Au fond de ce couloir lignifié d'ambre brûlé, vous traversez l'intérieur d'un vaisseau du xylème d'angiosperme mature, une conduite creuse d'environ 120 micromètres de diamètre dont les parois sont tapissées de centaines de ponctuations aréolées — chacune un disque en creux de six micromètres, bordé d'un anneau de cellulose imprégnée de lignine, et tendu d'une membrane fantôme translucide qui rappelle la pression différentielle qu'elle régulait naguère entre deux vaisseaux adjacents. Cette architecture de galerie répétitive n'est pas un ornement : elle est la solution évolutive des plantes vasculaires pour maintenir la continuité du flux hydrique sous tension négative, permettant à une colonne d'eau cohésive de s'élever de la racine jusqu'à la canopée sans se rompre, grâce à la cohésion des molécules d'eau et à l'adhésion aux parois hydrophiles. Pourtant, au fond du lumen parfaitement vide — sous vide hydraulique, sec jusqu'à l'absolu — une bulle d'air embolisante occupe toute la largeur du conduit comme un miroir de mercure convexe, sa surface réfléchissant en miniature distordue la galerie entière de ponctuations derrière vous. Cet embolie représente une rupture catastrophique du continuum sol-plante-atmosphère : le vaisseau est mort au transport de l'eau, condamné jusqu'à ce que la refilling nocturne ou la croissance de nouveaux éléments vasculaires rétablisse le circuit. La beauté froide de cette sphère argentée suspendue entre cohésion et cavitation dit tout de la fragilité exquise du système vasculaire végétal.

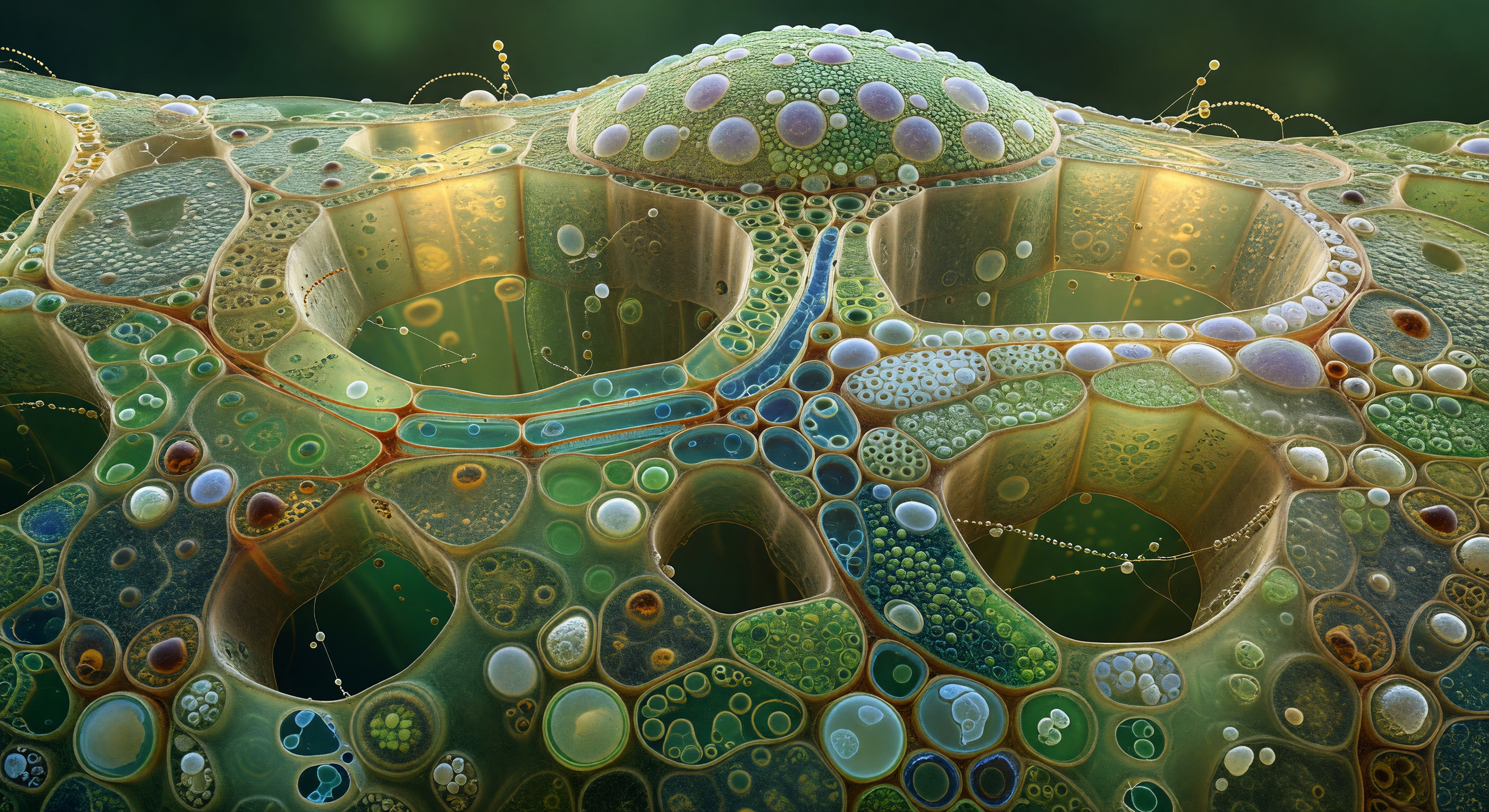
Levant les yeux depuis le plancher de la feuille, on se retrouve au pied d'une forêt de colonnes cylindriques géantes, leurs parois de verre ambré incrustées de centaines de chloroplastes biconvexes pressés côte à côte comme des tuiles d'émeraude vivantes, chacun irradiant une luminescence froide et verte qui se superpose aux autres en couches de jade de plus en plus profondes vers la base. Ces cellules du mésophylle palissadique, hautes d'une soixantaine de micromètres, sont spécialisées pour capturer la lumière avec une efficacité maximale : leur orientation verticale et la densité de leurs chloroplastes — chacun contenant des grana empilés de membranes thylakoïdiennes gorgées de chlorophylle — transforment chaque paroi en un panneau photovoltaïque organique opérant à l'échelle du micromètre. Depuis le plafond éloigné, la lumière descend en rideaux quasi parallèles au travers de la cuticule et des cellules épidermiques, qui diffusent les photons comme du verre dépoli, leur ôtant tout angle dur avant qu'ils ne baignent les colonnes d'une clarté égale et laiteuse. Entre les colonnes, les espaces intercellulaires s'ouvrent en crevasses presque noires, véritables allées menant aux cavernes du mésophylle spongieux, où circulent vapeur d'eau, dioxyde de carbone et oxygène — les matières premières et les produits d'une chimie solaire qui s'accomplit, silencieusement et en tous sens, à chaque fraction de seconde.
