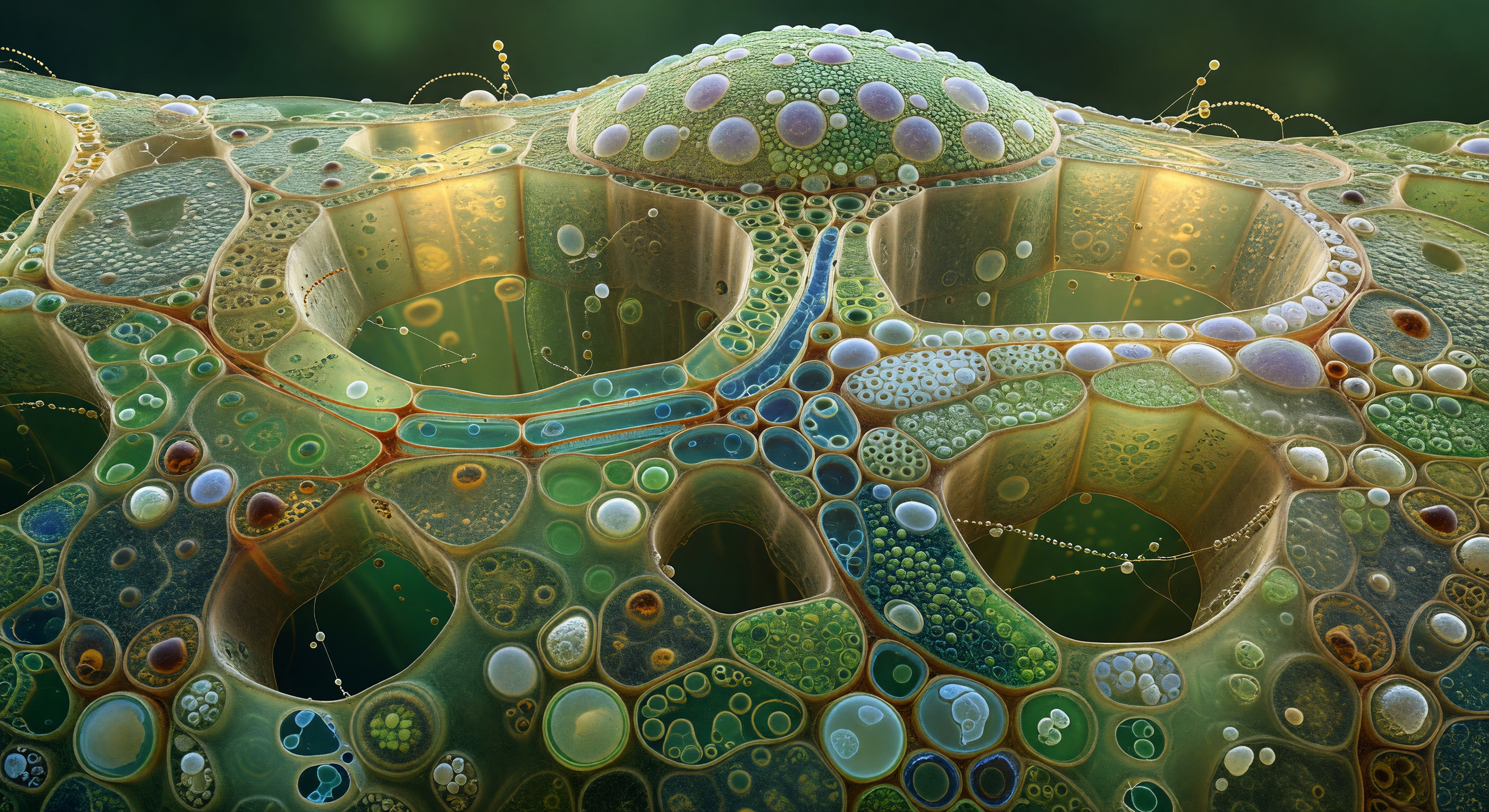
O observador encontra-se no interior de um corredor cilíndrico de cento e vinte micrómetros de diâmetro, esculpido inteiramente em lenhina e celulose secundária, cujas paredes âmbar-siena exibem centenas de pontuações areoladas dispostas em fileiras geométricas com regularidade quase cristalográfica — cada depressão circular de seis micrómetros orlada por um lábio ligeiramente saliente que capta a luz difusa como uma lua em quarto crescente. Estas pontuações não são meros ornamentos: são as válvulas de pressão do sistema vascular, membranas fantasmáticas nacaradas que mediam o fluxo de água entre vasos vizinhos sob tensão hidráulica negativa, sustentando colunas de líquido que sobem dezenas de metros em direção à luz do dossel sem qualquer bomba mecânica. O vaso está completamente vazio, em tensão activa, e o silêncio e a secura são absolutos — não há líquido aqui, apenas o sentido táctil do vácuo e da coesão molecular a segurar o vazio. Ao fundo do corredor, onde a perspetiva comprime o padrão de círculos repetidos numa tapeçaria ponteada que se afasta na penumbra, uma única bolha de embolismo de ar ocupa o lúmen como um espelho de mercúrio convexo, refletindo toda a galeria de pontuações em miniatura distorcida — uma fratura silenciosa no sistema de transporte, bela como uma fissura num cristal, imóvel na fronteira entre coesão e cavitação.

