Affidabilità scientifica: Alto

Ci troviamo all'interno di un mesoporoso del suolo, stretti tra pareti di lamelle smectitiche argentee e massi di quarzo traslucido, avanzando come viaggiatori di cinque micrometri in un labirinto minerale di assoluta oscurità. Davanti a noi si incurva l'ifa di un fungo micorrizico arbuscolare: un cilindro vitreoso con un tenue riflesso verde, la cui parete quasi trasparente rivela il lento fluire del citoplasma — granuli ambrati e globuli lipidici crema che si spostano con la deliberatezza silenziosa di una corrente sottomarina. Le pellicole organiche di colore ambra-ocra rivestono ogni superficie minerale come vernice antica indurita, mentre colonie di bastoncelli batterici — appena un quinto del diametro dell'ifa — si addensano in biofilm lungo gli angoli riparati tra le platelet argillose, rimpiccioliti dalla monumentalità del filamento vivente che domina il corridoio. Tre gallerie laterali si aprono nella parete opposta e nel pavimento del poro, inghiottite da un'oscurità impenetrabile, incorniciate da platelet sporgenti e croste organiche, evocando l'intimità schiacciante di una grotta sottomarina non cartografata, ridotta alla larghezza di un'unica cellula umana. In questo mondo privo di luce, ogni scambio — carbonio verso la radice, fosforo verso l'albero — avviene per chimica, per tensione superficiale, per il tocco impercettibile di membrane che si riconoscono nell'oscurità.
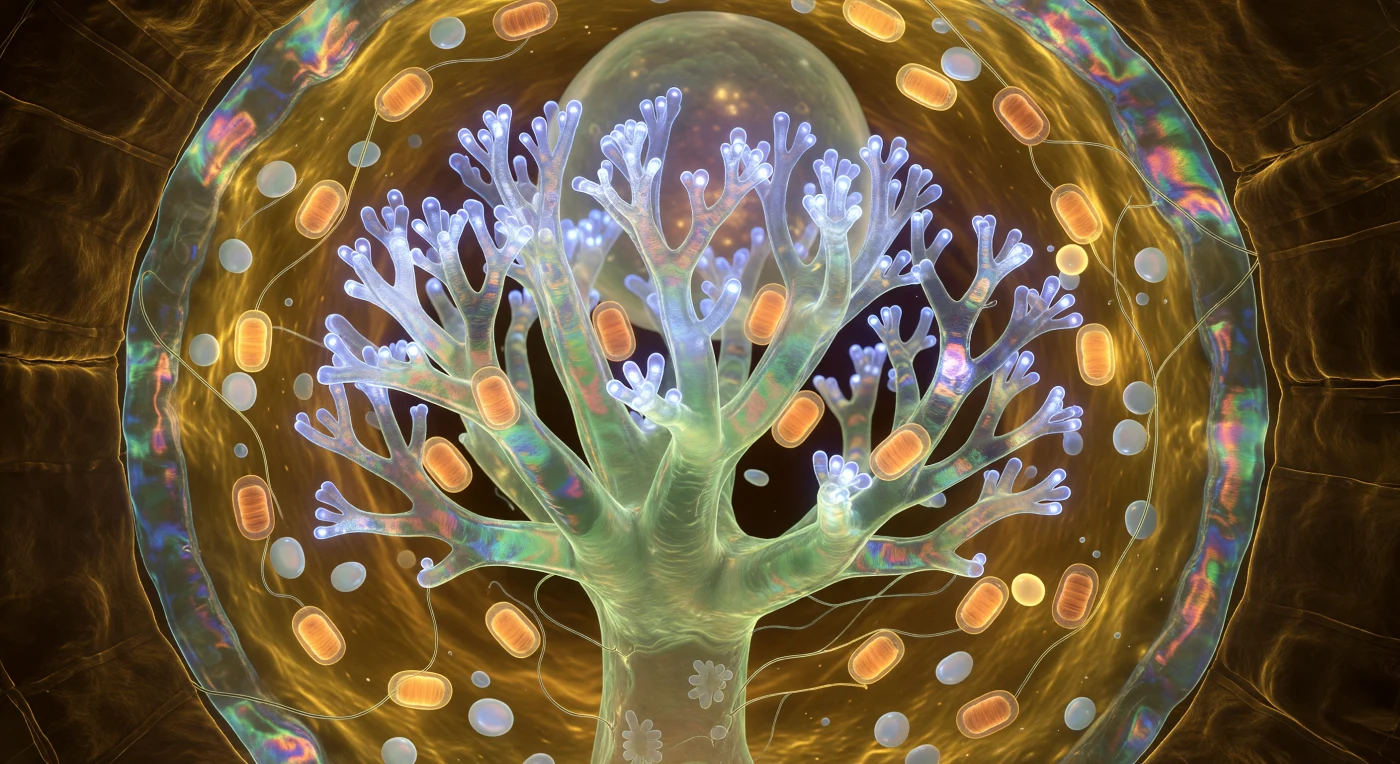
Sospesi al centro di questa cattedrale vivente, i vostri occhi incontrano per prima cosa l'arbuscolo: un albero di corallo invertito che si innalza davanti a voi in dicotomie sempre più fini, il suo tronco principale una colonna di giada traslucida da cui si diramano rami che si assottigliano fino a dissolversi in un tremore di luce cobalto-bianca, generata non da alcuna fonte esterna — qui la luce solare non penetra mai — ma dall'attività ciclica dei trasportatori di fosfato incastonati spalla a spalla nella membrana periarbuscolare, la cui iridescenza ambra-verde-rosa è il segno visibile di uno scambio biochimico che avviene in questo preciso istante. Questa struttura, che misura appena una ventina di micrometri nella sua interezza, rappresenta il cuore anatomico della simbiosi micorrizale arbuscolare: il fungo — appartenente al phylum Glomeromycota — ha spinto le sue ife