Wetenschappelijke betrouwbaarheid: Hoog

Je staat in het holle binnenste van een volwassen xyleem-vat, een cilindrische gang van ongeveer 120 micrometer doorsnede waarvan de wanden zijn opgebouwd uit versteend ligno-cellulose materiaal dat baadt in een diffuus barnsteengeel licht. Over het hele cilindrische oppervlak zijn honderden behofde poriën aangebracht in een strak geometrisch patroon: cirkelvormige ondiepe holtes van zo'n zes micrometer breed, elk omrand door een licht verhoogd ligninelipje en afgedekt met een vliesachtig membraanspook dat de drukwisselingen tussen aangrenzende vaten reguleerde toen het vat nog actief water transporteerde. Dit vat is nu leeg en staat onder negatieve hydraulische spanning — geen vloeistof vult de ruimte, alleen het gevoel van vacuüm, de cohesie van een waterkolom die ergens anders klimt richting bladerdak maar hier zijn greep heeft verloren. Op het einde van het vat, precies waar een perforatieplaatje had moeten zitten, hangt een lenticulaire luchtembolie als een kwikzilveren spiegel over de volledige doorsnede, zijn convexe meniscus weerkaatst de gehele putgalerij in een bolvormig miniatuur — een stilstaande breuk in het watertransportsysteem, mooi en dodelijk tegelijk.

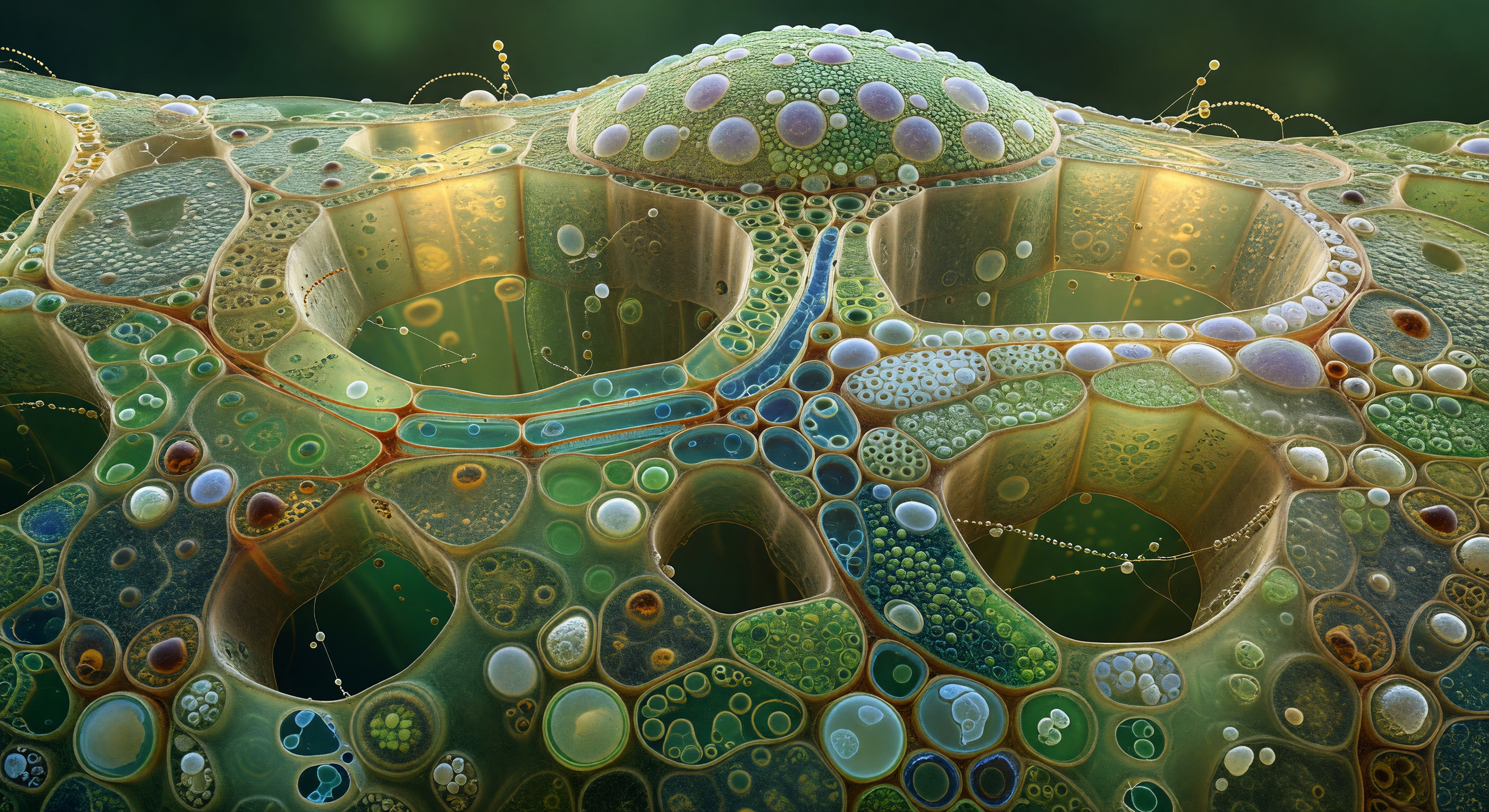
Je staat aan de voet van een woud van paleïde mesofylcellen, omhoogkijkend langs cilindrische torens die zeventig micrometer boven je uitrijzen — kolossale structuren, elk opgebouwd uit halfopake celwanden van cellulose en pectine die amber-groen gloeien in het licht dat van boven neerdaalt. Tegen de binnenwanden zijn honderden lensvormige chloroplasten gedrukt als nauw gevoegde smaragden tegels, elk drie tot acht micrometer breed, hun gestapelde thylakoïdgrana zichtbaar als donkere banden wanneer het licht ze schuin raakt — de dichtheid is zo groot dat de wanden leven als gekleurde glasramen, met overlappende jadegrüne schaduwen die naar de basis van elke kolom toe verdiepen. Boven je hoofd gloeit de bovenste epidermis als een beslagen glazen plafond, de waslaag van de cuticula gevend een licht iriserend glans waar schuine lichtstralen erop vallen, terwijl het licht zelf tussen de kolommen neerdaalt als zachte gordijnen, overal gedempt door de chloroplastwanden zodat geen harde schaduw bestaat — alleen de geleidelijke verduistering van de diepte. Tussen de kolommen door gapen de intercellulaire luchtruimten als smalle, bijna zwarte crevassen, passages die zich zijwaarts uitstrekken naar het sponsachtige mesofyl alsof het steegjes zijn tussen glazen wolkenkrabbers waarvan alle ramen zijn gemaakt van levende, groene lenzen die onophoudelijk licht omzetten in chemische energie.