Você está no fundo de uma catedral viva, cercado por torres cilíndricas que se erguem setenta micrómetros acima da sua cabeça — as células do mesófilo em paliçada, cada uma com paredes de âmbar translúcido reforçadas por teias diagonais de microfibrilas de celulose, cobertas por dentro de centenas de cloroplastos biconvexos que se encaixam como azulejos de esmeralda, os seus granas internos visíveis como bandas escuras quando a luz os atravessa em ângulo oblíquo. A luz desce diretamente do tecto, a epiderme superior, que brilha a um branco marfim suave e ligeiramente oleoso através da cera da cutícula, como luz de inverno filtrada por vidro fosco — difusa, quase sem sombras duras, descendo em cortinas paralelas que as paredes clorofiladas tingem de verde-jade antes de se perder na escuridão entre as colunas. Os espaços intercelulares abrem-se como fendas negras entre os pilares, passagens estreitas que dão acesso ao mesófilo esponjoso mais além, sugerindo cavernas internas onde o dióxido de carbono e o vapor de água circulam livremente num microclima de humidade quase total. Em cada superfície curva existe um filme aquoso ultrafino que apanha a luz descente como um verniz húmido, e onde dois cloroplastos se aproximam de lados opostos de uma fenda, as suas arestas verdes brilham no negro como neon. É aqui, neste silêncio arquitectónico de pigmento e estrutura, que a fotossíntese acontece em cada direcção em simultâneo — luz a transformar-se em química a uma velocidade que nenhum relógio humano consegue acompanhar.
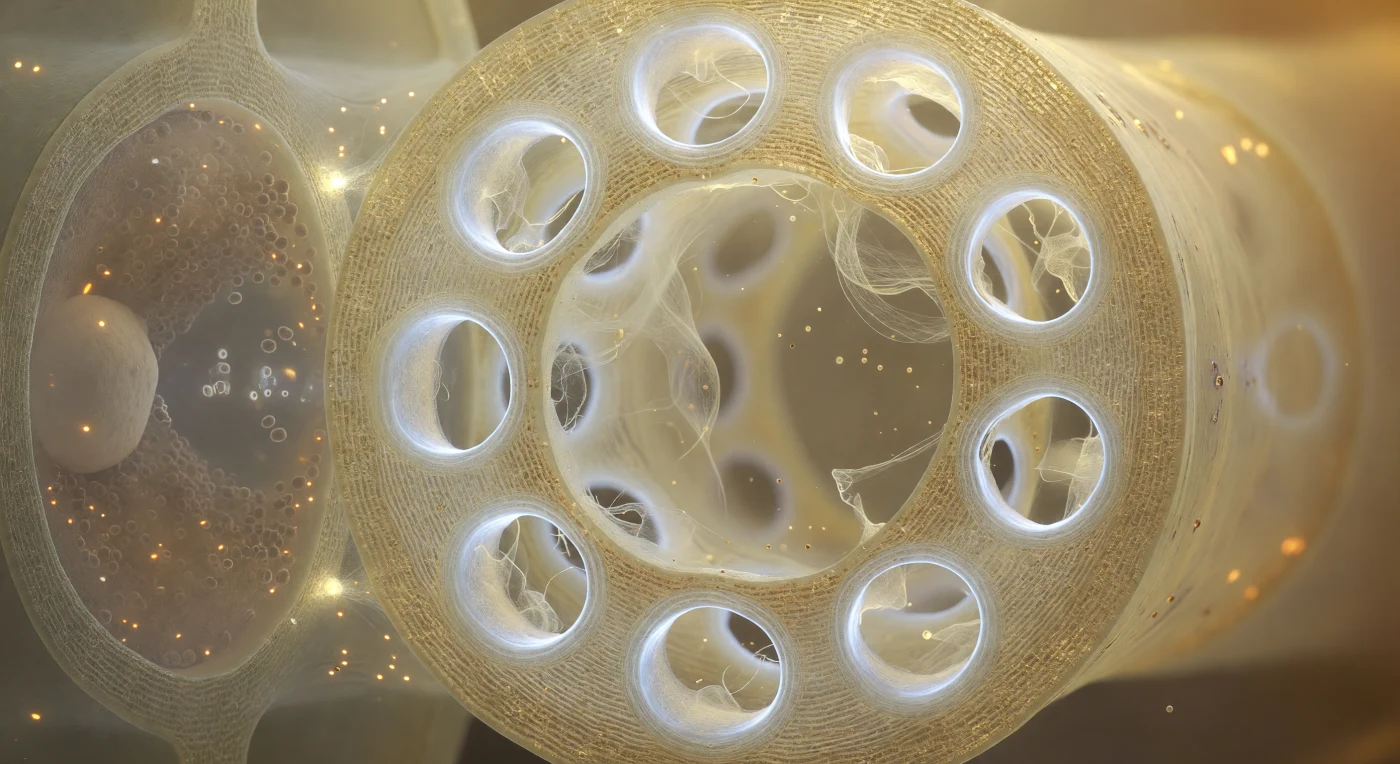
Dentro do lúmen de um elemento de tubo crivoso, a parede terminal ocupa todo o campo visual como uma antiga tela de catedral — uma partição dourado-âmbar de celulose translúcida, perfurada por doze poros dispostos em constelação irregular, cada um orlado por um espesso colar anular de calose que irradia uma luminescência branco-leitosa ligeiramente azulada na borda interna, onde a biopolímero se deposita em lamelas concêntricas densas e estreita a garganta de cada poro. Através dessas aberturas, filamentos de proteína P derivam em malha translúcida cor de creme iridescente, ora como cortina diáfana suspensa no fluxo quase imperceptível da seiva do floema, ora como fios isolados a tremer semelhantes a teia partida — vestígios de um sistema de bloqueio rápido que sela os poros quando a pressão de turgescência é subitamente interrompida. À esquerda, através da parede lateral compartilhada, a célula companheira pressiona contra o elemento crivoso como um quarto escuro visto por vidro fosco: o seu interior é quase opaco de densidade biológica — mitocôndrias como brasas alaranjadas, ribossomas e retículo endoplasmático formando uma névoa granular castanho-acinzentada, e um núcleo pálido encostado a uma face, coordenando o carregamento ativo de açúcares que alimenta o fluxo de massa pelo floema a cerca de um metro por hora. O contraste entre a claridade luminosa e âmbar do lúmen crivoso e a penumbra densa da célula companheira condensa, neste espaço de poucos micrómetros, toda a arquitectura funcional do transporte de fotoassimilados na planta viva.