all'interno della cellula corticale della radice ospite senza mai rompere la membrana plasmatica di quest'ultima, costringendola invece a piegarsi e avvolgersi attorno a ogni ramo come un guanto elastico, creando un'interfaccia di scambio la cui superficie totale supera di decine di volte quella della cellula originaria. Attorno al tronco arbuscolare, mitocondri a forma di sigaro ardono come lanterne ambrate, la loro respirazione chemiosmotica alimentando la domanda energetica enorme di una cellula impegnata a esportare zuccheri verso il fungo e a importare fosforo dal suolo, in un commercio che sostiene foreste intere a scale milioni di volte superiori a questo singolo vano luminoso. La parete cellulare si arcua tutto intorno come un bastione di ambra scura, le sue microfibrille di cellulosa intrecciate in eliche che catturano la luce metabolica dell'arbuscolo a basso angolo, ricordando la volta intagliata di una cappella costruita non da mani umane ma da quattrocento milioni di anni di coevoluzione silenziosa.

Nel suolo sabbioso si apre un paesaggio minerale di silenziosa vastità: le spore di funghi micorrizici arbuscolari dominano la scena come monumenti sferici, le loro pareti laminate di ambra e miele che catturano la luce diffusa in ricchi giochi di chiaroscuro, mentre i granuli di quarzo e feldspato rosato si innalzano tutt'intorno come boulder traslucidi che rifrangono la tenue luminescenza in caustica bianche e blu fantasma. Le spore di *Glomus*, *Gigaspora* e *Scutellospora* — ognuna distinta per tessitura, colore e struttura di germinazione — rappresentano riserve di lipidi e citoplasma racchiuse in pareti pluristratificate, capaci di sopravvivere decenni nel suolo in attesa della prossima radice ospite. Fra di esse, filamenti ifali argentati quasi invisibili si tendono in morbide curve catenarie attraverso i vuoti inter-granulari, strutture dal diametro di pochi micrometri che costituiranno, una volta germinate, la rete di scambio attraverso cui carbonio vegetale e fosforo del suolo transiteranno in minuti o ore. Scaglie scure di materia organica decomposta punteggiano il pavimento sabbioso come lastre di resina petrolizzata, mentre una foschia minerale vela la distanza media, conferendo profondità a un paesaggio dove immobilità geologica e architettura biologica vivente coesistono nel buio permanente del sottosuolo.