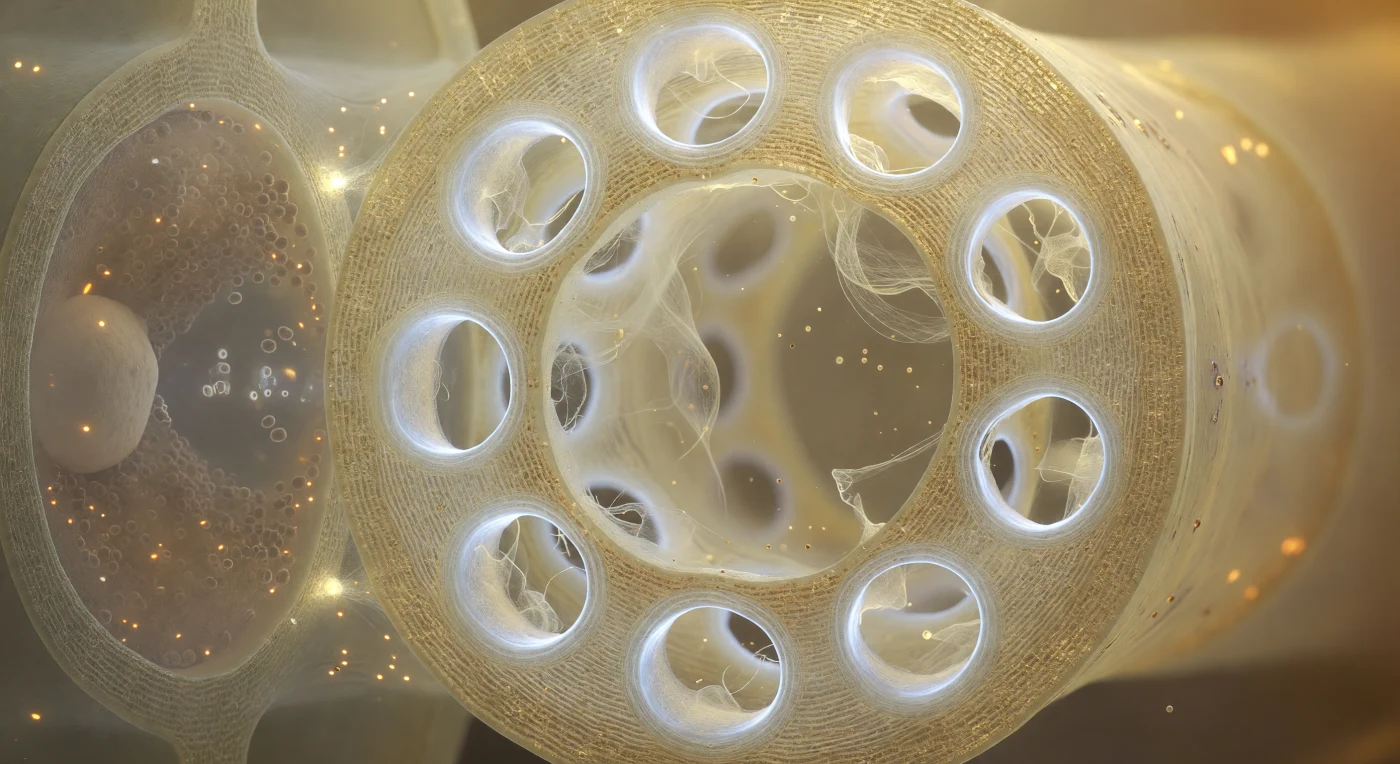
Je staat binnenin een levend zeefvatelement, omgeven door de zachte gouden gloed van het foemlumen, en de gehele eindwand vult je blikveld als een doorschijnend amber scherm: twaalf cirkelronde poriën, elk omzoomd door een dik ringvormig kraagje van callose dat helder wit en licht blauwachtig oplicht, als gepolijst beenderporselein dat het verspreide binnenlicht vangt en verstrooide. Tussen en door de poriën drijven P-proteïnefilamenten als een doorzichtig, licht iriserend web — dunner dan een fractie van een micrometer — zachtjes golvend in de nauwelijks voelbare stroming van foemsap dat zich van de ene kamer naar de volgende perst, in sommige poriën hangend als een haast onzichtbaar gordijn, in andere als slechts een paar gebroken draden die trillen in een onzichtbaar getij. Het floëem is het levende transportstelsel van de plant: zeefvatelementen begeleid door compagnoncellen drijven suikers en signaalmoleculen over afstanden van meters via een osmotisch gedreven drukstroom, terwijl callose de poriën kan verwijden of vernauwen als respons op verwonding of seizoensverandering. Links, door de dunne gedeelde wand, drukt de compagnoncel zich als een donkere, organelrijke massa tegen de heldere holte aan — nucleus, mitochondriën en ribosomen vormen een bijna ondoordringbare dichte nevel — verbonden via plasmodesmata die als haarfijne lichtpuntjes in de wand oplichten, de architectuur van een biologische samenwerking die op deze schaal de intimiteit aanneemt van twee nabuurstaten gescheiden door slechts een membraan.
