Confiance scientifique: Élevé

Au cœur d'un méandre de mésopore argileux, vous avancez à travers un couloir minéral de vingt micromètres de largeur, les parois tapissées de plaquettes de smectite argentée empilées comme des lames de rasoir, leurs bords irisés de reflets nacre et lilas dans une lumière sans source. Devant vous, dominant le passage, l'hyphe mycorrhizienne arbusculaire se déploie comme un cylindre de verre vivant — sa membrane quasi transparente lustrant d'un éclat vert borosilicate, son cytoplasme intérieur animé d'une dérive lente de globules lipidiques crème et de granules ambrés, marée imperceptible mais indéniablement vivante dans ce monde de roche et d'obscurité. Des biofilms bactériens tapissent les anfractuosités entre les plaquettes d'argile, leurs bâtonnets gris-beige ne représentant qu'un cinquième du diamètre de l'hyphe, minuscules villages collés à la pierre à côté de cette colonne biologique qui les écrase de sa présence. Trois tunnels latéraux s'ouvrent dans l'obscurité absolue au fond du pore, leurs entrées cerclées de croûtes organiques ocre, leurs intérieurs engloutis dans un noir sans fond où se prolonge, invisible, le réseau mycorhizien qui tisse silencieusement la forêt au-dessus.
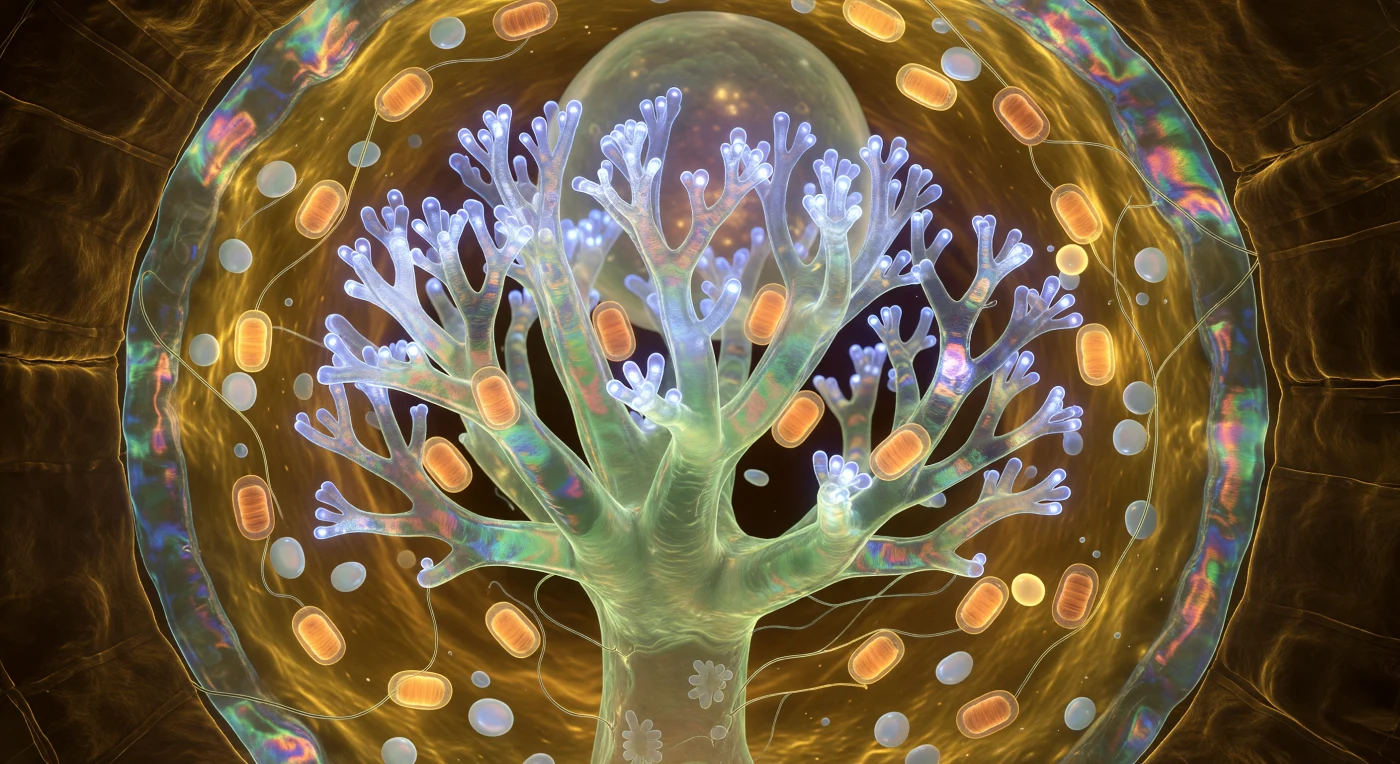
Vous flottez à l'intérieur d'une cellule corticale racinaire comme au cœur d'une cathédrale dont l'architecture aurait été cultivée plutôt que construite : devant vous, l'arbuscule s'élève en forêt de corail inversée, son tronc translucide jade-ivoire se ramifiant en dichotomies récursives jusqu'à ce que les branchettes terminales, larges d'à peine un demi-micron, se dissolvent en un halo tremblant de lumière cobalt électrique — l'éclat froid des protéines transporteuses de phosphate qui pulsent en vagues lentes de la base vers les pointes, seule source lumineuse dans cette obscurité absolue du sol. La membrane périarbusculaire enveloppe chaque branche comme un film de savon tendu à l'extrême, ses interférences optiques faisant glisser la couleur de l'ambre profond vers le rose et le sarcelle aux zones les plus minces, chaque nanomètre de sa surface occupé par des protéines en conformation cyclique dont l'activité collective produit cette bioluminescence métabolique qui éclaire toute la scène par elle-même. Autour du tronc arbusculaire, des mitochondries en forme de cigare se pressent comme des lanternes d'ambre chaud, leurs crêtes internes visibles par transparence, leurs halos respiratoires se superposant dans le cytoplasme — un gel viscoélastique doré, ni liquide ni solide, où dérivent des amyloplastes nacrés qui fractionnent la lumière bleue en traînées de blanc et de violet pâle. La paroi cellulaire referme l'ensemble comme un rempart d'ambre sombre dont les microfibrilles de cellulose en hélices croisées captent la lueur à bas angle, révélant une topographie de nervures et de creux semblable à la voûte intérieure d'une nef en bois sculpté — et dans l'étroit espace apoplastique entre cette paroi et les dernières branches de l'arbuscule, invisible mais réel, s'accomplit en ce moment précis l'échange de phosphate et de sucres qui sustente une forêt entière à des kilomètres de distance.