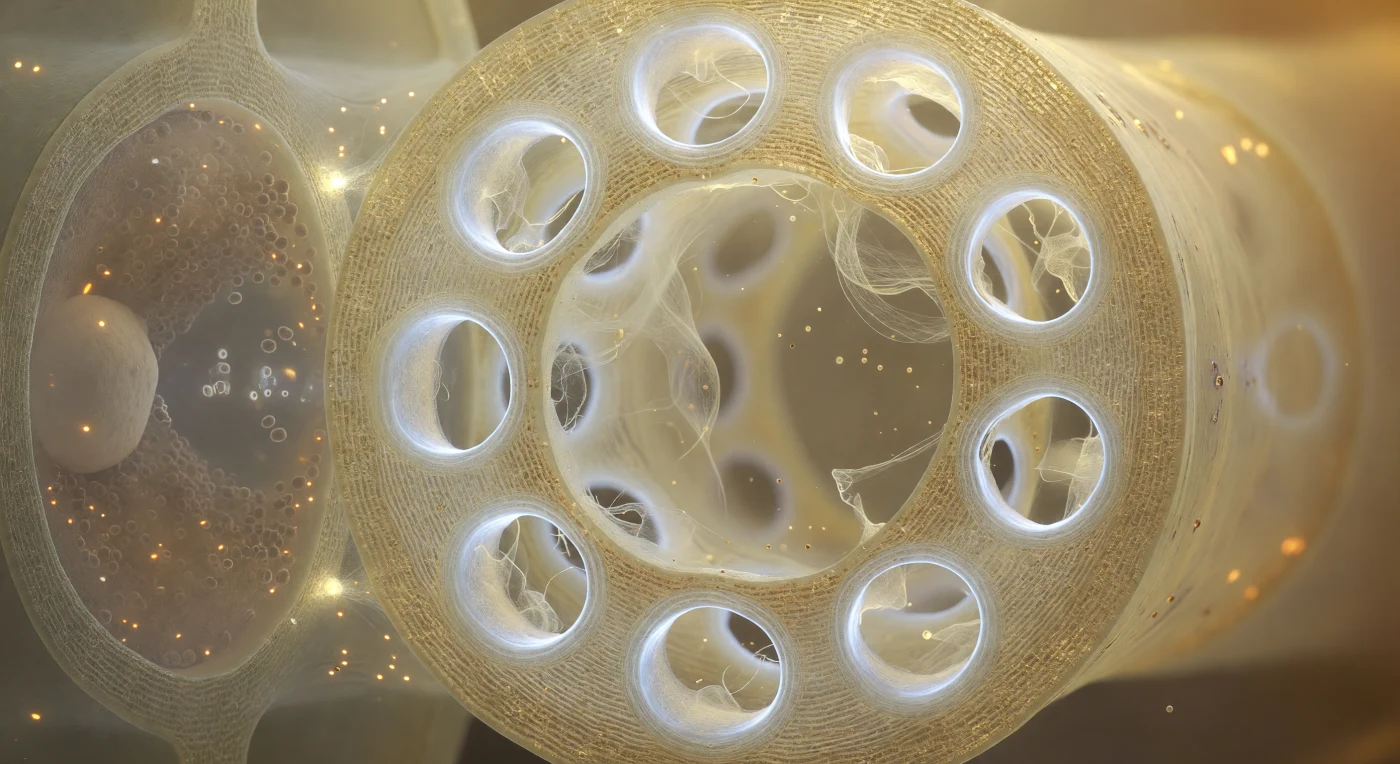
Vous êtes immergé à l'intérieur d'un élément de tube criblé, face à la plaque criblée qui ferme le compartiment comme un vitrail de cathédrale à l'échelle cellulaire : la paroi de cellulose, translucide comme de l'ambre pâle, est percée de douze pores disposés en constellation irrégulière, chacun cerclé d'un épais collier de callose aux reflets blanc laiteux légèrement bleutés, biopolymère déposé en lamelles concentriques qui régulent le flux en réduisant l'ouverture de chaque pore. Au travers de ces portails lumineux, des filaments de protéine P dérivent en un voile iridescent couleur de crème diluée, mus par le courant quasi imperceptible de la sève élaborée qui pulse d'un compartiment à l'autre à environ un mètre par heure, selon le mécanisme de flux de masse généré par les différences de pression osmotique entre les feuilles sources et les organes puits. Sur le côté gauche, séparée seulement par une mince paroi commune criblée de plasmodesmes — minuscules ponctuations lumineuses à peine résolubles —, la cellule compagne presse contre le tube criblé comme une pièce sombre contre une lanterne : son cytoplasme dense de mitochondries, de ribosomes et de réticulum endoplasmique renforce métaboliquement l'élément criblé voisin, qui a lui-même perdu son noyau à maturité. L'architecture de ce duo cellulaire, entièrement dévolue au transport des photoassimilats sur des distances pouvant atteindre plusieurs mètres, révèle une intimité biologique saisissante où la lumière chaude du lumen s'oppose à l'obscurité dense et grouillante de la cellule compagne.