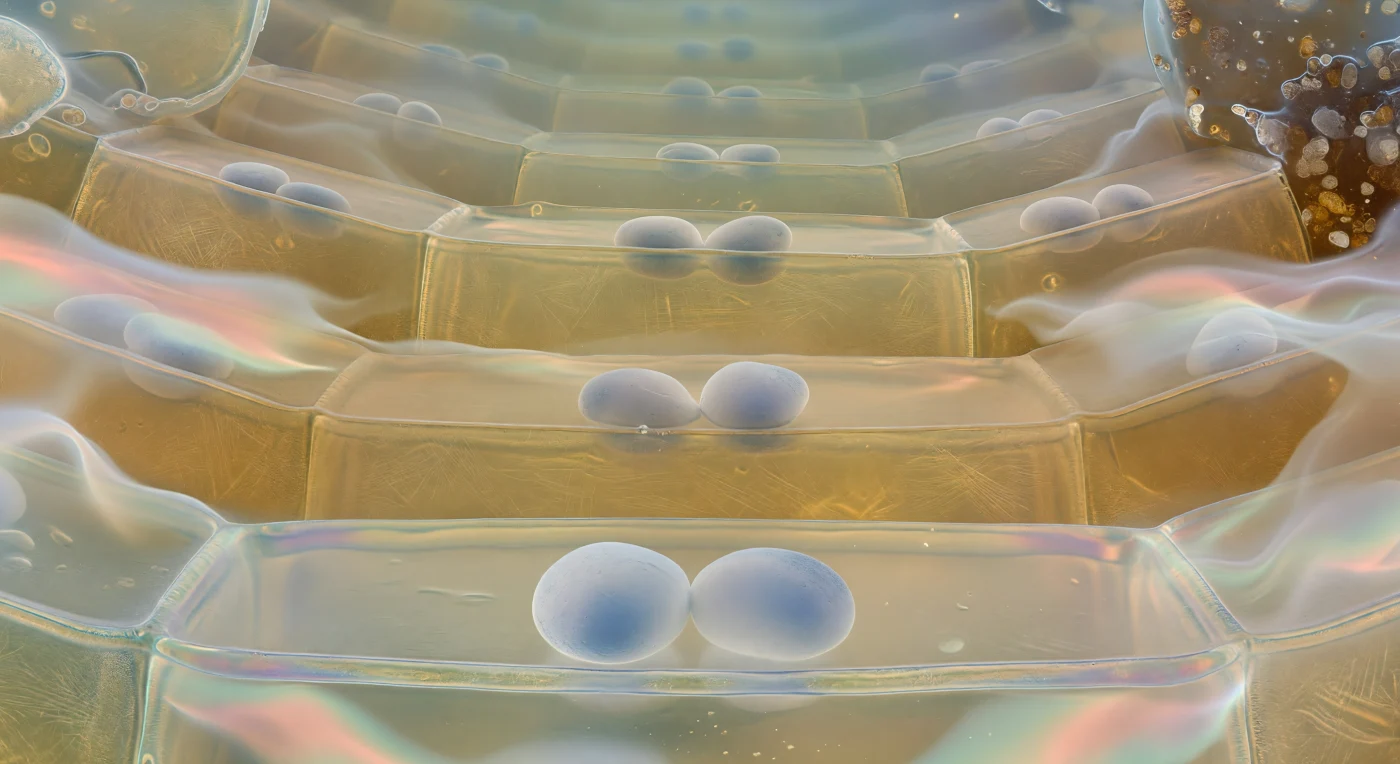
Je bevindt je op de vloer van één enkele columellacel, diep binnenin de wortelhoed van een groeiende plantenwortel, en kijkt omhoog door een kathedraal van levend barnsteenglas. Direct voor je rusten twee of drie amyloplaststatolithen op de gespannen onderste plasmamembraan — dichte, licht doorschijnende ovalen zo groot als keien op deze schaal, hun oppervlak mat krijtwit met een zweempje blauwgrijs waar het licht erdoorheen filtert, volmaakt stil liggend als geslepen stenen op een trommelvel, terwijl het omringende cytoplasma zo helder en kleurloos is als zuiver water. Deze statolithen zijn de zwaartekrachtssensoren van de plant: door te bezinken naar de laagste celwand registreren zij de richting van de zwaartekracht en sturen zo de asymmetrische auxineverdeling aan die de wortel naar beneden stuurt. Boven je rijzen de celwanden op als brede, zacht gebogen vlakken van doorschijnend barnsteen, met het kruiselingse weefsel van cellulosemicrofibrillen als een spookachtig raster gevangen in dikke honinggekleurde hars; door de halftransparante plafonds heen zijn de statolithen van de volgende cellaag al zichtbaar als wazige witte sikkelvormen, en dit patroon herhaalt zich tier na tier tot een zachte piramidaale top waar het warme licht vervaagt in een koelblauwgroene schemerzone. Aan de periferie lossen de buitenste randcellen op in een trillende hydrogelnevel van slijmstoffen — een biochemisch halo dat licht brak in zwakke spectraalbanden van zalmroze en aquamarijn, terwijl daarbuiten de donkere kwartskorrels en kleipartikels van de bodemmatrix glinsteren, nat van het slijm, en de kolossale hydraulische druk van de buitenwereld voelbaar aanwezig is als een stille, alomtegenwoordige kracht.

In het hart van een levende plantencel zweef je voor een minerale explosie: een calciumoxalaat-druse van 55 micrometer breed vult het volledige gezichtsveld als een gotisch roosvenster van gekristalliseerd leven, veertig afzonderlijke kristalvlakken stralend uitwaaiend vanuit een dicht, bijna ondoorzichtig centraal nucleus. Onder gekruiste polarisatoren branden de birefringente vlakken in verzadigde interferentiekleuren — diep kobaltblauw waar de retardatie maximaal is, verschuivend naar verbrande sienna en amber bij schuine hoeken, exploderende tot zuiver goud op de scherpgesneden randen waar constructieve interferentie golflengten opstapelt tot iets bijna gloeiends. De druse is een mineraal verslag van ionen-verzadigingsgebeurtenissen die de cel heeft bevroren in ruimte — calciumionen en oxalaatmoleculen die zijn neergeslagen tot kristallografische onvermijdelijkheid, vermoedelijk als afweer tegen herbivoren of als regulatie van de calciumhuishouding van de cel. Aan de uiterste rand van het beeld tekent de celwand van de parenchymcel zich af als een bleekgouden kader van cellulose-microfibrils, een stille herinnering dat deze minerale kathedraal hermetisch verzegeld leeft binnen één enkele levende cel, het omringende vacuolaire water volledig verslonden door het absolute zwart van de polarisatieopstelling.