Dans cette plaine minérale silencieuse, le regard découvre un paysage d'une densité étrange : trois spores de *Glomus* dominent le premier plan comme des galets de résine ambrée, leurs parois concentriques — lamelles translucides de caramel sombre empilées en anneaux — trahissant des structures de protection élaborées capables de survivre des décennies dans le sol avant la germination. À leurs côtés, une spore de *Gigaspora*, crémeuse et rosée, porte en surface des verrues irrégulières caractéristiques de sa famille, tandis qu'une *Scutellospora* lie-de-vin présente son bouclier de germination en croissant — plaque rigide ancrée à la paroi sporale qui orientera la future hyphé vers les racines hôtes. Entre ces masses céramiques, des cristaux de quartz et de feldspath translucides forment une architecture minérale dispersant la lumière en caustiques fantomatiques, et des filaments d'hyphes quasi invisibles — parois de chitine de quelques micromètres d'épaisseur à peine — tendent entre les grains leurs courbes de chaînette fragiles, premier chaînon d'un réseau qui, à d'autres échelles, court sous des forêts entières. Ici s'arrête toute distinction entre le minéral et le vivant : les spores attendent, le sol préserve, et les fils d'argent portent déjà la mémoire chimique de l'arbre auquel ils se raccorderont.
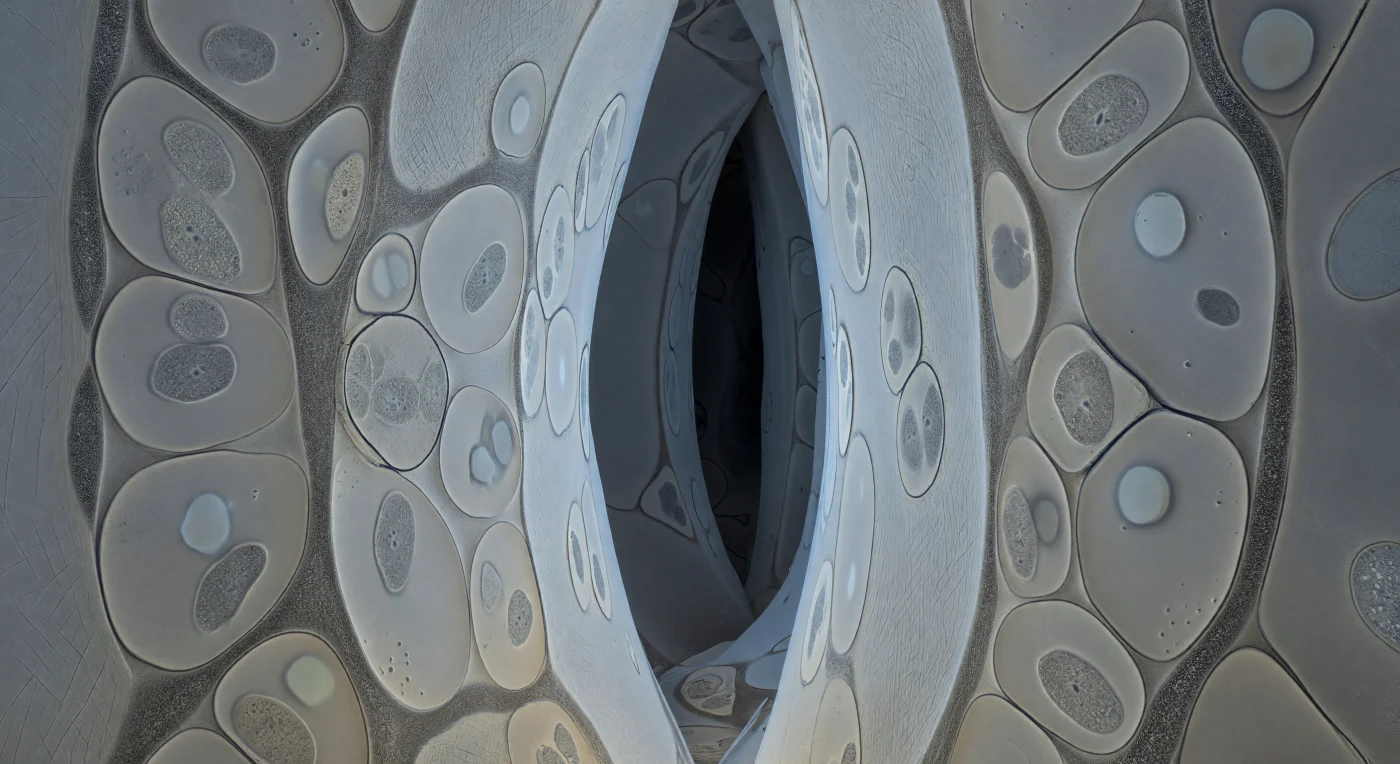
À cette échelle, vous êtes englouti dans un réseau de couloirs intercellulaires si étroits que les parois de cellulose — vastes feuillets crème aux microfibrilles entrelacées comme du papier comprimé depuis des millénaires — vous frôlent de chaque côté, et les sections transversales de hyphes fongiques s'y logent en formes lenticulaires aplaties, leurs membranes plasmiques tracées en graphite dense maintenant une tension presque palpable. Entre la paroi fongique et la paroi racinaire court en continu une bande granulaire de quinze à vingt-cinq nanomètres, maille glycoprotéique sécrétée simultanément des deux côtés : c'est ici, dans cette lisière négociée, que les ions phosphate quittent le cytoplasme fongique et que le saccharose végétal franchit le seuil en sens inverse, échange fondamental de l'ectomycorhize qui conditionne la nutrition de millions d'arbres forestiers. À l'intérieur de chaque profil hyphal, le cytoplasme gris tourterelle est ponctué de mitochondries ovoïdes à peine résolues et de vacuoles lumineuses qui lisent comme des absences optiques, tandis qu'une légère teinte ambrée trahit des gouttelettes lipidiques — réserves carbonées en transit depuis la canopée vers les profondeurs du sol. Les corridors plus lointains se perdent en fentes sombres, les parois y perdant tout détail fibrillaire pour n'être plus que des plans ardoise, rappelant que ce labyrinthe s'étend sans lumière et sans limite perceptible, régi par la seule logique du contraste structural et de la chimie membranaire.