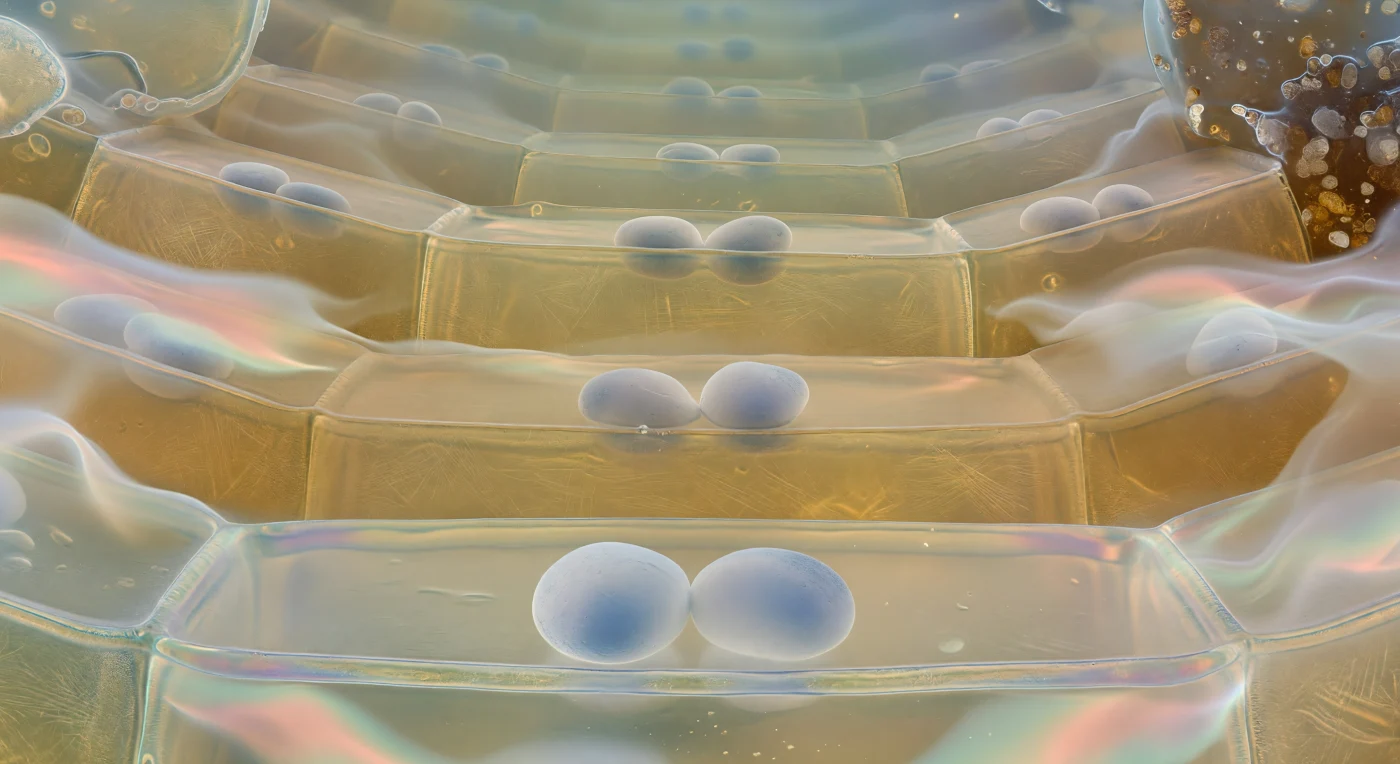
Suspendu juste au-dessus du plancher d'une cellule de columelle, le regard porté vers le haut traverse une cathédrale de verre ambré vivant : les parois en cellulose, épaisses comme des dalles de résine mielleuse, s'élèvent en plans doucement courbés dont la trame de microfibrilles se devine en filigrane, et à travers les plafonds semi-transparents se succèdent tier après tier d'autres chambres identiques, chacune abritant ses propres statolithes amyloplastiques — ces ovoïdes denses et mats, blancs virant au gris-bleu là où la lumière les traverse — qui reposent avec une immobilité absolue contre la membrane plasmique tendue comme une peau de tambour irisée aux reflets violets, dorés et bleu-glace. Ces corpuscules d'amidon sont les sédimenteurs de gravité de la plante : en se déposant systématiquement vers le bas de chaque cellule, ils renseignent le méristème racinaire sur l'orientation gravitationnelle, déclenchant via la redistribution d'auxine la courbure différentielle qui guide la croissance du pivot dans le sol. À la périphérie de cette architecture, les cellules bordières se dissolvent dans une brume de mucilage hydrogel qui réfracte la lumière transmise en rubans spectraux de rose saumon et d'aigue-marine pâle, pendant qu'au-delà de cette auréole biochimique scintillante s'entrevoit l'obscurité comprimée du sol — grains de quartz et argiles loam couverts d'un film humide luisant —, rappelant avec une force tranquille l'immensité de la pression hydraulique que ce minuscule espace cellulaire repousse en permanence.

Sous la lumière polarisée croisée, un drus de calcium oxalate se dresse au centre du champ visuel comme une rose gothique minéralisée : quarante facettes cristallines rayonnent d'un noyau central dense, chacune brûlant en cobalt profond, en ambre brûlé ou en or pur selon l'angle de rétardation biréfringente, le tout suspendu dans un vide absolu que le milieu aqueux de la vacuole a cédé à une obscurité parfaite. Cette structure d'environ cinquante-cinq micromètres est un précipité minéral figé à l'intérieur d'une cellule parenchymateuse vivante, résidu cristallographique des sursaturations en ions calcium et oxalate que la plante séquestre pour réguler sa chimie interne ou se défendre contre les herbivores. Chaque facette porte de fines terrasses de croissance — marches invisibles à toute autre échelle — qui modulen la couleur d'interférence ligne par ligne, transformant la topographie cristalline en un atlas spectral gravé dans le minéral biologique. Au cœur du drus, là où les cristallites individuels s'interpénètrent, les couleurs se fracturent en magenta, vert pâle et or chaotique, témoignant du moment de nucléation primordiale autour duquel la précipitation s'est propagée avec une inévitabilité cristallographique. À la toute périphérie du champ, la paroi cellulaire de cellulose trace un cadre doré et discret — rappel silencieux que cette cathédrale minérale resplendit enfermée dans le vivant.