Je zweeft boven een koepel die nauwelijks als bol herkenbaar is — pas wanneer je blik de rand volgt waar twee goudgele celwallen als kleine heuvels oprizen, begrijpt het oog dat het kijkt naar een architectuur die zichzelf naar buiten vouwt. Beneden jou ligt het schietpunt van de hele plant: de scheutapicaalmeristeem, een dome van isodiametrische cellen van elk zo'n 12 micrometer breed, hun cellulosewanden zo dun dat ze licht doorlaten als berijpt glas, elk een donkere ovale kern dragend die door de wand heen zichtbaar is als een steen op de bodem van ondiep water. Dit zijn de ongedifferentieerde stamcellen van de plant, dicht gepakt met ribosomen en organellen, nauwelijks gevacuoliseerd, voortdurend delend om alle toekomstige organen — stengels, bladeren, bloemen — uit zichzelf te genereren volgens signaalgradiënten van hormonen als cytokinine en auxine. De twee bladprimordia aan de flanken tonen hoe snel die identiteit verschuift: hun cellen zijn al warmer van kleur, amberkleurig, licht verlengd langs de primordiumas, en de overgangszone tussen meristematisch honingraatpatroon en het beginnende bladweefsel is geen scherpe grens maar een graduele onderhandeling van enkele cellagen breed. Hier, op de schaal van honderd micrometer boven de top van een levend organisme, is schepping geen metafoor maar een geometrisch feit dat je van boven af kunt lezen als een kaart.

Vanuit de substomataire holte kijk je recht omhoog naar de opening van een huidmondje, als een bezoeker die vanuit een kathedraalkelder de sluitstenen van het gewelf aanschouwt. De twee sluitcellen welven zich boogvormig als niervormige poortwachters, opgezwollen door turgordruk tot strakke, glanzende bogen die een lichtende spleet van slechts zeven micrometer omlijsten — een opening die samen met zijn omgeving het ademhalingsritme van de hele plant bepaalt: koolstofdioxide naar binnen, waterdamp naar buiten, beheerst door de asymmetrie van de celwanden die open blijft staan als de chloroplasten fotosynthese aandrijven. Langs de verdikt binnenste wanden liggen twaalf chloroplasten per sluitcel opgesteld in een dichte rij, elk een saffiergroen lenslichaampje van vier tot zes micrometer, gevuld met gestapelde granastapels die het licht van boven absorberen en de cellen van binnenuit doen oplichten alsof ze hun eigen gloed bezitten. De cuticulaire richels overhangen de porieopening als wazig glas — dun genoeg om het licht te breken in een smalle vouw van goud en viooltjespaars — terwijl de omringende epidermiscellen er bleek en kleurloos naast aftekenen, slechts stille plavuizen rondom dit groene kathedraalraam. Dieper in de holte beneden opent de sponsachtige mesofyllabyrint zich in een duistere groene nevel, elk celoppervlak behangt met condenserende waterfilm die het licht verstrooiit tot een levende, vochtige glinstering.

Je bevindt je in het hart van een levende muur, ingeklemd tussen twee werelden die zich in tegengestelde richtingen voltrekken: links verdikken de tracheïden zich rang na rang tot donker okergeel brons, hun secundaire wanden opgebouwd in concentrische ringen van lignine die het licht terugkaatsen als oud koper, terwijl de levende kern van elke cel al is opgegeven aan de leegte — een necropolis van perfect geëngineered hout die zich in de diepte verliest. Rondom je strekken de fusiforme initialen zich uit als kathedraalneuzen van nauwelijks zichtbare membranen, hun strogele cytoplasma een zacht gloeiende lantaarn waarin de opaalachtige kern zweeft als rook gevangen in barnsteenhars, de celwanden zo teer dat ze lijken mee te ademen met elk osmotisch gefluister. Dit is het cambium: een generatief vlies van amper twintig micrometer breed dat de gehele diktegroei van de boom draagt, zijn cellen samengeperst tussen het verhardende rijk van het xyleem en de zachtere, oplossende wereld van het floëem rechts, waar zeefbuizen hun kernen verliezen en callosekragen zich openen als opaalachtige ringen rond sieveplaatporiën, alles bevend op de grens van worden en vergaan. In de lente trilt dit vlies van delingsdrift — elke fusiforme initiaal zal zich tangentieel splitsen en één dochtercell naar het hout sturen, de andere naar de bast, terwijl hijzelf blijft, het stille middelpunt van een groeiende boom.

Voor je rijst de secreterende kop van een klierhaartje op, een trichoom dat de bladoppervlakte van een tomatenplant bewaakt als een architectonische toren. De vier schijfvormige secretiecellen vormen een lage kroon, elk een licht afgeplatte koepel van doorschijnend groen-goud cytoplasma, terwijl de cuticula daarboven is opgezwollen tot een strakgespannen, amber-getinte blaar van geaccumuleerde etherische oliën — terpenen en fenolverbindingen die door de cellen zijn gesynthetiseerd en onder de cuticula zijn opgehoopt tot het punt van bijna-barsten. Het licht breekt door het olieblad in een vloeiende, verschuivende glinstering, fijne dichtheidsgradiënten onthullen de gelaagde secretie van verschillende terpenoïde verbindingen, allemaal gevangen onder de groeiende spanning van de cuticula als een zeepbel op het randje van loslaten. Beneden daalt de zespuntige celsteel af in steeds smallere ringen naar de epidermis, waar de basiscel is verankerd tussen de kenmerkende puzzelstukken-gevormde cellen van het dicotyle bladoppervlak, omgeven door een spaarzaam woud van wasstaven die harde kleine schaduwen werpen over het matte, blauwachtig berijpte oppervlak — zodat het lichtgevende olieblaar erboven flonkert als het enige heldere object in een verder gedempte, geweven landschap van doorschijnende wanden en wasachtige richels.