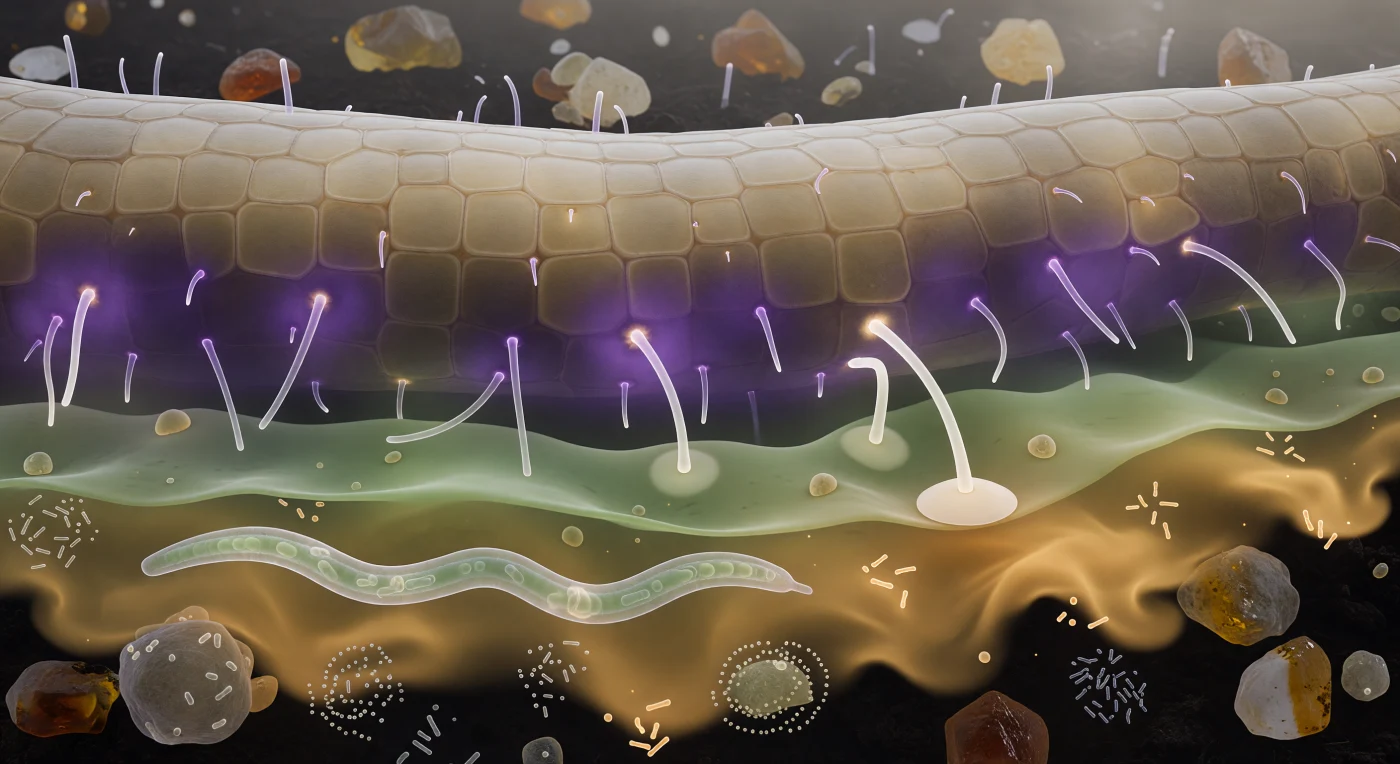
Entre vous et la paroi racinaire, un paysage chimique normalement invisible s'est matérialisé en couches d'aurore superposées : un halo violet dense de strigolactones et de flavonoïdes colle à l'épiderme comme une brume thermale contre une falaise de grès pâle, les poils absorbants — cylindres de verre parfaitement lisses — le traversant en captant sa lumière froide le long de leurs parois comme une couleur d'interférence. Plus loin, à cinquante micromètres de la surface, un gradient vert-gris d'acides aminés se dilue dans les films d'eau inter-agrégats telle une aquarelle saignant dans du papier mouillé, avant de céder sa place à un panache ambre-doré d'exsudats sucrés qui dérive jusqu'à votre hauteur, ses volutes se dissolvant en lambeaux irréguliers dans l'obscurité du sol minéral. Dans la partie inférieure du champ, un nématode translucide de deux cents micromètres progresse en sinusoïde gracieuse au milieu de nuées bactériennes spiralées, tandis que deux pointes hypales — ivoire, lisses — se courbent délibérément à travers la couche verte, suivant le gradient violet comme un navire longe une côte ; l'une d'elles est déjà arrivée, son extrémité aplatie en un large disque d'appressorium parfaitement collé contre une cellule épidermique convexe, déjà en train d'y sonder enzymatiquement la paroi. C'est ici que se négocie, molécule par molécule, le pacte fondamental entre plante et champignon : un échange de carbone contre du phosphore qui structure la fertilité des écosystèmes terrestres depuis quatre cent cinquante millions d'années.