Suspendu à cent-cinquante micromètres au-dessus du dôme, le regard plonge sur une voûte légèrement convexe dont la courbure semble presque inexistante jusqu'à ce que les deux primordiums foliaires dorés aux flancs révèlent, par contraste, la logique sphérique de l'ensemble. La surface se déploie en un honeycomb d'ivoire et de jade pâle, chaque cellule isodiamétrique d'une douzaine de micromètres bordée par des parois cellulosiques si minces qu'elles fonctionnent comme des panneaux de verre dépoli, laissant filtrer une luminosité douce et sourceless qui efface toute ombre franche. Au centre de chaque facette hexagonale, un noyau ovale se distingue comme un galet sombre pressé contre du verre givré — signe de cellules à peine vacuolées, denses en ribosomes et en machinerie mitotique, qui se divisent activement pour générer l'intégralité des organes à venir selon un programme moléculaire orchestré par les gènes WUSCHEL et CLAVATA. Aux marges du dôme, là où les primordia s'élèvent en crêtes couleur ambre, la géométrie régulière du méristème cède progressivement à des cellules qui s'allongent et épaississent leurs parois, traduisant en architecture cellulaire visible le moment précis où une identité organique commence à s'écrire dans la matière vivante.

Depuis le fond de la cavité sous-stomatique, le regard s'élève à travers les brumes de vapeur d'eau vers l'ouverture au-dessus, où deux cellules de garde arquées encadrent une fente lumineuse de sept micromètres — une porte cathedral vers l'atmosphère libre, baignée d'une lumière froide et blanche qui tranche sur le vert électrique des cellules vivantes. Chaque cellule de garde rayonne d'une intensité presque autonome : douze chloroplastes, chacun large de quatre à six micromètres, se pressent en file serrée le long des parois internes épaissies, leurs membranes gorgées de grana absorbant et renvoyant la lumière comme des lanternes de jade humide. Ces parois internes, lamellées d'une ivoire pâle de microfibrilles de cellulose densément tressées, résistent à la pression osmotique considérable du contenu vacuolaire — c'est cette asymétrie structurale précise, et non un mécanisme musculaire, qui maintient l'ouverture béante plutôt qu'effondrée. Les lèvres cuticulaires surplombent le pore comme les bords d'une citerne de cristal, effilées jusqu'à une épaisseur quasi moléculaire où elles diffractent la lumière en un liseré iridescent de or pâle et de violet, tandis qu'en dessous, dans le labyrinthe spongieux du mésophylle, les surfaces cellulaires luisent de films aqueux condensés qui diffusent la lumière en une brume biologique vert-lumineuse, estompant les profondeurs en une clarté vivante et indéfinie.

Vous vous tenez suspendu dans un couloir de membranes à peine visibles, prisonnier d'un instant de printemps figé à l'intérieur du cambium vasculaire, cette frontière vivante qui gouverne en silence l'épaississement d'un arbre. Devant vous, les initiales fusiformes s'étendent comme des nefs de cathédrale translucides, leurs parois si minces qu'elles semblent respirer — des voiles de soie mouillée tendus entre deux empires en train de se solidifier, leur cytoplasme d'un jaune paille lumineux vous enveloppant comme l'intérieur d'une lanterne habitée. À gauche, le monde du xylème s'arme sans bruit : les trachéides récemment formées déposent leurs parois secondaires en anneaux concentriques d'ocre et de rouille, la lignine s'infiltrant dans la cellulose pour créer des tubes morts d'une précision architecturale absolue, cette nécropole ingénieure qui conduira l'eau des racines jusqu'aux feuilles à des vitesses impossibles. À droite, le phloème se fait plus doux, presque liquide : les éléments criblés abandonnent volontairement leurs noyaux et s'ouvrent à la circulation, leurs plaques criblées se perforant sous des halos de callose opalescents, tout un réseau en train de se donner au flux de sève élaborée. Le cambium lui-même — cette couche à peine épaisse d'une cellule, invisible à l'œil nu dans l'écorce — est la membrane génératrice qui fabrique en permanence l'arbre vers l'intérieur et vers l'extérieur, une frontière vivante et presque transparente coincée entre deux mondes qui durcissent.

Vous planez à la surface d'une feuille de tomate, mais le paysage qui s'étend devant vous n'a rien d'ordinaire : une tour cellulaire à six étages s'élève depuis l'épiderme en contrebas, ses assises de cellules diminuant légèrement de diamètre vers le haut, leurs parois communes épaissies de pectine visibles comme des lignes sombres et précises, jusqu'à la couronne de quatre cellules sécrétoires aplaties qui coiffe l'ensemble comme un diadème translucide. Au-dessus de cette couronne, la cuticule a été repoussée vers l'extérieur par la pression des huiles essentielles accumulées en dessous, formant une ampoule tendue et ambrée dont la surface bombée capte la lumière oblique en un reflet liquide et lenticulaire — à l'intérieur, de fines variations de densité trahissent la superposition de composés terpénoïdes distincts, secrétés successivement et retenus sous une tension croissante, à l'instant précis avant que la cuticule ne cède. Tout autour, la surface épidermique s'étend comme une plaine tectonique aux reliefs délicats : les cellules à parois sinueuses et interdigitées des dicotylédones forment un carrelage légèrement bombé de territoires translucides, recouvert d'une forêt éparse de micro-cristaux de cire épicuticulaire en bâtonnets et en plaques qui projettent de petites ombres dures sur le fond mat et bleuté — rendant l'ampoule d'huile au sommet du trichome d'autant plus lumineuse, seul objet brillant dans ce paysage de parois et de cire.