Op het moment dat je blik de corridor in trekt, bevindt je je in het binnenste van een wortelzone waar cellen zich uitstrekken tot tweehonderdvijftig micrometer in de lengterichting, elk een reusachtige centrale vacuole torend die negentig procent van het celvolume inneemt — een kamer vol optisch zuiver water, strak gespannen onder turgordruk die de celwanden licht naar buiten welft in afgeronde, hydraulisch geladen geometrie. De wanden zelf bestaan uit gelaagde cellulosefibrilllen die het doorvallende licht doorgeven als gepolijst zeeglas, blauw-wit gloeiend aan de vlakke vlakken, iets warmer en dikker bij de hoeken, de weefseltextuur nauwelijks zichtbaar als een fijn geweven patroon wanneer je er dichtbij kijkt. Het levende cytoplasma heeft zich teruggetrokken tot een nauwelijks zichtbare pariëtale film — een bleekgroene biologische residu gedrukt tegen de binnenzijde van elke wand, alleen verdikt waar het een plasmodesma overbrugt of een kleine kern omhult: een amberkleurige bol van tien micrometer die als een lamp drijft aan de rand van een oceaan en de werkelijke schaal van de ruimte eromheen onthuld als duizelingwekkend en onmetelijk. Diep in de corridor, tientallen cellagen verderop, tekent de centrale stele zich af als een donkerindigoblauw axiale kolom, zijn verhoute protoxylemwanden het licht absorberend in plaats van doorlatend, een stil vasculair ruggengraat dat de lumineuze tunnel middendoor snijdt terwijl de turgorspanning van cel tot cel de groei van de wortel naar voren drijft.

Op het moment dat je door de buitenste huid van een boom naar binnen kijkt, wordt de wereld een architectuur van dood en leven in scherp contrast. Om je heen rijzen rijen kurkige rechthoekige kamers op — kurkcellen waarvan de wanden zijn opgebouwd uit suberine-lamellen die gloeien in een diepe mahonieachtige amber, warm als oud cognac tegen kaarslicht, terwijl de holten zelf absolute leegte zijn: verzegeld op alle zes vlakken, nooit meer doordrongen door cytoplasma of organellen. De wanden zijn gevormd door het phellogen, een enkelvoudig levend cellaagje dat als lateraal meristeem voortdurend naar buiten dode kurkcellen afscheidt en naar binnen levende phellodermparenchymcellen genereert — een asymmetrische productie die de peridermis jaarlijks verdikt. Plotseling doorbreekt een lenticel de strakke geometrie: hier ontbreken de suberine-impregnaties gedeeltelijk, de cellen liggen los opeengepakt als complementaire cellen, en door de open intercellulairen kan gaswisseling plaatsvinden — de enige weg waarlangs de levende cellen dieper in de schors zuurstof kunnen bereiken nu de afsluitende kurk elke directe uitwisseling met de buitenlucht blokkert. Elke amberen wand rijst hier meerdere malen hoger op dan jouw veronderstelde positie, en de radiale bestanden van cellen strekken zich uit tot een horizon die nauwelijks merkbaar kromt, als de binnenwand van een enorme houten cilinder die het gehele levende cambium omhult.

Je zweeft in het midden van een reusachtige luchttunnel, omsloten door een continue mozaïek van zeshoekige chlorenchymcellen waarvan de wanden zo dun zijn als nat perkament en flets goudgeel oplichten langs de cellulose-verdikte randen — elke cel een met chloroplasten gevuld kamertje van zo'n 200 micrometer doorsnede, het geheel gewelfd als de binnenkant van een organische drukvat. Dit is het aerenchym van een waterlelie-bladsteel: een netwerk van grote luchtkanalen dat door de plant wordt aangelegd om zuurstof van de bladeren naar de wortels in het zuurstofarme slibbed te transporteren, een levende longas gedragen door plantweefsel. Direct voor je hangt het diafragma: een kanten schijf van stervormige cellen die als armen uitwaaieren vanuit een centraal middelpunt, hun afgeronde uiteinden niet gesloten maar opengelaten in veelhoekige hiaten groot genoeg om doorheen te drijven, terwijl diffuus licht van achter de structuur doorheen sijpelt en elke arm omzoomt met een goudgroene halo. Dunne waterfilmpjes trillen als zeepbellen in sommige openingen en werpen paarse en koperen interferentiekleuren op de groene celwanden, want de lucht hier is verzadigd met waterdamp — geen lucht in de gewone zin, maar een warme, levende atmosfeer opgesloten in het hart van een plantenweefselsysteem dat ouder is dan de eerste bloem. De tunnel verdwijnt in beide richtingen in een teel-blauw vervagende verte, het cellulaire mozaïek steeds fijner wordend als stadslichten door mist, een oneindig zacht corridor van levend groen.