Le sol sous vos pieds est une tapisserie vivante — un plectenchyme serré de cellules fongiques allongées, emboîtées comme des carreaux de céramique posés à la main, chacune de cinq à quinze micromètres de large, leurs parois crémeuses bordées d'un jaune soufre pâle là où les membranes se pressent les unes contre les autres, l'ensemble baigné d'un lustre perlé et froid qui monte de la cellulose elle-même plutôt que d'une source lumineuse extérieure. Derrière vous, la colonne racinaire s'élève comme la coque d'un navire échoué — une muraille lisse et incurvée de tissu brun-fauve aux stries longitudinales, si vaste que sa courbure se perd dans le brouillard avant d'achever son arc, et là où le manteau épouse l'épiderme racinaire, une zone d'ambre plus profond trahit l'entrée invisible du réseau de Hartig, ce labyrinthe intercellulaire qui constitue l'interface métabolique fondamentale de l'ectomycorhize. À la frange du manteau, le tissu tressé se défait en filaments individuels — des hyphes extraradiculaires d'un seul tube transparent, de trois à six micromètres de diamètre, dont les parois réfractent faiblement la lumière nulle comme du verre filé dans le vide, chacun sondant la porosité du sol non colonisé en arcs doucement incurvés, leur cytoplasme en mouvement à peine visible comme un filet de vie interne. C'est depuis ces filaments que la forêt négocie le phosphore contre le carbone, à travers des membranes dont l'échange se joue en secondes, dans une obscurité si totale qu'elle acquiert la densité d'une matière physique.
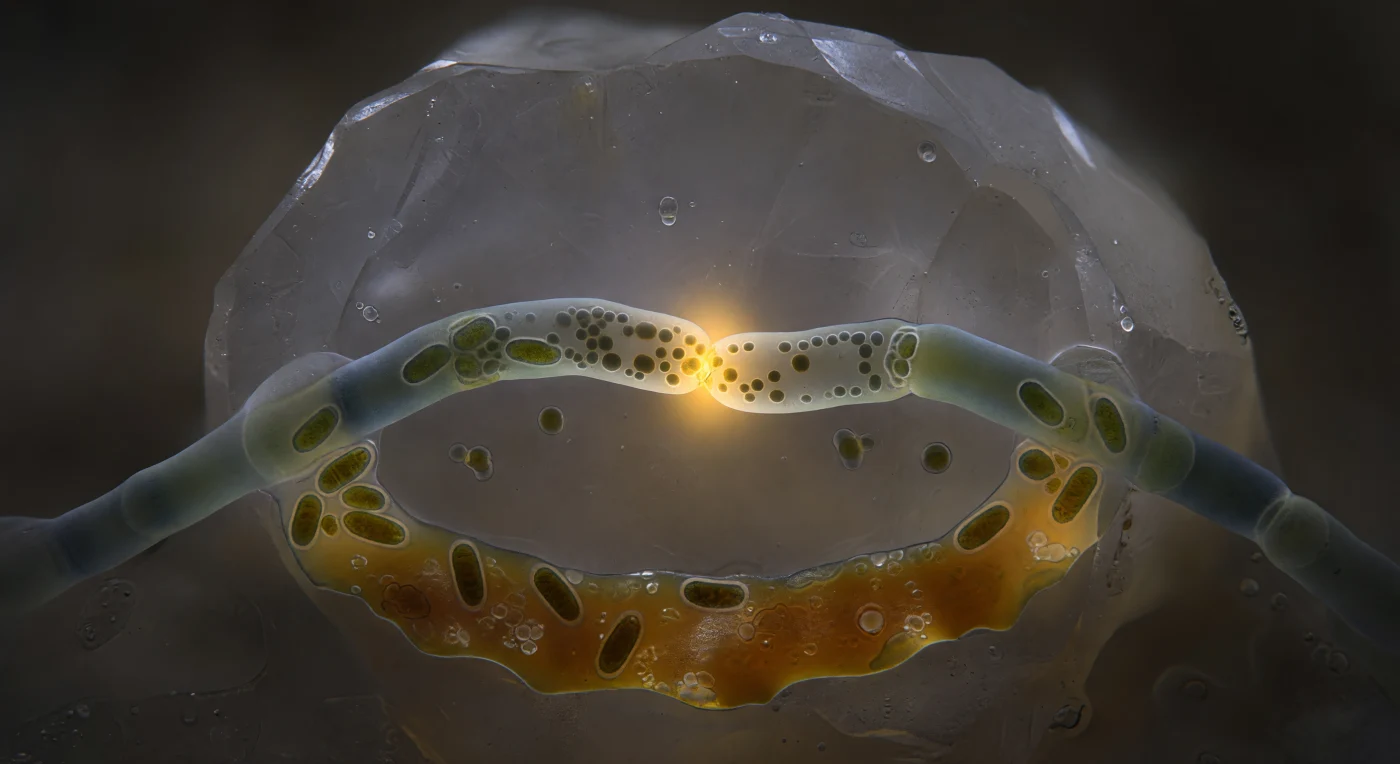
Au cœur d'un pore de sol plongé dans une obscurité absolue, deux hyphes d'un champignon mycorhizien à arbuscules s'incurvent l'une vers l'autre avec une lenteur inexorable, leurs parois de chitine-glucane translucides évoquant du verre de mer dépoli, à peine cinq à six micromètres de diamètre chacune. À l'unique point de contact, les parois cellulaires se dissolvent mutuellement pour ouvrir un pore de fusion d'à peine un micromètre et demi — un seuil biologique où la frontière entre deux organismes s'efface dans un embrasement ambre-or pulsatile : c'est le phénomène d'anastomose hyphal, processus par lequel les réseaux mycéliens se soudent, redistribuent cytoplasme, noyaux et signaux métaboliques pour former des architectures interconnectées capables de coordonner les échanges carbone-phosphore à l'échelle d'une forêt entière. Les mitochondries s'y pressent comme aux portes d'un marché, leurs membranes internes striées de cuivre sombre vibrant d'une activité oxydative intense, tandis qu'un cordon visqueux de granules lipidiques s'écoule du lobe gauche vers le droit dans un flux continu et rythmé. Derrière cette confluence, la falaise de feldspath s'estompe en gris-argent dans le hors-champ, et le film d'eau interstitielle diffuse le halo ambré de la zone de fusion en une auréole douce dans les ténèbres brunes du sol — rendant cette scène à la fois intime comme un geste cellulaire et vertigineuse comme une vue depuis le fond de l'océan.