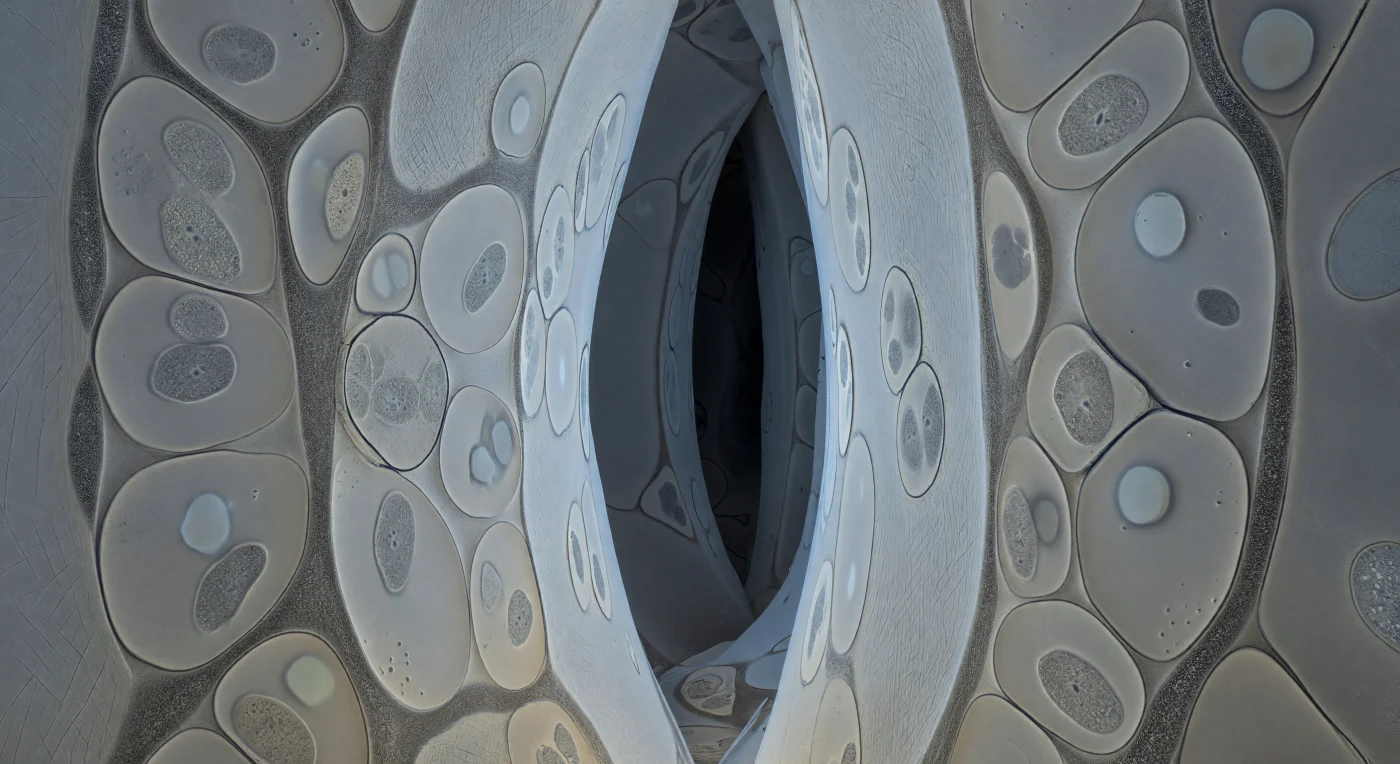
Ci si trova sospesi all'interno di un corridoio intercellulare strettissimo, stretto tra vaste pareti di cellulosa color avorio pallido che si innalzano come facciate di edifici antichi, le loro superfici percorse da fasci di microfibrille disposti in diagonali incrociate come la grana di una carta compressa nei secoli. Ogni interstizio è occupato da sezioni trasversali di ife fungine schiacciate in profili lenticolari, i loro contorni di membrana plasmatica tracciati in una linea grafite tagliente che conserva la tensione di curvatura come un bordo teso, mentre all'interno il citoplasma grigio tortora custodisce mitocondri puntiformi e vacuoli lucenti che appaiono come assenze ottiche nel buio. Tra la parete fungina e quella vegetale corre senza interruzione una striscia granulare di 15–25 nm di matrice interfacciale — né membrana né vuoto, una soglia negoziata di glicoproteine secretate da entrambi i lati — che in questo istante congelato è il luogo dove ioni fosfato abbandonano il citoplasma fungino e saccarosio arriva dalla pianta, l'intera economia biochimica della simbiosi ridotta a una texture appena più calda di ciò che la circonda. La scena retrocede verso corridoi più scuri, piani di ardesia profonda privi di dettaglio fibrilare, e l'unica nota cromatica è un impercettibile ambra tiepida dove goccioline lipidiche si addensano in un angolo basso di un profilo ifale — tutto il resto è contrasto strutturale puro, un mondo interiore dove ogni interfaccia proietta la propria ombra sottile e la geometria stessa diventa illuminazione.