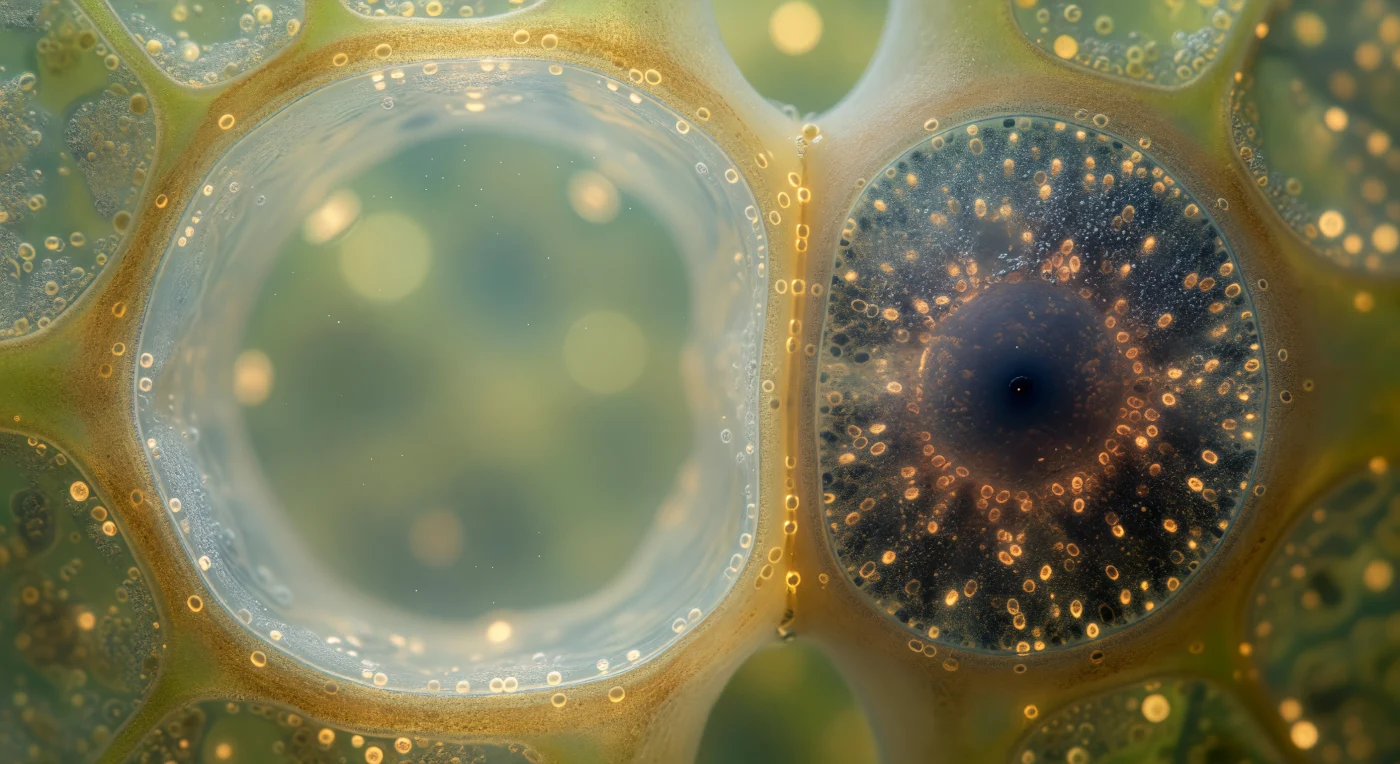
In dit microscopisch universum zweeft u in de vaatbundel van een levende stengel, precies op de grens tussen twee cellulaire werelden die nauwelijks meer van elkaar zouden kunnen verschillen. Links strekt zich het inwendige van het zeefvatelement uit als een kathedraal van bleek barnsteen — een bijna leeg, translucent reservoir waar een trage stroom van opgeloste suikers door beweegt met een snelheid van ongeveer een meter per uur, gedreven door de osmotische drukgradiënten van bron tot put in de plant. Onmiddellijk daartegenaan gedrukt, als een schaduw met lichamelijke massa, ligt de begeleidende cel: een kwart zo breed maar volledig gevuld met biologische urgentie, haar cytoplasma donkerblauw en koffiebruin van de ribosomen en mitochondriën die de ATP-energie leveren om actief suikers in het zeefvatelement te pompen via gespecialiseerde transporteiwitten. De gedeelde wand tussen deze twee cellen — slechts zo'n tweehonderd nanometer dik op de plaatsen waar plasmodesmata doorheen schieten als nanoscopische communicatiekanalen — markeert een van de meest intieme functionele verbindingen in de plantenbiologie, want zonder de metabolische arbeid van de begeleidende cel zou het kernloze zeefvatelement zijn transportfunctie binnen uren verliezen. De omringende parenchymcellen, met hun zachte olijfgroene en chartreusekleurige tinten, omringen dit frapante duo als een publiek dat getuige is van een partnerschap waarop de suikervoorziening van de gehele plant berust.

Op de drempel tussen twee werelden hangt u bewegingloos in de levende wortel van een jonge plant, ingesloten door prismatische endodermiscellen die als kathedraalzuilen om u heen rijzen, elk dertig micrometer breed en gevuld met een trage, barnsteenkleurige stroom van cytoplasma bezaaid met zilverwitte zetmeelkorrels. Precies waar de radiale celwanden van buurcellen elkaar raken, loopt ononderbroken de Casparian strip — een gordel van suberine en lignine die als gegloeid koper gloeit, cel na cel aaneengesloten tot een ongebroken vurige ketting die de gehele stele omsluit en de doorstroom van water en opgeloste stoffen via de celwanden absoluut blokkeert, zodat alle mineraaltransport door het levende cytoplasma van de endodermiscellen zelf moet passeren. Achter u opent het cortexparenchyma zich in een luchtige, blauwzilveren architectuur van dunwandige cellen en wijde intercellulaire ruimten gevuld met vochtige lucht, terwijl voor u de stele zich dramatisch verdicht tot een compact, dieppaars-indigo gewelf van procambiale cellen met prominente kernen — een samengeperst juweeltje van biologische intensiteit. Dit chromatische contrast tussen het warme barnsteenvuur van de strip en het koele maanlichtvioer van de stele maakt de endodermis zichtbaar als wat zij werkelijk is: een architecturaal meesterwerk van gecontroleerde scheiding, de grenspost waar de plant beslist wat de vaatbundel binnenkomt en wat buiten blijft.