À quelques centimètres au-dessus de cette surface sombre comme de la lave refroidie, un unique macroagrégat de sol occupe tout le champ de vision, sa courbure s'étirant à l'horizon comme la face d'une lune volcanique : la matrice comprimée de matière organique humifiée, de mélanine fongique ancienne et de silicates minéraux forme une géologie dense, espresso-noir mouchetée d'ocre chaud, que recouvre un film de glomaline — cette glycoprotéine sécrétée par les champignons mycorhiziens arbusculaires — en une fine laque ambre-or rendant la surface légèrement hydrophobe, preuve en est la gouttelette d'eau qui perle parfaitement, lenticulaire et lourde comme du mercure, son angle de contact raide et fier contre la surface scellée. Des fissures équatoriales de l'agrégat émergent des filaments mycéliens blancs en arcs parallèles, pas plus épais que du verre filé, véritables sutures de soie cousues dans le cuir sombre qui maintiennent la structure en cohésion — car c'est précisément ce rôle que joue le réseau fongique : lier les particules minérales et organiques en agrégats stables résistant à l'érosion et à la compaction. Dans le coin inférieur droit, un éclairage ultraviolet révèle ce que la lumière ambiante dissimule : la glomalin explose en fluorescence jaune-vert incandescente, cartographiant en relief lumineux l'architecture invisible du carbone séquestré, blazant le plus intensément sur les arêtes et les bords de fractures où le film protéique est le plus épais, s'estompant en chartreuse dans les bassins creux, tandis que le reste du cadre bascule dans un noir d'espace, comme si deux soleils de longueurs d'onde différentes illuminaient simultanément ce monde miniature que les champignons ont commencé à construire il y a quatre cents millions d'années.
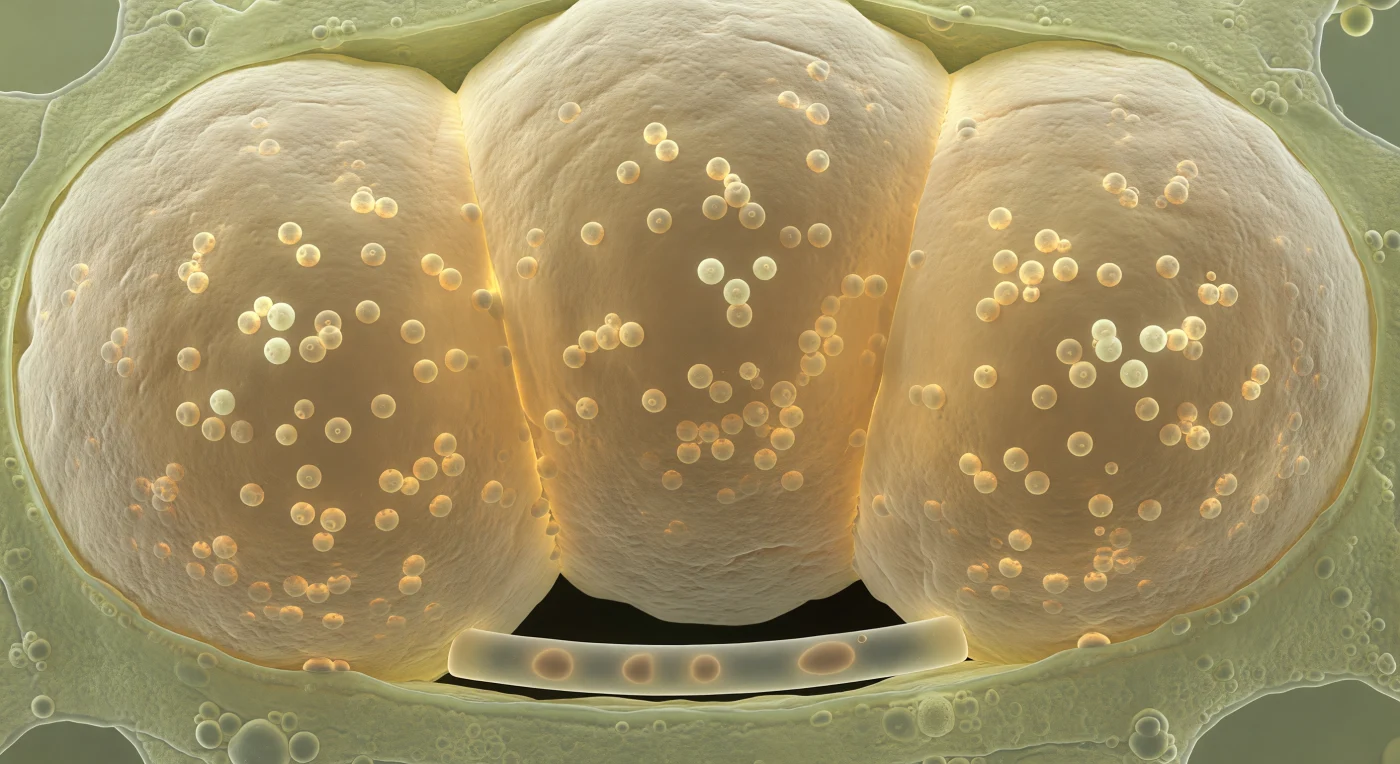
Dans cet espace comprimé, vous vous tenez pris en étau entre trois vésicules arbusculaires mycorrhiziennes qui occupent presque entièrement la cellule corticale de la racine, leurs parois de chitine et de glucanes stratifiées — ivoire rugueux à l'extérieur, ambre vitreux et lumineux de l'intérieur — pressées les unes contre les autres en zones de contact aplaties, comme des ballons trop gonflés pour leur contenant. Chaque vésicule constitue un dépôt lipidique massif formé par le champignon *Rhizophagus irregularis* ou un parent proche, réservoir de triglycérides et d'acides gras qui représentent la monnaie carbonée transférée depuis les photoassimilats de la plante hôte : les dizaines de globules sphériques de cinq à dix micromètres visibles à l'intérieur, flottant en suspension dans un milieu légèrement visqueux, captent la lumière diffuse comme des gouttes d'huile dorée prises dans de la résine. Le cytoplasme de la cellule hôte, réduit à un mince film vert pâle granulaire aux marges, témoigne de la pression exercée par ces structures fongiques intracellulaires qui ne franchissent pourtant jamais vraiment la membrane plasmique — une intimité extraordinaire maintenue par l'enveloppe périvésiculaire. Presque invisible entre deux vésicules, une hype intraradicale solitaire de cinq micromètres de diamètre — cylindre transparent et tendu, à peine plus large qu'un poignet — s'étire vers l'obscurité absolue au-delà de la paroi cellulaire, seul lien physique entre ce coffre-fort lipidique et le réseau externe qui s'étend, invisible, à travers le sol.