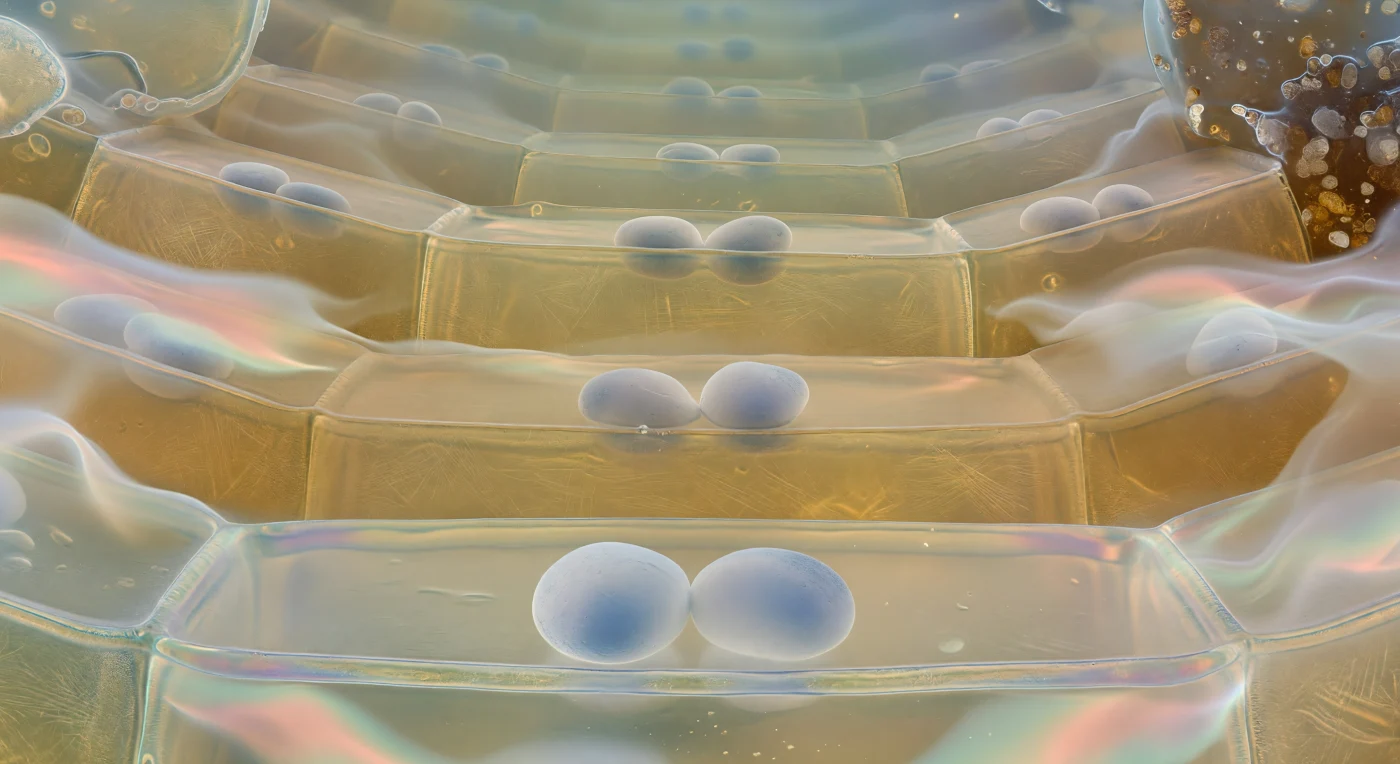
Suspenso logo acima do pavimento de uma célula da columela, o olhar sobe por uma catedral de arquitectura viva que recua em distância âmbar-esverdeada até um cume distante e luminoso: em primeiro plano repousam dois ou três amiloplastos estatólitos directamente sobre a membrana plasmática inferior, cada um um ovoide denso e levemente translúcido do tamanho de um rochedo nesta escala, de superfície branca-giz a tingir-se de azul-cinzento onde a luz os atravessa, perfeitamente imóveis sobre a membrana como pedras polidas largadas numa pele de tambor iridescente em violeta pálido, dourado e azul-gelo. Estes estatólitos são os gravímetros da planta: organelos ricos em amido cujo peso sedimenta em resposta à gravidade, sinalizando à raiz a direcção do crescimento geotrópico com uma precisão que desafia a intuição para estruturas tão pequenas. As paredes celulares ascendem como planos curvos de vidro âmbar translúcido, e através do tecto semi-transparente entrevêem-se os estatólitos do tier seguinte como crescentes brancos e enevoados, repetindo-se a geometria pirâmide acima, câmara sobre câmara, até as células do topo se dissolverem numa névoa azul-verde fria da zona meristemática. Na periferia, as células-fronteira desintegram as suas membranas numa bruma de mucilagem hidrogélea e iridescente que captura partículas minerais do solo, formando um halo bioquímico glistening que mantém à distância a pressão hidráulica da terra escura e comprimida lá fora.

Pairado no coração geométrico de um cristal de oxalato de cálcio, o olhar encontra uma explosão mineral de quarenta facetas que irradiam de um núcleo central denso como pétalas de uma rosácea gótica, cada uma delas um prisma birefringente ardendo em azul-cobalto profundo, âmbar queimado e ouro puro contra um fundo de negro absoluto — o vazio criado pelos polarizadores cruzados que aniquilam toda a água vacuolar circundante, tornando invisível o meio aquoso que envolve o cristal. Esta estrutura de 55 micrómetros é um registo mineralizado de eventos de saturação iónica: quando a concentração de iões cálcio e oxalato ultrapassou o limiar de solubilidade no interior da vacúola, a precipitação cascadeou outward a partir de um núcleo primordial, cada ramificação cristalina crescendo com inevitabilidade cristalográfica até terminar numa ponta de estilete mergulhada na escuridão. As cristas entre facetas adjacentes funcionam como arestas de prisma que refractam a luz polarizada em halos iridescentes de pavão, e onde dois planos cristalinos se interpenetram perto do núcleo, a interferência de cores torna-se caótica — cobalto a sangrar em magenta, ouro em verde pálido — revelando uma micro-topografia de terraços de crescimento que só existe aqui, a esta escala, dentro da célula viva. À periferia do campo visual, mal perceptível, a parede da célula parenquimática traça uma moldura de ouro pálido feita de microfibrilas de celulose, lembrando que toda esta catedral cristalina existe selada dentro de um organismo que respira, a pressão vacuolar pressionando silenciosamente para fora contra essa fronteira viva.