Op oog hoogte rijs je als nietige waarnemer temidden van een eindeloos uitgestrekt amberkleurig nettenkasteel, waarvan de aaneengesloten sporopollenine-wallen zich in alle richtingen om je heen uitstrekken als de ruïnes van een versteend kristalrooster. Het sporopollenine zelf — een van de meest chemisch resistente biopolymeren die de natuur voortbrengt, bestand tegen zuren, basen en miljoenen jaren verval — polymeriseerde tijdens de ontwikkeling van het stuifmeelkorrel in een reticulaat patroon dat door inwendige spanning en celdynamiek werd opgelegd, zodat elk onregelmatig zeshoekig lumen een stille getuige is van de biofysische krachten die de exine vormgaven. Schuin licht dat over het oppervlak scheert transformeert elke walrand in een gloeiende kam van warm goud terwijl de diepere lumina wegzinken in ochergele schaduw, een ritme dat zich honderden cellen ver ononderbroken voortzet met absolute, vochtloze helderheid. Aan de horizon opent zich de colpus — een langgerekte bleekgouden vallei waar de exinewanden uitdunnen tot bijna doorschijnend sporopollenine, de apertuurtzone die bij kieming de pollenslang doorlaat en die hier, vanuit dit standpunt, aanvoelt als een tektonische breuk die het bolvormige microwereld doorkruist. Niets beweegt, de lucht draagt geen nevel, en de autofluorescentie van het sporopollenine ademt een ingehouden amber gloed die doet vergeten dat dit hele landschap slechts één enkel stuifmeelkorrel is.
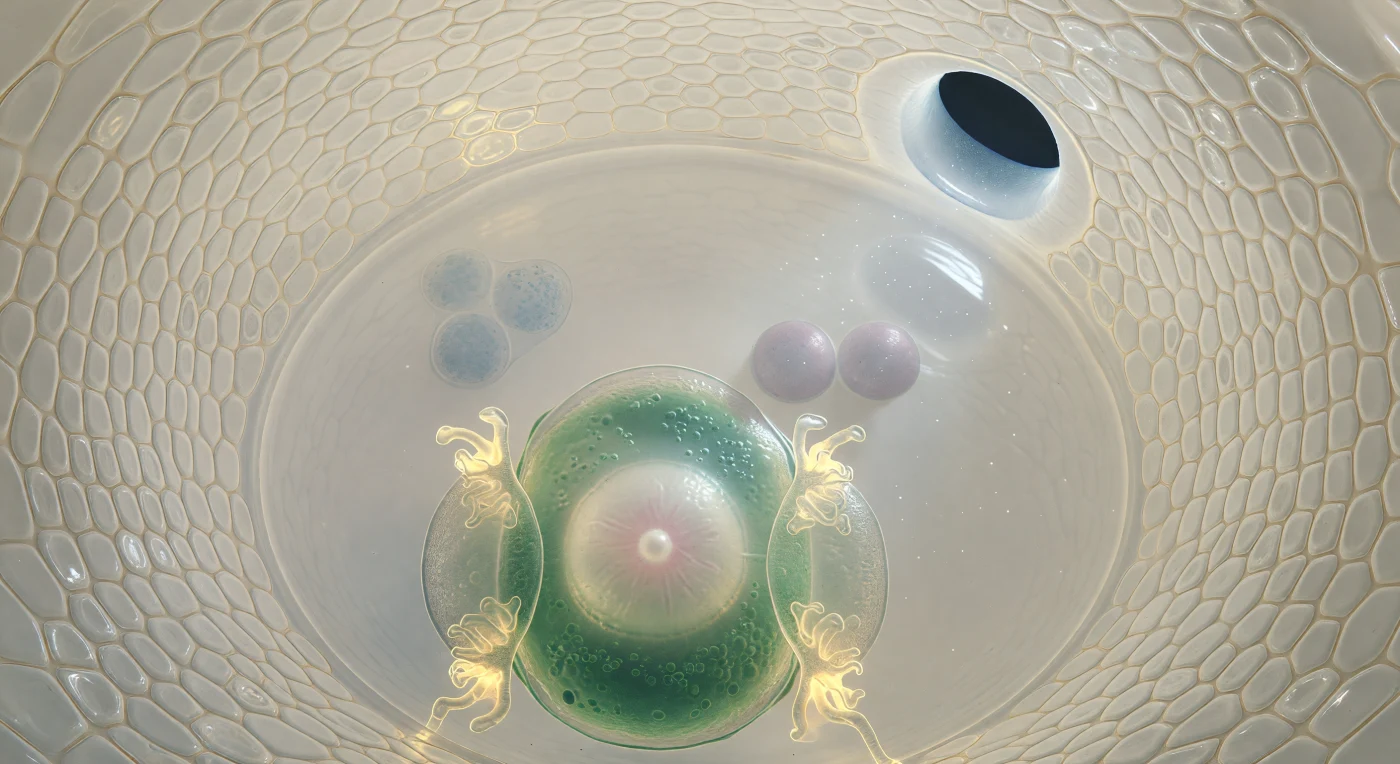
Je bevindt je in de binnenste holte van een eicel-zak, omsloten door twee integumentlagen die zich als de gewelven van een ivoren grot boven je buigen — elke wand een mozaïek van strak aaneengesloten cellen, doorschenen door een diffuus, intern licht dat het weefsel zelf lijkt te emaneren. Op veertig micrometer van de eicel zweef je tegenover haar peer-vormige gestalte: de buitenste cytoplasmazone diep jadegroen van organellen, het inwendige verhelderend rond een groot, alabastwit celkern waarvan de nucleolus glanst als een parel in stil water, terwijl de twee synergidecellen aan weerszijden hun filiform wandinvouwingen tonen als gouden, vertakte filamenten die het licht terugkaatsen als smeulende kool. Achter de eicel strekt de centrale cel zich uit als een kristalheldere zaal gevuld met optisch puur cytoplasma, waarin twee polaire kernen als lavendergrijze manen drijven, en ver weg aan de chalazale pool zijn drie antipodale cellen vaag te onderscheiden — ronde schaduwen vervaagd door de interveniërende diepte, als keien gezien door helder maar diep water. Deze architectuur is het toneel van dubbele bevruchting, het proces waarbij één stuifmeelbuis twee kernen afgeeft: één voor de eicel, één voor de polaire kernen, waarmee zowel het embryo als het voedende endosperm in één enkel biologisch moment worden aangelegd.