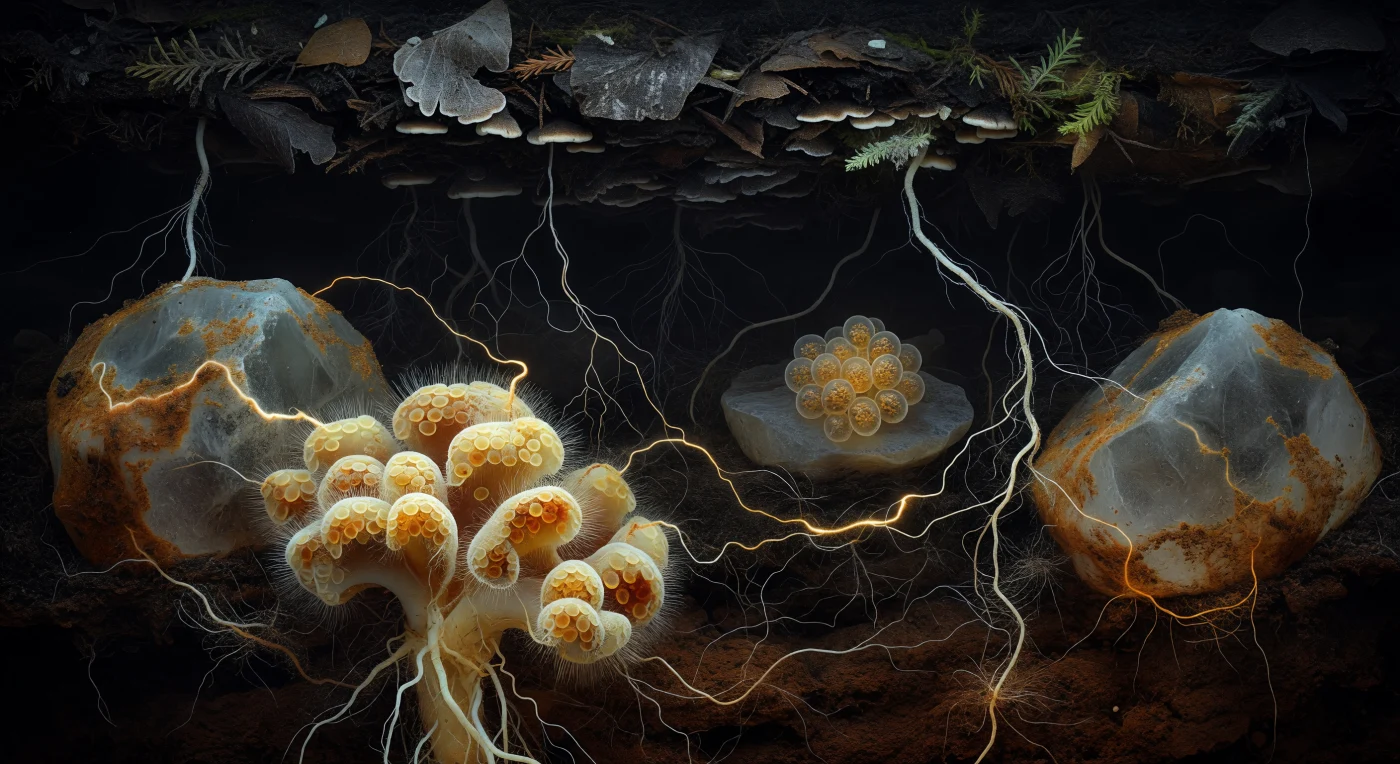
Dans les entrailles obscures d'un sol de vieille forêt, à l'échelle d'un grain de quartz, le regard plonge dans un panorama souterrain que nulle lumière naturelle n'atteint jamais : des agrégats minéraux aux flancs recouverts d'un vernis ambré de matière organique encadrent un réseau de filaments blanc-gossamer si ténus qu'ils semblent tracés à la pointe d'une aiguille à travers la matrice sombre, formant une architecture tridimensionnelle récurrente qui relie racines fines d'épicéa, de hêtre et de chêne sur des dizaines de centimètres de ténèbres. Au premier plan, un manchon ectomycorrhizien domine la scène comme une formation corallienne baroque — ses couches crème, jaune beurre et rouille de pseudoparenchyme enveloppent l'apex racinaire d'une étreinte fongique précise, chaque facette cellulaire humide révélant la frontière entre chaleur biologique et froid minéral. Ce manteau est le siège d'un échange incessant : les sucres photosynthétiques descendent du houppier vers le champignon en quelques heures de flux cytoplasmique, tandis que phosphore et azote remontent vers l'arbre en quelques minutes au niveau des membranes d'interface, une économie chimique invisible qui soutient la forêt entière. À mi-distance, un amas de spores ambres repose contre un plan de feldspath comme des œufs de verre translucide, leurs parois stratifiées et leurs globules lipidiques internes diffusant une lueur mielleuse dans l'obscurité absolue. C'est le réseau mycorhizien commun — la Wood Wide Web — qui donne à ce monde souterrain sa véritable cohérence : des millions de filaments en croissance lente, progressivement plus ténus jusqu'à disparaître dans l'argile plus profonde, tissent entre les arbres de surface un tissu d'interdépendance aussi ancien que la forêt elle-même.