Suspenso a uma distância mínima acima do ápice caulinar vegetativo, o olhar desce sobre uma cúpula levemente convexa pavimentada por um mosaico quase perfeito de células isodiamêtricas, cada uma com cerca de doze micrómetros de lado, cujas paredes de celulose primária formam uma grelha cor de marfim-menta translúcida — fina o suficiente para dispersar a luz difusa como painéis de vidro fosco, deixando visível em cada célula um núcleo oval e escuro pousado no centro como uma semente sob gelo. Este é o meristema apical caulinar, o reservatório de células pluripotentes que governa toda a arquitectura futura da planta: a zona central mantém um núcleo de divisão lenta, enquanto a zona periférica recruta rapidamente células fundadoras dos primórdios foliares que aqui emergem nos flancos como duas cristas douradas, com paredes já mais espessas e células que começam a alongar-se, rompendo a regularidade hexagonal em geometrias direccionais. O citoplasma de cada célula é denso, quase sem vacúolo, saturado de ribossomas e organelos que conferem ao interior uma textura granular âmbar, pois aqui não existe espaço desperdiçado — só a maquinaria molecular da morfogénese a trabalhar em regime contínuo. Contemplar esta superfície é contemplar o momento anterior a toda a forma vegetal, o instante em que a informação genética se converte, célula a célula, na arquitectura de um ser vivo.

O observador encontra-se suspenso dentro da cavidade subestomática de uma folha viva, olhando para cima em direção à abertura que rasga a epiderme como um portal luminoso: duas células-guarda arqueiam-se sobre a cena como as aduelas de uma abóbada gótica, cada uma inchada de pressão de turgor até adquirir a silhueta característica de rim, emoldurando uma fenda de sete micrómetros por onde o céu externo derrama uma luz branca e fria que contrasta absolutamente com o verde elétrico das membranas fotossintéticas. Ao longo das paredes internas espessadas de cada célula-guarda — cujas laminações de microfibrilas de celulose, densamente empacotadas, resistem mecanicamente à pressão osmótica e mantêm o poro aberto — doze cloroplastos dispõem-se em fila cerrada como lentes de jade polido, a sua fluorescência interna tornando-os quase autoluminosos contra a solução vacuolar translúcida que os envolve. As bordas cuticulares projetam-se sobre as margens do poro como os lábios de uma cisterna de vidro, finas ao ponto de refratar a luz em franjas iridescentes de ouro e violeta pálido onde a espessura da cutícula se reduz à escala molecular. Em torno desta abóbada viva, as células epidérmicas pavimentosas recuam em anonimato — vastas, descoradas, de citoplasma quase inexistente —, e abaixo desta posição de observação o mesófilo esponjoso abre-se em labirinto de câmaras úmidas onde filmes de vapor condensado cintilam sobre cada superfície côncava, dissolvendo o espaço mais profundo numa névoa biológica e luminosa.

Suspended no interior de uma célula fusiforme do câmbio vascular durante o despertar primaveril, o observador encontra-se dentro de uma estrutura viva que se estende à sua volta como a nave de uma catedral, com paredes tão finas e translúcidas que parecem respirar — membranas de seda molhada separando dois mundos em transformação oposta. À esquerda, os traqueídos do xilema recém-formados já se armam com camadas concêntricas de parede secundária lignificada, o âmbar a aprofundar-se progressivamente numa necrópole de tubos perfeitamente engenhados que recuam para a sombra acastanhada; à direita, os elementos crivosos do floema desfazem-se voluntariamente, os núcleos a dissolver-se em véus fantasmáticos enquanto os poros das placas crivosas se abrem como halos opalescentes de calose, tudo ali a tornar-se permeável ao fluxo. Entre estes dois impérios a solidificar — um que endurece, outro que se entrega à corrente —, as iniciais fusiformes existem como uma membrana gerativa quase transparente, banhadas numa luminescência amarelo-palha contínua, o núcleo de cada célula suspenso como uma esfera opalescente no centro desse haze de âmbar, o cromatino a nublar-se como fumo em resina. É aqui, nesta zona de compressão silenciosa entre dois destinos celulares irrevogáveis, que a planta decide o que cresce, o que conduz água e o que transporta açúcar — tudo a partir de células tão finas que a sua própria existência parece uma questão de sussurro osmótico.

À superfície desta folha, ergue-se diante de você uma estrutura que, à primeira vista, evoca uma baliza luminosa numa paisagem de platôs e cristas: a cabeça secretora de um tricoma glandular de tomateiro, com suas quatro células discóides dispostas em coroa baixa, cada uma um domo translúcido de citoplasma verde-dourado pressionando delicadamente contra a cuticula como impressões de polegar em vidro morno. Sobre essa coroa, a cuticula foi empurrada para fora em uma bolha tensa e reluzente — uma lente de óleo essencial acumulado, tingida de âmbar e sutilmente refrativa, cuja superfície captura a luz oblíqua em um brilho líquido e curvo que percorre o equador da bolha como o terminador numa lua em miniatura, revelando nos gradientes internos as camadas sucessivas de compostos terpenóides secretados pelas células abaixo. O pedúnculo de seis andares desce em tiers decrescentes de pares celulares, suas paredes compartilhadas visíveis como linhas escuras de parede primária cimentada por pectina, toda a coluna exibindo uma gravidade quase arquitetônica antes de se ancorar na epiderme subjacente, onde a célula basal se alarga e espessa para resistir ao suave balanço turgescente da folha. Ao redor, a superfície epidérmica estende-se como uma planície tectônica de células com paredes anticlinais onduladas — o padrão característico de quebra-cabeça das dicotiledôneas —, salpicada de micro-cristais de cera cuticular em forma de bastonetes que projetam sombras rígidas sobre o fundo mate e levemente azulado, fazendo com que a bolha de óleo acima brilhe como o único objeto luminoso numa paisagem de paredes translúcidas e cristas de cera silenciosas.