Aan de grens tussen vroeghout en laathout zweef je in een tweedeling zo scherp dat ze aanvoelt als een geologische breuk: links gapen de vatengroepen van het eikenhout als donkere zeegrotten, elk lumen een holte van 250 µm doorsnede, omrand door een nauwelijks zichtbare amber halo van behofde poriën die het gepolariseerde licht opvangen als decoratief lijstwerk in versteende architectuur. Dan, abrupt als een klifrand, sluit de wereld zich: rechts vullen de laathoutfibers het blikveld met bijna massieve celwanden, hun kleine gletsjerachtige lumens van slechts 15 µm opengesperd als haarscheuren in obsidiaan, terwijl de secundaire celwanden onder gepolariseerd licht ontploffen in interferentiekleuren van elektrisch kobaltblauw en gebrand goud — een mozaïek van Byzantijns email in organisch steen. Horizontaal door beide werelden snijden de straalparenchymlinten als geologische lagen door zandsteen: honingkleurige baksteenvormige cellen, twee tot vier cellen breed, die de grens tussen vroeghout en laathout volledig negeren en een zachte, matte gloed uitstralen die de iriserende chaos rondom hen verankert. De hele doorsnede staat in vlam onder het gefiltreerde licht, een gebrandschilderd glasraam van levende geologie — kristallijn, monumentaal en volkomen stil.
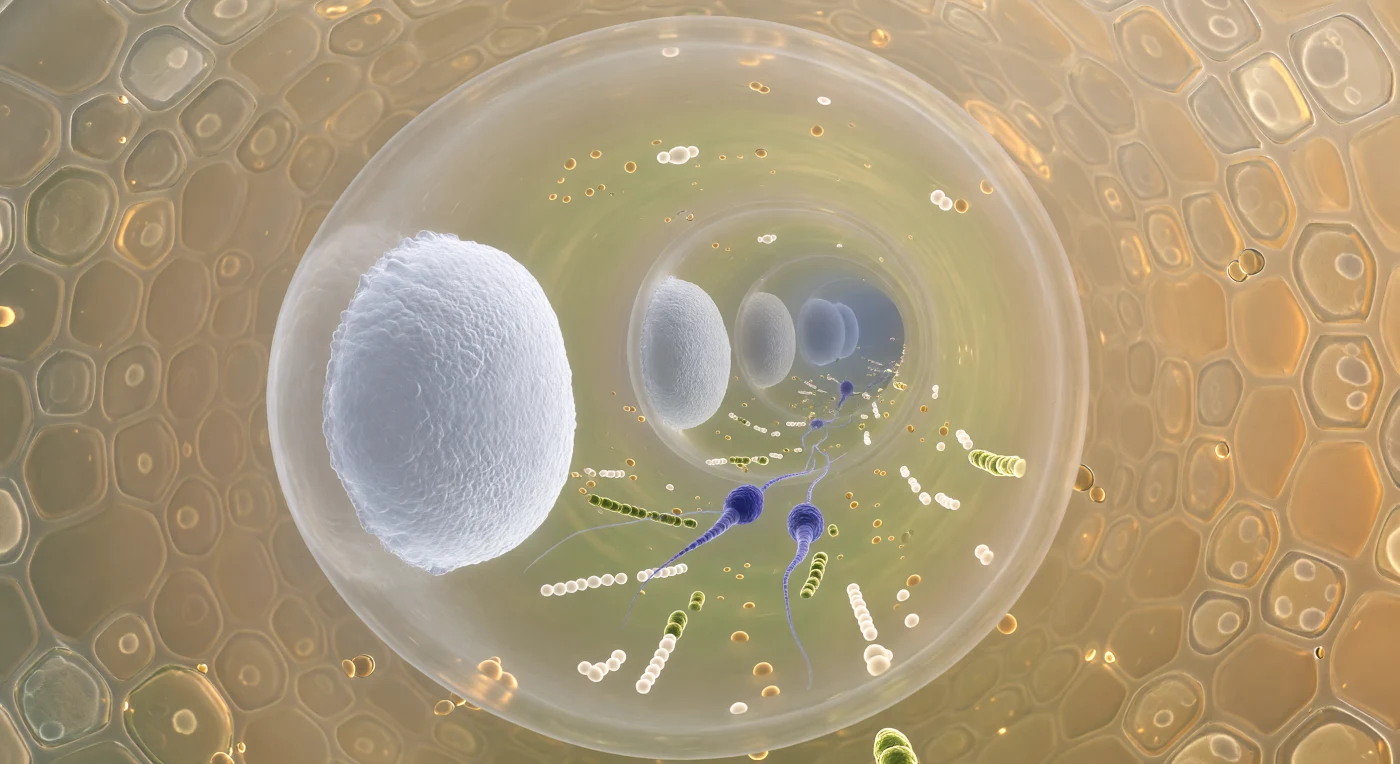
In een fractie van een ademteug bevinden wij ons diep in de tunnel van een groeiende pollenbuis, slechts twaalf micrometer breed, terwijl de licht barnsteenkleurige cellulosewand ons van alle kanten omsluit zoals het interieur van een doorschijnende glazen capillair. Vóór ons rijst een reeks callosepfropfen op als bevroren porseleinwitte bulkhoofden, elk afgedicht met gecomprimeerde (1→3)-β-glucaanvezels die het cytoplasma in afzonderlijke compartimenten verdelen en zo de mechanische integriteit van de buis waarborgen terwijl de punt zich door het stijlweefsel boort. Tussen de dichtstbijzijnde twee schotten draagt een gecomprimeerde stroom organellen — amberkleurige lipidendruppels, gelig oplichtende mitochondriën, clusters van bleekgroene vesikels — twee spoelvormige spermatofyten van vijf micrometer mee naar voren, hun chromatine zo gecondenseerd dat ze als indigo-violette mineralen door de gouden cytoplasmabrij bewegen. Buiten de buiswand drukken de secretoire cellen van het transmissieweefsel van de stijl als een honingraatmozaïek tegen ons aan, hun polysaccharidesecretie diffuus gloeiend als dik barnsteenglas in namiддаgszon, en het geheel herinnert eraan dat dit microscopische corridorstelsel de enige route is waarlangs genetische informatie van stuifmeel de eicel kan bereiken.