Au fil d'un corridor vivant, le regard se perd dans une enfilade de chambres translucides qui s'étire vers l'infini aqueux, chaque cellule dressée sur près de deux cent cinquante micromètres comme une cathédrale de verre dépoli dont les parois de cellulose transmettent la lumière à la manière d'une vitre givrée teintée de bleu-blanc. L'intérieur de chaque chambre est presque entièrement occupé par une vacuole centrale colossale, volume d'eau pure à la limpidité optique parfaite, maintenu sous une pression de turgescence si intense que les parois se bombent légèrement vers l'extérieur, chaque angle arrondi témoignant silencieusement d'une force hydraulique immense suspendue dans un instant immobile. Le cytoplasme vivant, réduit à une mince pellicule pariétale d'un vert pâle à peine perceptible, se plaque contre la face interne de chaque paroi comme un vernis biologique résiduel, interrompu çà et là par un noyau — sphère dense d'une dizaine de micromètres baignant dans cette marge ténue comme une lanterne à la lisière d'un océan, révélant par contraste le vertige de l'espace qui l'entoure. Loin devant, au terme de dizaines de cellules empilées dont les parois communes forment une grille de septa lumineux qui s'effacent progressivement dans la brume céruléenne, le stèle central se profile comme une colonne indigo sombre, tissu vasculaire dont les parois lignifiées absorbent la lumière transmise plutôt qu'elles ne la laissent passer, axe silencieux autour duquel s'organise toute l'architecture de cette élongation racinaire captée entre deux respirations.

Au seuil entre le monde des vivants et celui de la matière inerte, vous vous trouvez à l'intérieur même de l'écorce d'un arbre, immergé dans une architecture cellulaire d'une rigueur presque minérale : devant vous s'étendent, colonne après colonne, les cellules mortes du liège, leurs parois subérisées rayonnant d'un ambre chaud de cognac vieilli, leurs lumens scellés en vides absolus — chaque chambre une salle close depuis longtemps, imperméable à l'eau, aux gaz, à toute chimie du vivant, grâce aux lamelles de subérine qui imprègnent progressivement les parois au fil des saisons, les plus anciennes vers la surface extérieure virant à un brun-sienna plus sombre et plus dense. Cette forteresse de liège — le phellem — est le produit du phelloderme vers l'extérieur, fabriqué par une assise génératrice unique, le phelloderme, dont la ligne pâle et translucide surgit soudain comme une membrane vivante tendue à travers tout le champ visuel, ses cellules fines et lumineuses contrastant de façon presque violente avec l'ambre lourd qui les précède. Au-delà, le phelloderme parenchymateux s'entrouvre en cellules plus rondes, teintées du vert diffus de chloroplastes endormis, respirant encore une chaleur métabolique lente. Et là, sur le côté, là où la géométrie serrée s'effondre en un lenticelle — zone pâle de cellules incomplètement subérisées, aux espaces intercellulaires ouverts — la lumière s'engouffre obliquement en éclats d'or et de crème, rappelant que cette muraille hermétique n'est pas absolue : elle respire, par endroits, à travers ses propres failles consenties.

Vous flottez en suspension dans l'air chaud et saturé d'humidité d'un canal aérenchymateux de pétiole de nénuphar, un tunnel organique de huit cents micromètres de diamètre dont la paroi incurvée se déploie tout autour de vous comme la coque intérieure d'un sous-marin vivant — une mosaïque continue de cellules hexagonales et polygonales gorgées de chlorophylle, chacune large de deux cents micromètres, leurs parois communes minces comme du parchemin mouillé et légèrement ambrées aux jonctions où la cellulose s'épaissit. Ce tissu de chlorenchyme qui tapisse le canal remplit une double fonction : il capte la lumière filtrée jusque dans le pétiole immergé et approvisionne en oxygène photosynthétique le réseau de lacunes d'air qui parcourt toute la plante aquatique, permettant aux feuilles de respirer leurs racines enfouies dans la vase anoxique. Directement devant vous, un diaphragme stellaire s'étend d'une paroi à l'autre comme un médaillon de dentelle en suspension dans le vide — une couronne de cellules en bras allongés rayonnant depuis un moyeu central, leurs extrémités translucides ne rejoignant jamais tout à fait la paroi du canal, laissant entre elles des vides polygonaux ouverts assez larges pour y passer. Une lumière diffuse venant du fond du tunnel traverse cette cloison lacunaire et la transforme en silhouette rayonnante, chaque bras cerclé d'un halo doré-vert là où la membrane cellulaire réfracte la lueur transmise, tandis que de fins films d'eau irisés tremblent dans certaines des ouvertures, projetant de brèves interférences de cuivre et de violet sur les parois émeraude du canal qui s'enfonce, dans les deux directions, vers une obscurité atmosphérique douce et sans fond.