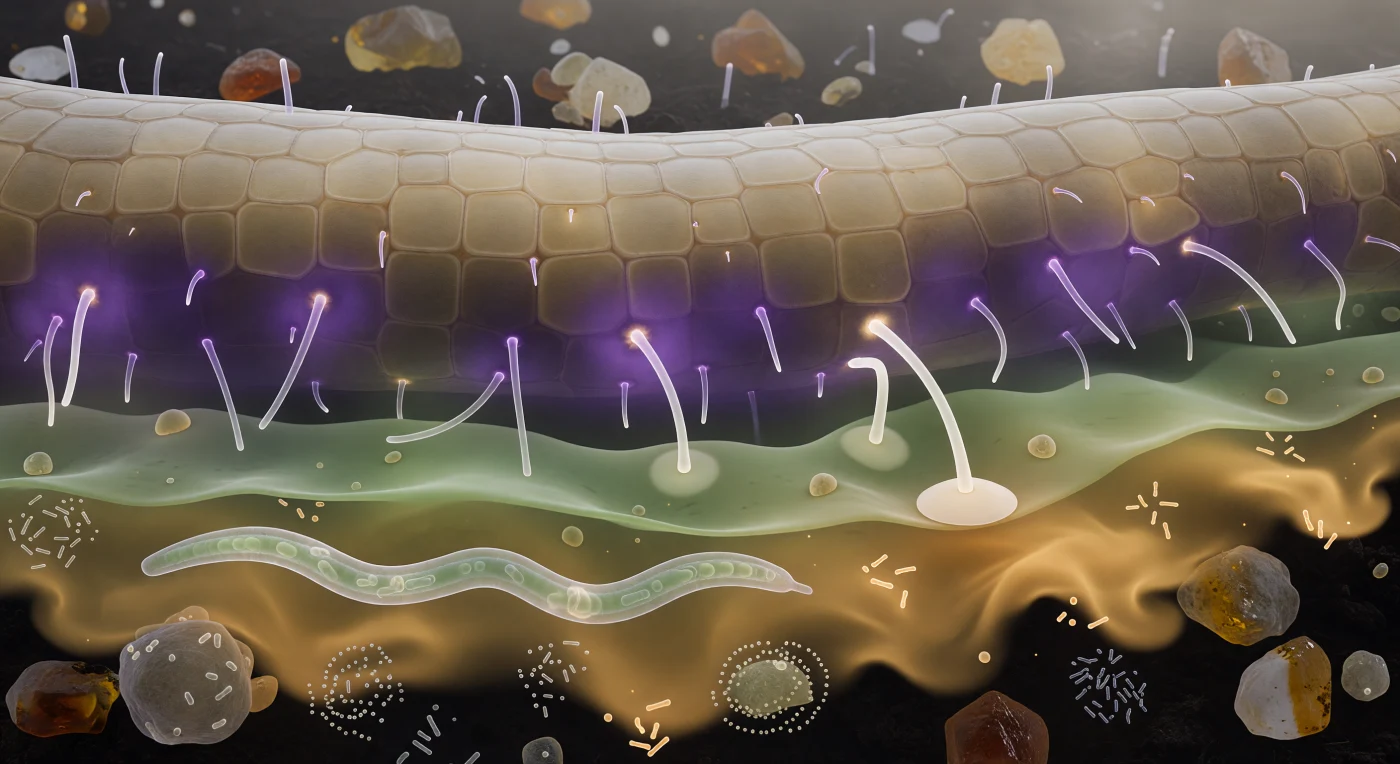
Sospesi a mezza strada tra la parete radicale e l'oscurità del suolo, si percepisce la radice come una falesia di arenaria pallida che si incurva verso l'alto fino a occupare metà del campo visivo, i suoi peli radicali proiettati come cilindri di vetro che captano la luminescenza chimica della rizosfera e la restituiscono come filamenti bianchi e freddi. Lo spazio tra noi e quella superficie non è vuoto: è una stratificazione di atmosfere invisibili rese finalmente percepibili, una corona viola densa di strigolattoni e flavonoidi che abbraccia le cellule epidermiche, poi un gradiente verde-grigio di aminoacidi che si diffonde come acquerello in carta bagnata, e oltre ancora un pennacchio ambrato di essudati zuccherini che deriva verso di noi con la fluidità di una corrente abissale bioluminescente. In basso, un nematode translucido attraversa la scena con un moto sinusoidale misurato, circondato da sciami batterici che pulsano in spirali sulle superfici degli aggregati minerali, mentre due punte ifali si piegano visibilmente verso la radice seguendo il gradiente violetto come una nave segue la costa. Una di esse è già arrivata: il suo apice si è appiattito contro una piastrella epidermica in un disco appressorio perfettamente aderente, mentre comincia in silenzio a esplorare la parete cellulare sottostante con i propri enzimi, primo contatto concreto tra il fungo e la pianta ospite.

