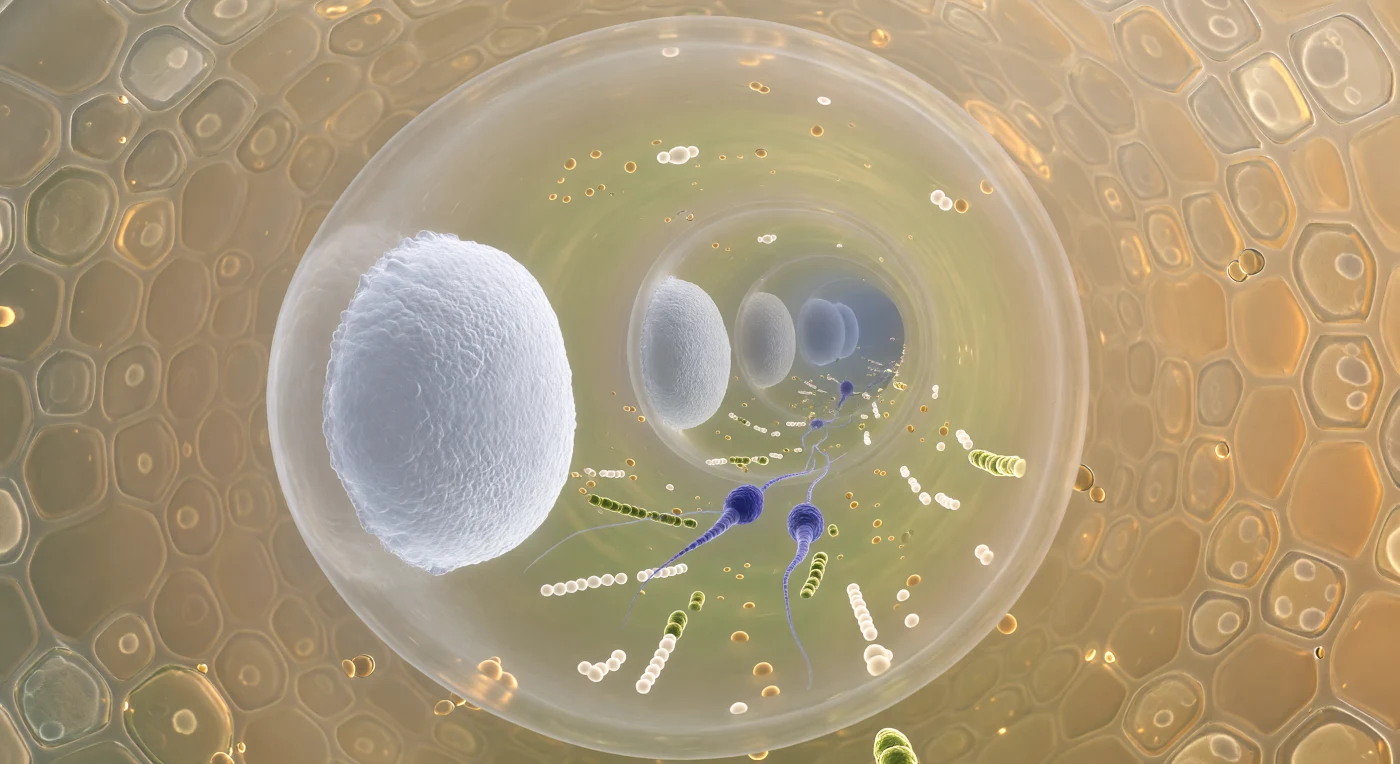
En este instante suspendido en el interior de un tubo polínico, la mirada avanza por un corredor cilíndrico de apenas doce micrómetros de diámetro, cuya pared de celulosa y calosa se curva en torno a nosotros como un tubo de vidrio ámbar levemente luminoso. A lo largo del eje del canal, una sucesión de tabiques blancos —los tapones de calosa— se alza como mamparos de porcelana traslúcida, cada uno separado del siguiente por unos ochenta micrómetros, creando una columnata de cámaras que se pierde hacia la punta en crecimiento en una perspectiva de discos cada vez más pálidos y azulados. Entre los dos tabiques más próximos, el citoplasma fluye como un río presurizado de orgánulos —esferas lipídicas ámbar, mitocondrias amarillo-verdosas, vesículas en caravana— y en medio de esa corriente avanzan dos células espermáticas fusiformes, de apenas cinco micrómetros de longitud, teñidas de un intenso violeta índigo que contrasta con el fondo dorado del fluido, su cromatina tan condensada que parece mineral sólido, arrastradas sin esfuerzo hacia el óvulo en espera. Más allá de la pared del tubo, el tejido transmisor del estilo presiona desde fuera como una mosaico de células secretoras doradas, embebidas en una matriz de polisacáridos refractivos y húmedos que bañan el exterior con una niebla ambarína cálida, recordando que este corredor microscópico atraviesa un organismo vivo cuya escala nos resulta tan íntima como abrumadora.