Você se encontra dentro de um corredor vivo que se estende à sua frente como uma catedral aquática sem fim, cada célula erguendo-se como uma câmara de duzentos e cinquenta micrómetros de altura, quase inteiramente ocupada por um vacúolo central colossal cujo conteúdo hídrico, transparente como cristal, mantém as paredes de celulose arqueadas para fora sob uma pressão de turgor silenciosa e absoluta. A vida propriamente dita retrocedeu para as margens: o citoplasma vivo não passa de um filme parietal ténue, de um verde-espuma pálido, colado contra a face interna da parede como uma memória biológica, espessando-se apenas onde ocorrem plasmodesmas ou onde um núcleo âmbar de dez micrómetros — uma lanterna suspensa numa película de oceano — revela, pela sua própria pequenez, a dimensão vertiginosa do espaço que o rodeia. As paredes de microfibrilas de celulose transmitem a luz como vidro marinho fosco, emitindo um brilho azul-branco que percorre toda a coluna celular e se vai aprofundando em aquamarino à medida que o corredor recua para a distância. Ao fundo, talvez cinquenta células adiante, a estela central impõe-se como uma coluna axial índigo, densa e escura, onde as paredes lenhificadas do proto-xilema absorvem a luz em vez de a deixar passar, ancorando este espaço de água e pressão numa anatomia que move, em silêncio, toda a seiva da raiz para cima.

Estás de pé dentro da espessura radial da pele exterior de uma árvore, olhando em direção ao interior vivo, e o mundo que te rodeia é uma fortaleza de câmaras rectangulares mortas — células de cortiça cujas paredes suberizadas irradiam um âmbar quente de mogno, espesso e translúcido como cognac velho segurado contra uma chama, enquanto os seus lúmenes são vazios absolutos, caixas seladas em seis faces que nunca mais admitirão fluido, organelo ou citoplasma algum. Cada parede de suberin, construída em lamelas sobrepostas durante anos de actividade do felogénio, é uma barreira impermeável a gases e água, responsável por tornar a cortiça um dos materiais de isolamento mais notáveis do mundo biológico — e aqui, nesta secção radial, as filas de células empilham-se em colunas precisas que se estendem até um horizonte que curva imperceptivelmente, lembrando-te que estás dentro de um cilindro que envolve toda a árvore viva. Abruptamente, o âmbar profundo cede a uma única camada do felogénio — células achatadas de paredes finas como papel de arroz molhado, vivas de uma forma que a fortaleza atrás delas nunca voltará a ser, os seus interiores levemente luminosos com o azul-verde diluído do citoplasma ativo, uma membrana biológica tensa sobre todo o campo visual como um limiar resplandecente. Para o lado, a quarenta larguras de célula de distância, a geometria ordenada da cortiça dissolve-se num lenticelo — uma zona pálida e fragmentada onde as paredes suberizadas se tornam incompletas e os espaços entre células abrem canais irregulares que exalam para a vastidão exterior, uma fenda de respiração onde a fortaleza hermética admite finalmente o mundo.

Você flutua no interior de um canal de aerênquima do pecíolo de um nenúfar, suspenso numa atmosfera de vapor saturado de umidade, envolto por uma abóbada cilíndrica cujas paredes formam um mosaico contínuo de células poligonais repletas de cloroplastos — cada uma com cerca de duzentos micrômetros de largura, seus septos de celulose finos como papel molhado e levemente âmbar nas bordas. O aerênquima é um tecido especializado de transporte gasoso presente em plantas aquáticas, onde grandes lacunas de ar intercelulares sustentam a difusão de oxigênio e dióxido de carbono ao longo do caule e do pecíolo, permitindo que a planta sobreviva em ambientes encharcados e com baixa disponibilidade de ar. Diretamente à sua frente, atravessando todo o diâmetro do canal, um diafragma flutua como uma renda suspensa no vazio: uma tela estrelada de células alongadas que irradiam de um núcleo central, suas pontas arredondadas quase — mas não completamente — tocando a parede do canal, deixando aberturas poligonais abertas pelas quais seria possível deslizar sem resistência. Iluminada por trás por uma luz difusa que percorre o comprimento úmido do canal — talvez autofluorescência da clorofila ou simplesmente luminosidade filtrada ao longo do tecido —, a tela transforma-se numa silhueta incandescente de extraordinária delicadeza, cada braço celular orlado por um halo dourado-esverdeado onde a membrana refrata a luz transmitida, enquanto finas películas de água iridescente tremem nas aberturas, projetando interferências de violeta e cobre nas paredes esmeraldas ao redor.