Face à vous, la coupe transversale du rhizomorphe s'ouvre comme la section d'un pilier enfoui, son architecture interne exposée dans l'obscurité absolue du sol : une couronne extérieure de cellules mélanisées densément jointées, noires comme de l'obsidienne carbonisée, forme une armure biologique imperméable à toute lumière, puis cède brusquement la place à une cathédrale intérieure de cellules-tuyaux pâles dont les larges lumens vacuolaires luisent d'une phosphorescence bleu-blanc diffuse, comme des fenêtres de papier de riz mouillé éclairées de l'intérieur. Au centre de cette architecture, un canal axial creux court droit dans l'obscurité, ses parois légèrement humides captant le reflet de cette luminosité intérieure en un mince fil de lumière mouillée. Cette structure n'est pas aléatoire : le rhizomorphe ectomycorhizien est un organe de transport sophistiqué, dont la gaine mélanisée résiste à la compression mécanique et aux attaques microbiennes tandis que les cellules médullaires à paroi mince et à large lumen assurent le flux rapide de l'eau, des sucres et des nutriments sur des distances pouvant atteindre plusieurs mètres à travers le réseau forestier. Autour du bord extérieur, de fines hyphes satellites s'irradient comme des fibres optiques dans le labyrinthe compressé de grains de quartz et d'agrégats argileux — l'interface active où ce câble biologique enfoui négocie silencieusement les échanges chimiques qui soutiennent les forêts entières.

Depuis cette altitude de poussière suspendue, le regard embrasse un paysage entièrement gouverné par la matière vivante en train de conquérir un territoire obscur. Un front mycélien d'arbuscules à mycorhizes arbusculaires (CMA) s'étale devant vous comme un delta fluvial vu du ras du sol : depuis un hyphe conducteur central, des dizaines de ramifications s'élancent en éventail vers une paroi racinaire à peine devinée dans la pénombre brune, chaque pointe arrondie rayonnant d'une lueur cytoplasmique bleu-blanc froide, dense d'organites mobilisés pour la progression. Les segments plus anciens, en retrait du front, révèlent leur architecture interne — de grandes vacuoles claires se succèdent à intervalles réguliers, transformant ces filaments en chapelets de perles de verre dépoli légèrement jaunies aux ventres lipidiques, témoins du stockage et du transit des ressources carbonées captées par la plante hôte en échange de phosphore minéral. Certains hyphes s'insinuent en file indienne dans des chenaux de huit micromètres à peine, déformant légèrement leur section en ovale sous la contrainte minérale, tandis que d'autres franchissent des macropores ouverts comme des câbles tendus entre deux falaises de quartz ferrugineux. Sur les grains de calcite couleur crème, des auréoles jaune-orangé de dissolution oxalique marquent les points d'attaque chimique du champignon, une chaleur soufrée qui contraste avec la froideur spectrale du front vivant, rappelant que cet organisme invisible à l'œil nu démantèle littéralement la roche pour nourrir la forêt entière.
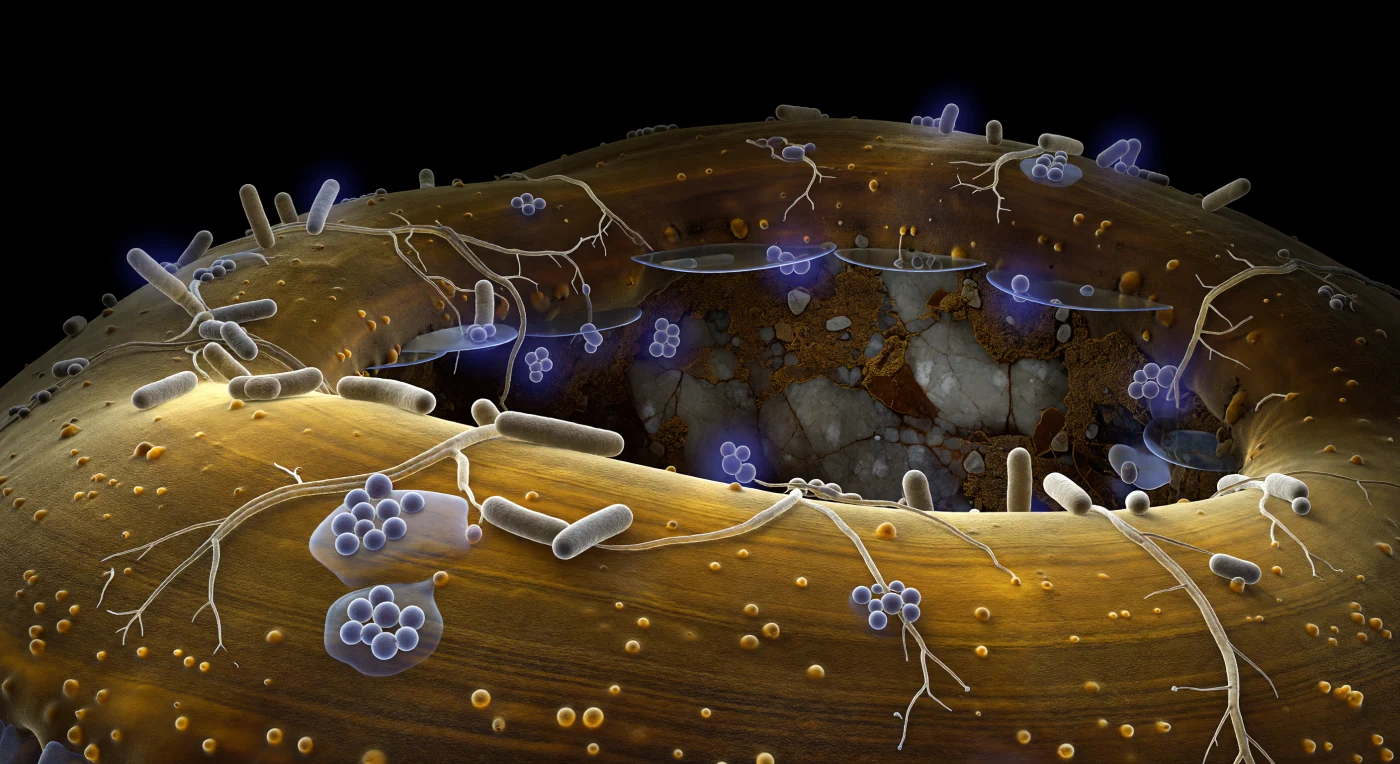
Au ras de la paroi d'une hyphe ectomycorhizienne, la surface ambrée s'incurve comme le flanc d'une falaise de grès poli, backlit de l'intérieur par la lueur froide du cytoplasme à travers la paroi chitineuse semi-translucide — seule source lumineuse dans cette nuit souterraine absolue. Des bactéries en bâtonnets gris-beige, trois fois plus hautes que le point d'observation, s'agrippent à cette topographie en colonie dispersée ou en petits amas où un gel d'exopolysaccharide translucide, semblable à de la glycérine renversée, capte et réfracte la lueur ambiante en halos prismatiques discrets. Des filaments de Streptomyces tracent des réseaux ramifiés à la surface de l'hyphe, ancrés par des attaches adhésives quasi-invisibles qui ne se révèlent qu'à l'instant précis où la lumière chimique effleure leur angle de contact. Certaines bactéries isolées sont enveloppées de halos diffus en violet profond — molécules signal rayonnant en gradients sphériques qui s'éteignent en quelques longueurs de cellule dans le film aqueux environnant, leur conférant l'aspect de lanternes tenues contre la paroi ambrée. Dans le fond, à peine discernable dans l'obscurité du pore, une surface minérale de feldspath fracturé scintille brièvement là où un film d'eau-miroir attrape, une dernière fois, le reflet de cette lumière intérieure.