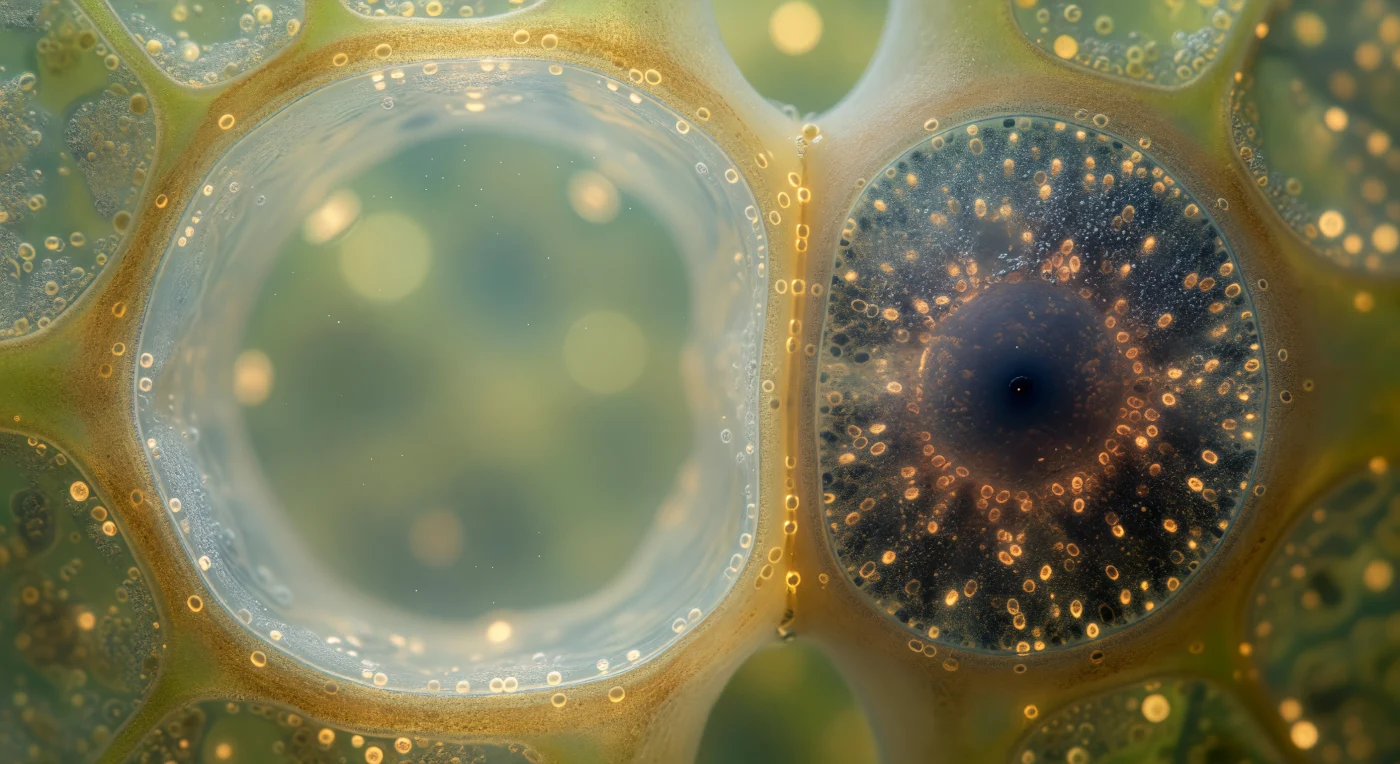
Dans cette coupe transversale du faisceau libérien, deux mondes cellulaires radicalement opposés s'affrontent dans un même champ de vision : à gauche, l'élément de tube criblé s'étend comme une cathédrale vide, sa lumière aqueuse d'un ambre pâle presque translucide baignant dans un silence sucré, la couche pariétale cytoplasmique réduite à un film nacré à peine visible le long des parois, tandis qu'à droite la cellule compagne — un quart de sa taille — concentre dans son volume restreint une obscurité dense et fiévreuse, bleu-noir et café, saturée de mitochondries ovales luisant faiblement à leurs membranes et d'un tapis de ribosomes donnant au cytoplasme la texture d'un sable volcanique humide. Ce contraste entre la chambre lumineuse et patiente du tube criblé et la forge métabolique bondée de sa cellule compagne traduit une division du travail d'une précision absolue : dépourvus de noyau et de la plupart de leurs organites à maturité, les tubes criblés abandonnent leur autonomie cellulaire et délèguent à la cellule compagne l'intégralité de leur entretien enzymatique et énergétique, les deux êtres reliés par des plasmodesmes spécialisés — visibles comme une constellation de perforations dorées ponctuant la paroi commune — qui maintiennent entre eux un dialogue moléculaire ininterrompu. C'est par ce couloir vasculaire, à la frontière de la lumière et de l'ombre, que la plante entière achemine ses sucres folaires vers ses racines, à environ un mètre par heure, dans un flux invisible mais vital.

Au seuil entre deux territoires biologiques, vous êtes suspendu dans la racine d'une jeune plante, face à une architecture vivante d'une précision absolue : les cellules endodermiques s'organisent en colonnade prismatique, leurs parois translucides d'un ivoire verdâtre encadrant un cytoplasme lentement animé, tandis qu'une ceinture de subérine et de lignine — la bande de Caspari — court sans interruption d'une paroi radiale à l'autre, formant un collier de feu ambré qui ceint le stèle tout entier. Cette barrière chimique n'est pas une simple membrane : c'est un joint moléculaire qui force chaque ion, chaque molécule d'eau, à traverser le cytoplasme endodermique plutôt que de circuler librement dans les parois, conférant à la plante un contrôle sélectif absolu sur ce qui pénètre dans son système vasculaire. Derrière vous, le cortex s'ouvre en une architecture lâche et aérée, pâle comme du verre dépoli, ses grands espaces intercellulaires baignés d'une brume bleu-blanc ; devant, le stèle se resserre en une masse dense d'indigo et de violet, les cellules procambiales tassées autour de leur futur xylème et phloème comme un joyau comprimé sous pression. Entre ces deux mondes — l'un diffus et perméable, l'autre ordonné et turgescent — la bande de Caspari brûle avec la constance d'un joint de forge, frontière lumineuse et infranchissable au cœur du végétal.

Vous vous tenez à la surface d'un monde entièrement construit en remparts d'ambre, un réseau continu de crêtes de sporopollenine s'élevant à hauteur de regard pour former une mosaïque de cellules polygonales irrégulières, chacune large de quelques micromètres, leurs planchers s'enfonçant dans des bassins teintés d'ocre que la lumière rasante ne parvient qu'à effleurer. Cette architecture réticulate n'est pas une décoration mais une armure biochimique — la sporopollenine, l'un des polymères organiques les plus résistants du vivant, capable de traverser des millions d'années fossilisé dans des sédiments sans se dégrader, protégeant le contenu gamétique du grain contre la dessiccation, les UV et les agents oxydants. À mi-distance, le terrain se dérobe le long d'une vallée pâle et translucide — le colpus — là où l'exine s'amincit pour permettre l'hydratation et l'émission du tube pollinique lors de la germination, zone de fragilité calculée au sein d'une forteresse par ailleurs indestructible. La lumière elle-même semble emprisonnée dans la matière : chaque crête émet une autofluorescence dorée douce, non pas un éclat vif mais la restitution lente d'une énergie absorbée, tandis que les ombres s'accumulent dans les lumina comme une géologie de l'obscurité. Ce paysage microscopique, silencieux et géométriquement infini, est le premier décor que rencontrera tout autre organisme cherchant à féconder cette fleur.