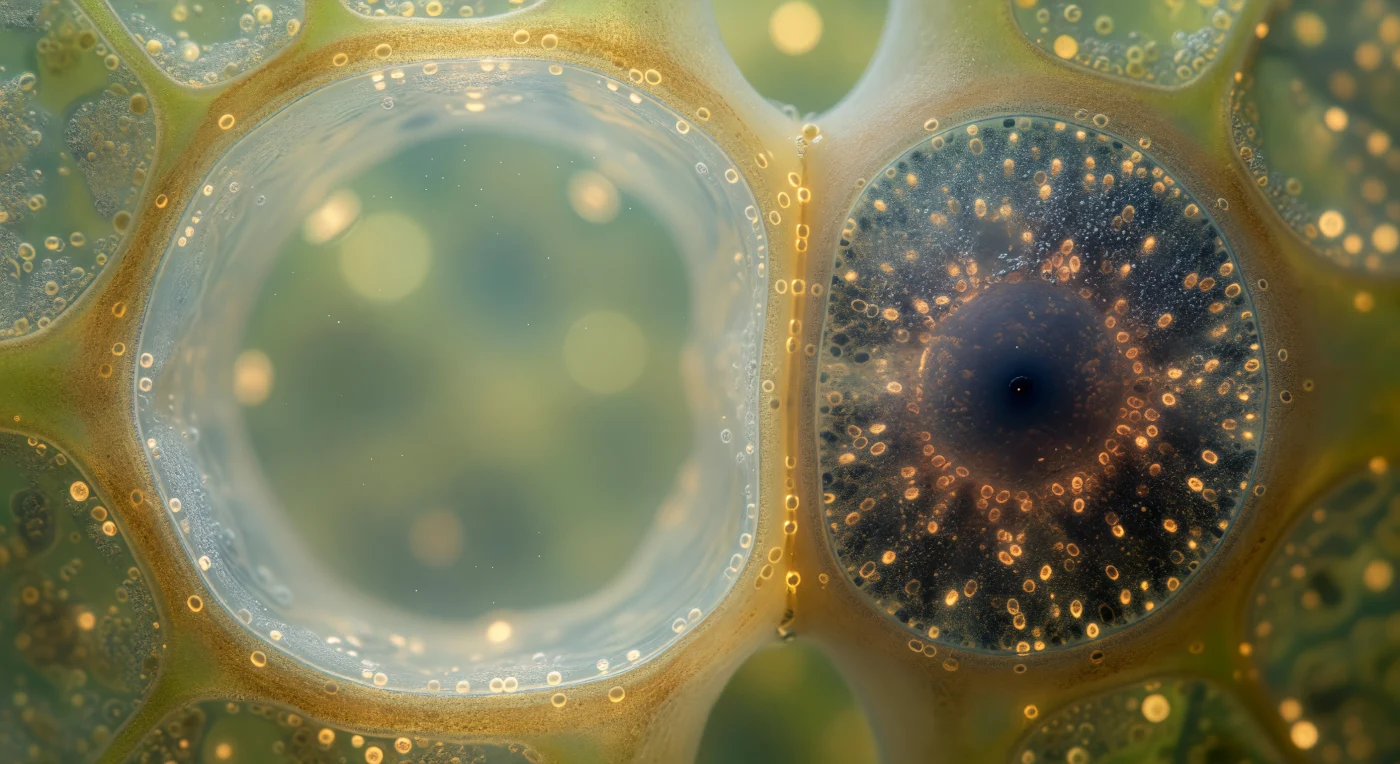
Suspenso no corredor vascular de um caule vivo, o observador encontra-se na fronteira entre dois mundos celulares radicalmente opostos: à esquerda, o elemento do tubo crivoso abre-se como uma catedral quase vazia, de paredes âmbar translúcidas e lúmen aquoso onde uma corrente invisível de açúcares dissolvidos percorre o silêncio a cerca de um metro por hora, a camada citoplásmica parietal reduzida a um véu nacarado tão fino que mal se distingue da parede celular. À direita, comprimida contra essa parede luminosa como uma sombra com massa, a célula companheira condensa em um quarto do volume toda a urgência metabólica do floema — citoplasma denso e quase opaco, saturado de mitocôndrias ovais que irradiam o calor da respiração celular intensa, ribossomas que formam uma névoa granular e um núcleo redondo e soberano, semelhante a um forge em plena combustão visto por uma janela estreita. Entre as duas células, a parede partilhada está perfurada por plasmodesmos — canais cilíndricos de duzentos nanómetros de espessura que se lêem como uma constelação de pontos âmbar quente — através dos quais a célula companheira regula ativamente o carregamento de sacarose no tubo crivoso, compensando com o seu metabolismo fervilhante a quase total ausência de organelos no elemento condutor que ela serve e sustenta.

O que se vê é uma secção transversal do interior de uma raiz jovem ao nível da endoderme, uma única fila de células prismáticas e translúcidas — cada uma com cerca de trinta micrómetros de largura — que formam uma fronteira contínua entre o córtex exterior e o estelo interior. Na parede radial de cada célula, exatamente onde as células vizinhas se tocam, corre uma faixa ininterrupta de material suberizado e lignificado — a banda de Caspary — que brilha com um âmbar-laranja profundo, como cobre aquecido ou resina antiga iluminada de dentro, uma barreira apoplástica que força toda a água e os iões absorvidos do solo a atravessar o interior vivo das células endodérmicas antes de alcançar o estelo, garantindo ao vegetal um controlo rigoroso sobre o que entra no sistema vascular. Atrás do observador, o parênquima cortical abre-se numa arquitectura mais frouxa e arejada — células de contorno arredondado, vacúolos grandes e quase transparentes, espaços intercelulares que se enchem de ar húmido numa penumbra azul-prateada e difusa. À frente, o estelo condensa-se num núcleo denso de células procambiais, mais pequenas e empacotadas com citoplasma saturado de púrpura e índigo, os seus núcleos pálidos destacando-se como pedras num colar. A faixa de Caspary arde silenciosa entre estes dois territórios de temperatura cromática oposta — o âmbar quente contra o violeta lunar —, lembrando uma arquitectura viva de separação absoluta, bela e sem hesitação.

Diante de você, um mundo de âmbar construído em fortalezas orgânicas se estende em todas as direções — a exina do grão de pólen de rosa, composta de esporoporenina, um dos biopolímeros mais resistentes já produzidos pela vida, capaz de sobreviver intacto por milhões de anos em sedimentos e turfeiras. As cristas arredondadas da malha reticulada formam células poligonais ligeiramente irregulares, cada uma com cerca de 2,5 micrômetros de largura, seus lúmens afundando em bacias de sombra ocre com uma profundidade igual à sua largura, criando uma arquitetura cujo ritmo geométrico reflete as tensões físicas e químicas que governaram a deposição da esporoporenina durante o desenvolvimento do micrósporo. Uma iluminação rasante de baixo ângulo atravessa o horizonte e esculpe cada crista em relevos dramáticos de ouro brilhante e sombra queimada, enquanto o próprio material emite uma autofluorescência quente e difusa — não uma luz externa, mas uma luminescência interior acumulada no polímero. Ao fundo, o colpo rasga o mundo como um vale tectônico: as paredes da exina afeiçoam-se ali até quase a transparência, as últimas cristas atenuadas desfazendo-se em esporões costeiros onde a superfície esférica do grão mergulha nessa abertura funcional, a única zona permeável num território que de resto se fecha sobre si mesmo como uma fortaleza fossilizada do mundo vegetal.