Vous flottez dans l'obscurité totale d'une fissure de sol humide, et devant vous, la graine de *Dactylorhiza* occupe tout l'espace visible comme une lanterne de papier translucide amarrée contre une falaise de grit basaltique noir — son tégument, membraneux et iridescent, ne fait qu'une cellule d'épaisseur, difractant la chimie des eaux porales en reflets argentés et bleu-vert, laissant entrevoir l'embryon intérieur comme une constellation de cellules pâles suspendues dans une attente métabolique. Depuis le bord inférieur gauche, l'hyphe de *Rhizoctonia* avance tel un cylindre laqué d'ocre et d'ambre, sa paroi de chitine épaisse comme du bambou verni, sa cytoplasme animée de courants lents de granules, et là où elle a percé le tégument, le tissu vivant s'est refermé en un anneau serré autour du point d'entrée. À l'intérieur d'une cellule embryonnaire, le péloton qu'elle a formé remplit l'espace cellulaire entier : un enroulement dense et impossible d'hyphes repliées sur elles-mêmes des dizaines de fois, resserré comme un ressort de montre, déformant la paroi cellulaire en légère ellipse sous sa pression — non pas une invasion, mais une négociation biochimique entre deux organismes qui décident ensemble si cette rencontre sera parasitisme ou alliance, fondement de presque toute la flore terrestre. Les cellules voisines, déjà légèrement élargies et réchauffées d'une lueur crème-dorée, annoncent les premiers pas moléculaires de la différenciation, tandis que le reste de la scène retombe dans un noir minéral absolu, épais comme une présence matérielle, troué seulement, à la limite de la netteté, de minuscules bâtonnets bactériens accrochés à des surfaces de quartz comme des bernacles sur une falaise sous-marine.

Suspendu à l'échelle d'un apex hyphal au cœur d'un pore du sol, le regard plonge dans une cathédrale de ténèbres ambrées : des tours de quartz hautes comme des immeubles de bureaux luisent de leur pellicule de glomaline couleur de miel cru, tandis qu'au-dessus, le cortex racinaire mort s'effondre en rubans de cellules brun chocolat et noir profond, laissant échapper en slow motion des spores de champignons mycorhiziens arbusculaires — sphères et ellipsoïdes de la taille de rochers ou de maisons à cette échelle, leurs parois épaisses irisant l'ambre, le rouge-vin et l'ocre, certaines déjà fissurées et suintant un fluide lipidique pâle dans le film d'eau. À mi-distance, les filaments résiduels du réseau mycorhizien commun courent horizontalement comme des câbles argentés et des fils de soie, portant encore la lueur translucide d'un flux cytoplasmique qui se dépressurise lentement depuis la mort de l'arbre, reliant les survivants bien au-delà de l'horizon minéral. Contre cette élégance, le mycélium saprotrophique colonise les ruines du cortex avec une architecture franchement agressive : hyphes crème et opaques, presque deux fois plus larges que les filaments mycorhiziens, grippant la matière en décomposition de leurs anastomoses denses et mats. Depuis la gauche avance un voile de filaments jaune pâle captant la lumière ambrée des minéraux, depuis la droite une nappe blanc-crème aux branchements aigus qui commence déjà à contacter les premières spores libérées — deux réseaux convergents se disputant en silence l'héritage chimique colossal d'un géant tombé.