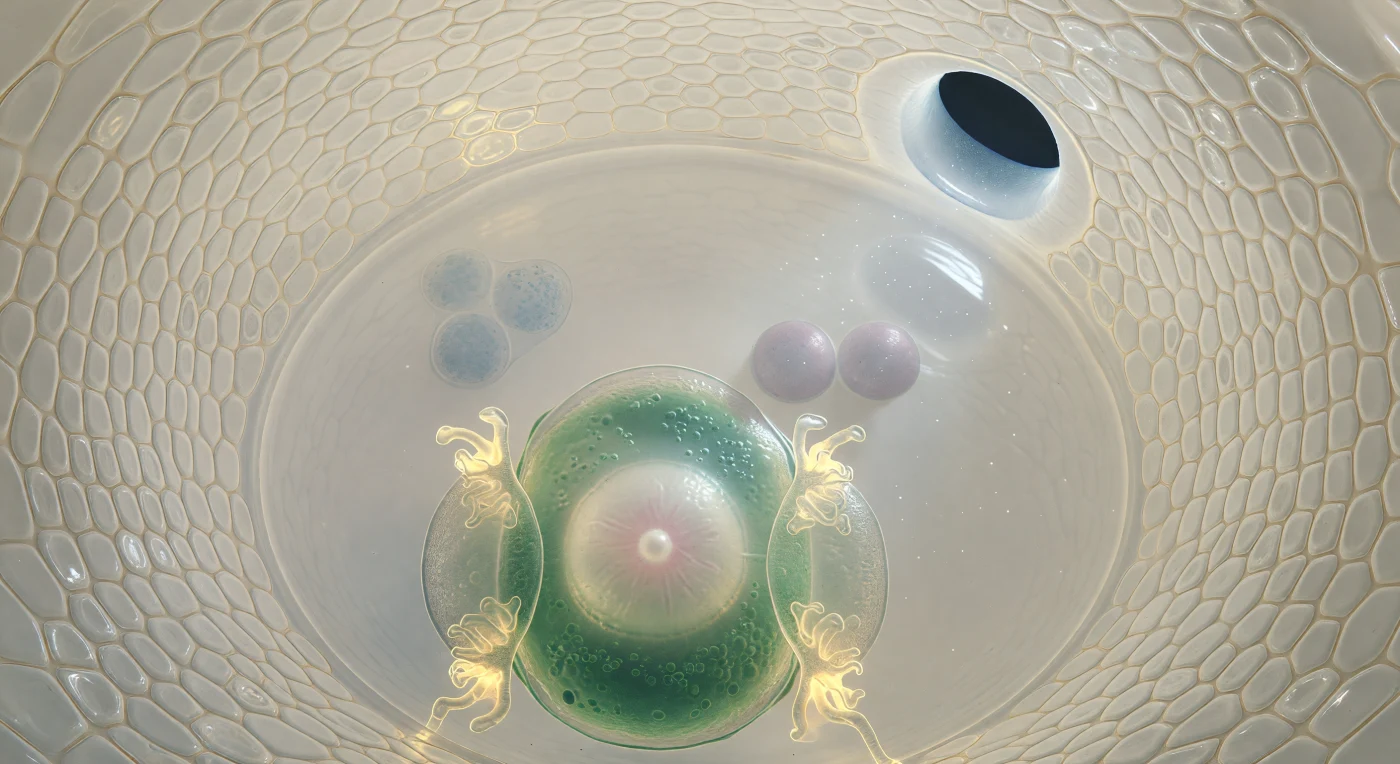
Au seuil de ce monde clos, le regard plonge dans l'intérieur de la sac embryonnaire comme dans la nef d'une cathédrale vivante : les deux couches d'intéguments s'incurvent tout autour, mosaïques de cellules ivoires pressées les unes contre les autres, leurs parois communes tracées en fines lignes ambrées dans une translucidité douce et sourceless, tandis qu'au coin supérieur droit le micropyle s'ouvre en oval sombre de quinze micromètres à peine, laissant filtrer un mince faisceau de lumière extérieure qui illumine en suspension les particules du fluide apoplasmique. Au premier plan, la cellule-œuf occupe l'espace comme une masse en forme de poire d'un vert aqueux profond à sa périphérie chargée d'organites, son grand noyau — sphère d'albâtre veiné de rose pâle, avec son nucléole nacré — flottant au pôle micropylaire, flanqué de deux cellules synergides dont les ingrowths filiforme brillent d'un or ambré concentré, lacis de membranes rayonnant comme des braises. Derrière elles, la cellule centrale s'ouvre en une chambre d'une clarté cristalline presque irréelle, où deux noyaux polaires couleur de lilas gris dérivent côte à côte, effleurés par un reflet irisé du faisceau micropylaire — deux lunes suspendues dans un océan immobile. Loin, au pôle chalazal, trois cellules antipodales se devinent en amas granuleux et bleuté, comme des pierres entrevues à travers une eau profonde et parfaitement limpide, aux confins d'un univers entier contenu dans une seule graine.