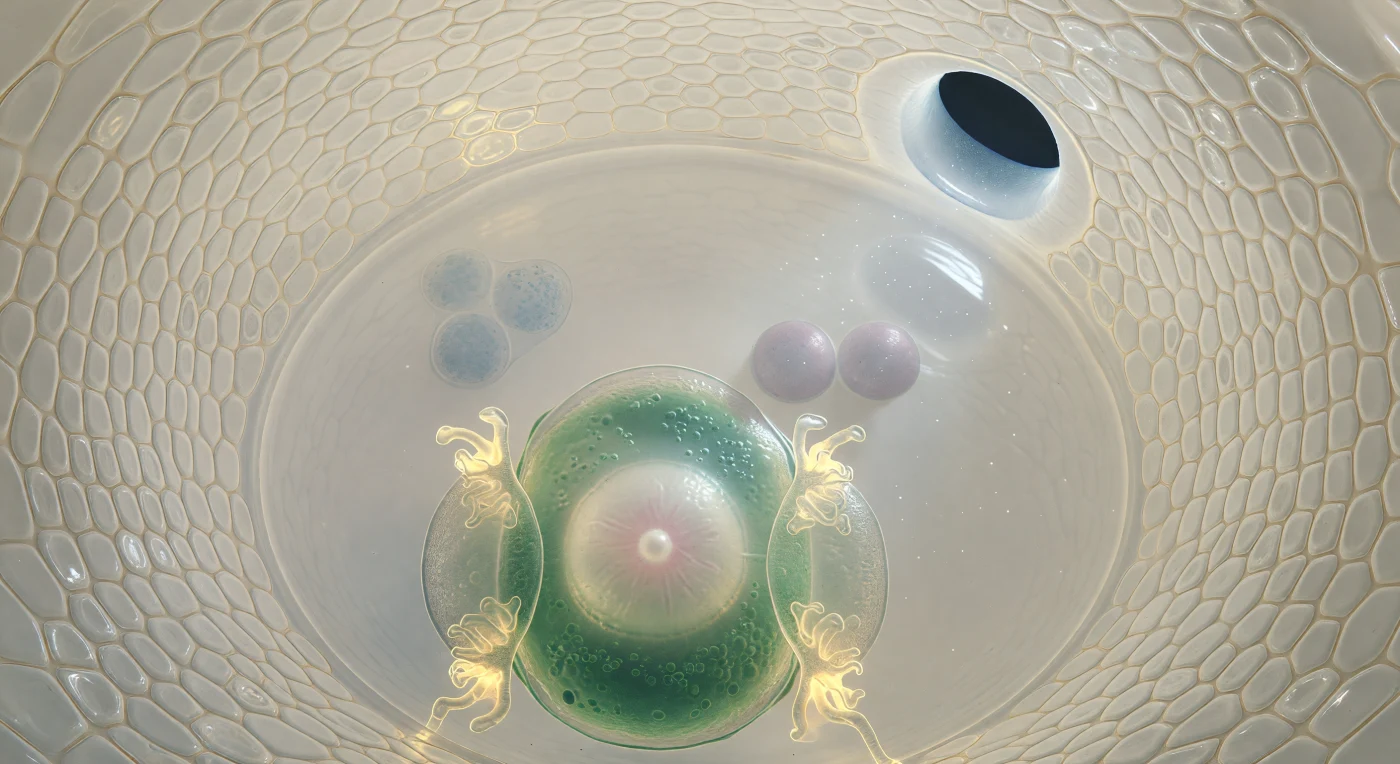
Suspenso no interior do saco embrionário de um óvulo quase maduro, o observador encontra-se a cerca de quarenta micrómetros da superfície convexa da célula-ovo, uma massa periforme de citoplasma de tonalidade verde-jade cuja periferia rica em organelos envolve um núcleo grande e luminoso, alvo como alabastro, com um nucléolo denso que capta a luz como uma pérola em água parada. As duas camadas de integumento arqueiam-se em volta como as paredes de uma gruta de marfim, as suas células achatadas e comprimidas unidas por tênues costuras âmbar, enquanto pelo micrópilar — uma passagem oval de apenas quinze micrómetros na extremidade superior direita — entra um fio de luz exterior que ilumina partículas em suspensão no fluido apoplástico. Flanqueando a célula-ovo, as duas sinérgidas pressionam o seu corpo mais refractivo contra ela, as suas invaginações filiformes na ponta micropilar a brilhar com uma fluorescência âmbar-dourada, rendas membranares que dispersam a luz em filamentos ramificados como brasas. Para lá da célula-ovo, a célula central abre-se numa câmara de citoplasma tão transparente que parece uma sala cheia de água, no centro da qual dois núcleos polares flutuam como luas lilás-cinzentas com um ligeiro iridescente, e no polo calazal mais distante três células antipodais agrupam-se em sombra difusa, azul-acinzentadas como pedras vistas através de água funda, marcando o limite mais remoto desta câmara viva.

Suspenso na fronteira entre dois mundos opostos, o olhar percorre simultaneamente a vastidão dos lumens vasculares do lenho inicial — túneis circulares de 250 µm de diâmetro que se abrem como grutas submarinas cortadas em vidro âmbar — e a densidade quase mineral do lenho tardio à direita, onde paredes de fibras libriformes espessadas reduzem os lumens a fendas de 15 µm, mal visíveis como fissuras em obsidiana. Esta fronteira abrupta, mais parecida com uma falésia do que com uma transição gradual, regista em madeira de carvalho a memória de uma estação inteira: o lenho inicial formado em primavera pela divisão acelerada do câmbio vascular, otimizado para o transporte de água em massa; o lenho tardio construído no verão e outono, enriquecido em celulose cristalina orientada em hélice nas paredes secundárias S2, conferindo resistência mecânica ao tronco. Sob luz polarizada, essa arquitetura molecular torna-se visível como paisagem: as microfibrilas de celulose, com ângulos de deposição distintos entre as camadas S1, S2 e S3, desviam os planos de vibração da luz produzindo cores de interferência em azul-cobalto elétrico e ouro brunido que transformam a secção num mosaico de esmalte bizantino. As fitas de parênquima radial — células em tijolo cor de mel, de paredes primárias finas, dois a quatro elementos de largura — atravessam toda esta fronteira horizontalmente como estratos geológicos, indiferentes à divisão funcional, lembrando que a comunicação lateral entre xilema e câmbio é contínua, independentemente da estação que a madeira ao redor registou.
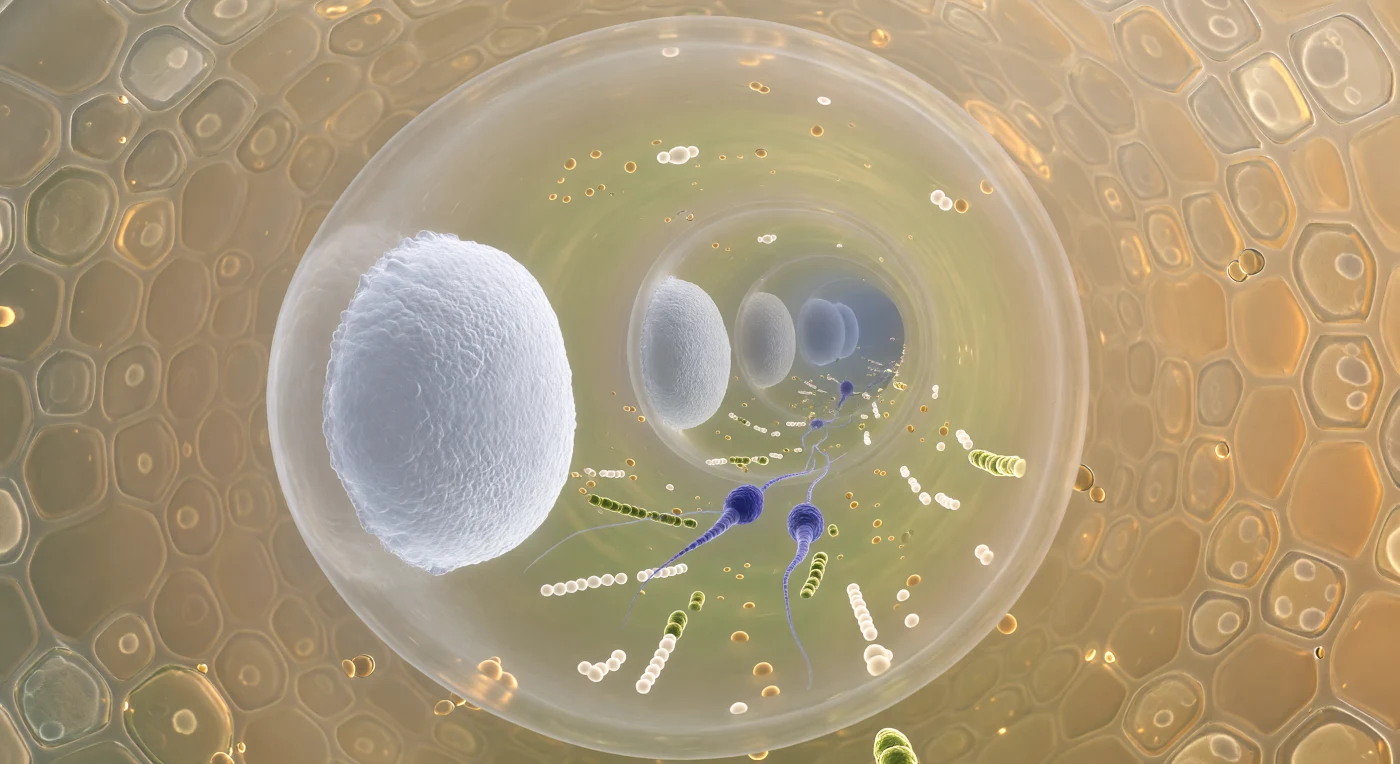
Entre duas sólidas comportas de calose que selam o corredor como paredes translúcidas de porcelana fosforescente, o interior do tubo polínico revela-se um canal vivo com cerca de doze micrómetros de diâmetro, tão estreito que a curvatura da parede celulósica se fecha suavemente à nossa volta como o interior de um tubo de vidro âmbar. A corrente citoplasmática transporta uma massa densa de organelos — gotículas lipídicas cor de âmbar, mitocôndrias com um brilho amarelo-esverdeado nas cristas, vesículas agrupadas em cadeias — e, no meio desse tráfego, dois gâmetas fusiformes de cor azul-violeta intenso avançam de forma ordenada, os seus envólucros nucleares distendidos em pontas e a cromatina tão condensada que parecem inclusões minerais flutuando em maré quente. Para além da parede do tubo, o tecido transmissor estilar pressiona de perto — células secretoras em mosaico dourado-pálido, os seus vacúolos cheios de uma secreção polissacarídica refratante que banha o exterior com uma neblina âmbar viscosa e adesiva, filtrando a luz em tons de laranja-dourado que fazem os septos de calose parecerem velas acesas num corredor invisível. Esta procissão biológica — urgente, pressurizada, geometricamente precisa — representa um dos transportes celulares mais rápidos e dirigidos do mundo vegetal, garantindo que os gâmetas masculinos percorram dezenas de milímetros até à oosfera no interior do óvulo, sem jamais deixarem o interior do tubo que os criou.