Vous flottez au seuil de deux continents cellulaires, suspendu à la frontière la plus abrupte que le règne végétal puisse tracer en une seule saison de croissance. À gauche, les vaisseaux du bois initial s'ouvrent comme des cavernes sous-marines taillées dans l'ambre — leurs lumières circulaires de 250 µm de diamètre, absolument noires et sans fond, cerclées d'un halo ambré là où les chambres des ponctuations aréolées captent la lumière polarisée en minces reliefs d'architecture pétrifiée. La frontière arrive sans transition, telle une falaise : à droite, le bois final oppose une matière quasi solide, les lumières réduites à de fines fentes de 15 µm luisant comme des craquelures dans l'obsidienne, leurs parois secondaires de cellulose cristalline explosant sous la lumière polarisée en mosaïques de cobalt électrique et d'or byzantin — énergie mécanique comprimée dans la matière organique comme dans une roche métamorphique. Traversant les deux mondes à l'horizontale, les rubans de parenchyme radial découpent la frontière comme des strates géologiques courant à travers le grès : leurs cellules en briques couleur de miel pâle, aux parois primaires fines et aux intérieurs doucement lumineux, ignorent souverainement la division earlywood-latewood et rappellent que ce bois fut, une saison entière, un organisme vivant transportant sucres et signaux entre l'écorce et le cœur.
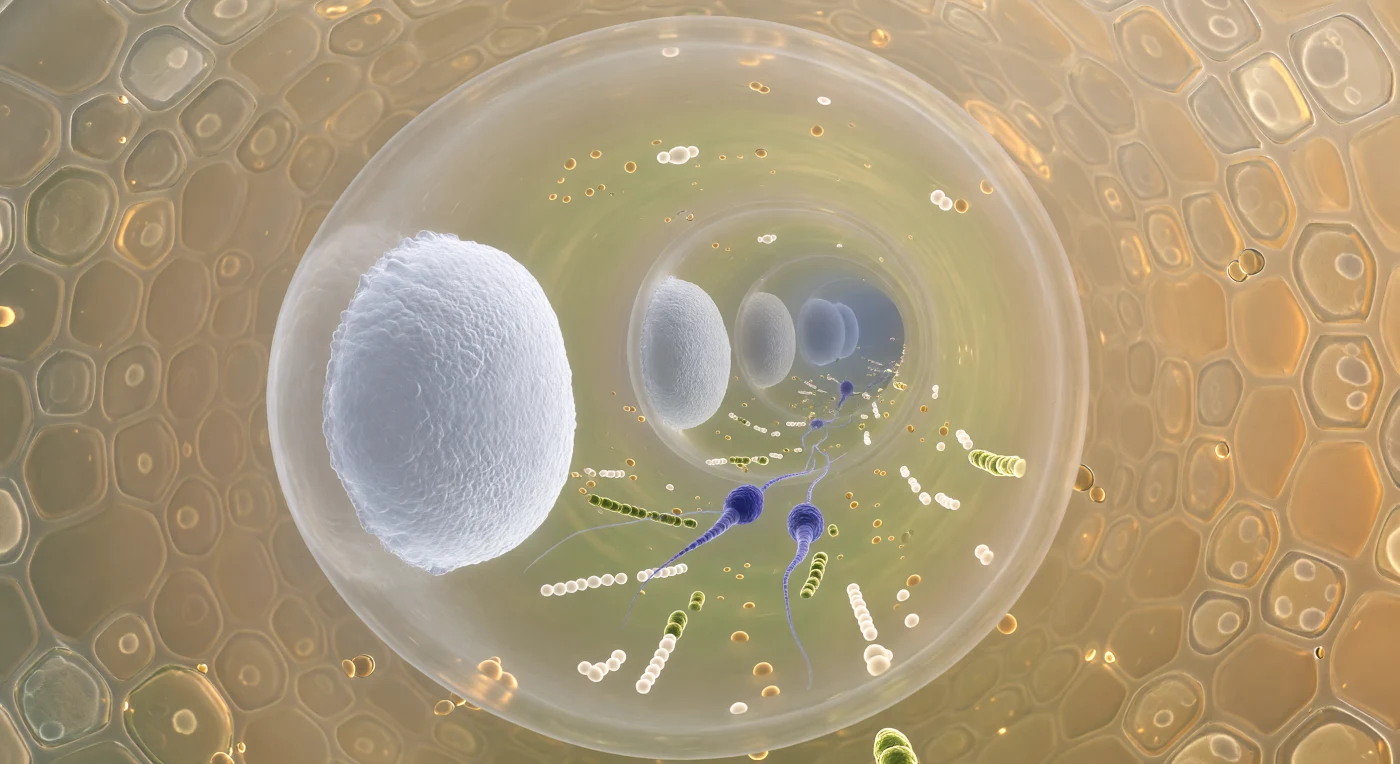
Dans ce voyage en vision tunnel à l'intérieur d'un tube pollinique vivant, le regard plonge en avant à travers un cylindre de douze micromètres à peine, dont la paroi translucide teintée d'ambre — une trame de cellulose renforcée de callose — laisse filtrer la lueur dorée diffuse du tissu transmetteur stylair qui presse de toutes parts, ses cellules sécrétoires polygonales noyées dans une matrice polysaccharidique visqueuse comme du miel chaud. Droit devant, à quelques dizaines de micromètres, une première cloison de callose se dresse comme un hublot givré et légèrement convexe, d'un blanc porcelaine presque phosphorescent au centre et vitreux sur ses bords, avant qu'un deuxième puis un troisième bouchon ne s'effacent en disques de plus en plus pâles et bleutés, formant une colonnade de septa lumineux qui converge vers la pointe de croissance invisible. Entre ces deux premières cloisons, le cytoplasme est traversé d'un flux dense d'organites — gouttelettes lipidiques ambre, mitochondries jaune-vert, vésicules en chapelets — au sein duquel deux cellules spermatiques fusiformes, d'un indigo-violet intense, sont portées sans effort, leur chromatine si condensée qu'elle paraît minérale, drapées de filaments de réticulum endoplasmique comme des rubans de soie dans le courant. Toute la géographie de ce corridor intérieur, plus étroit qu'un cheveu humain, vibre d'une urgence biologique silencieuse : celle d'un voyage à sens unique vers l'ovule.