La superficie su cui ti trovi è un tessuto vivente — una plectenchima fitta di cellule fungine allungate, disposte come mattonelle smaltate a mano, ciascuna translucida al centro e orlata di giallo zolfo pallido dove le membrane si toccano, l'insieme che emette una luminosità perlacea fredda sufficiente a rivelare ogni giuntura e rilievo della trama come sampietrini bagnati di rugiada al chiaro di luna. Dietro di te si erge la colonna radicale come lo scafo di una nave incagliata, una parete immensa di tessuto bruno-tabacco dalla curvatura così ampia da dissolversi nella foschia prima di piegarsi fuori dalla vista, mentre alla giunzione tra manto e epidermide radicale un'ambra più profonda tradisce l'inizio invisibile della rete di Hartig che si insinua tra le cellule come luce di lampada attraverso carta di riso. Ai tuoi piedi, granuli di quarzo e scaglie di argilla — massi dalla tua prospettiva, rivestiti di una pellicola organica ambrata — si sono incuneati nelle depressioni della superficie tessuta, premendo contro il tessuto vivo con l'indifferenza della sedimentazione. All'orlo sfrangiato del manto, qualche decina di passi davanti a te, le ife extraradicali si lanciano nell'oscurità assoluta del poro del suolo come filamenti di fibra ottica: tubi trasparenti di tre-sei micrometri, la parete rifrangente come vetro soffiato, il contenuto citoplasmatico appena visibile come un filo interno in lento movimento, ciascuno che sonda l'ignoto mentre il contrasto tra quella luce vitrea e il nero fisico del suolo non colonizzato conferisce alla scena la qualità di una grotta percorsa solo dalla luce degli esseri che la abitano.
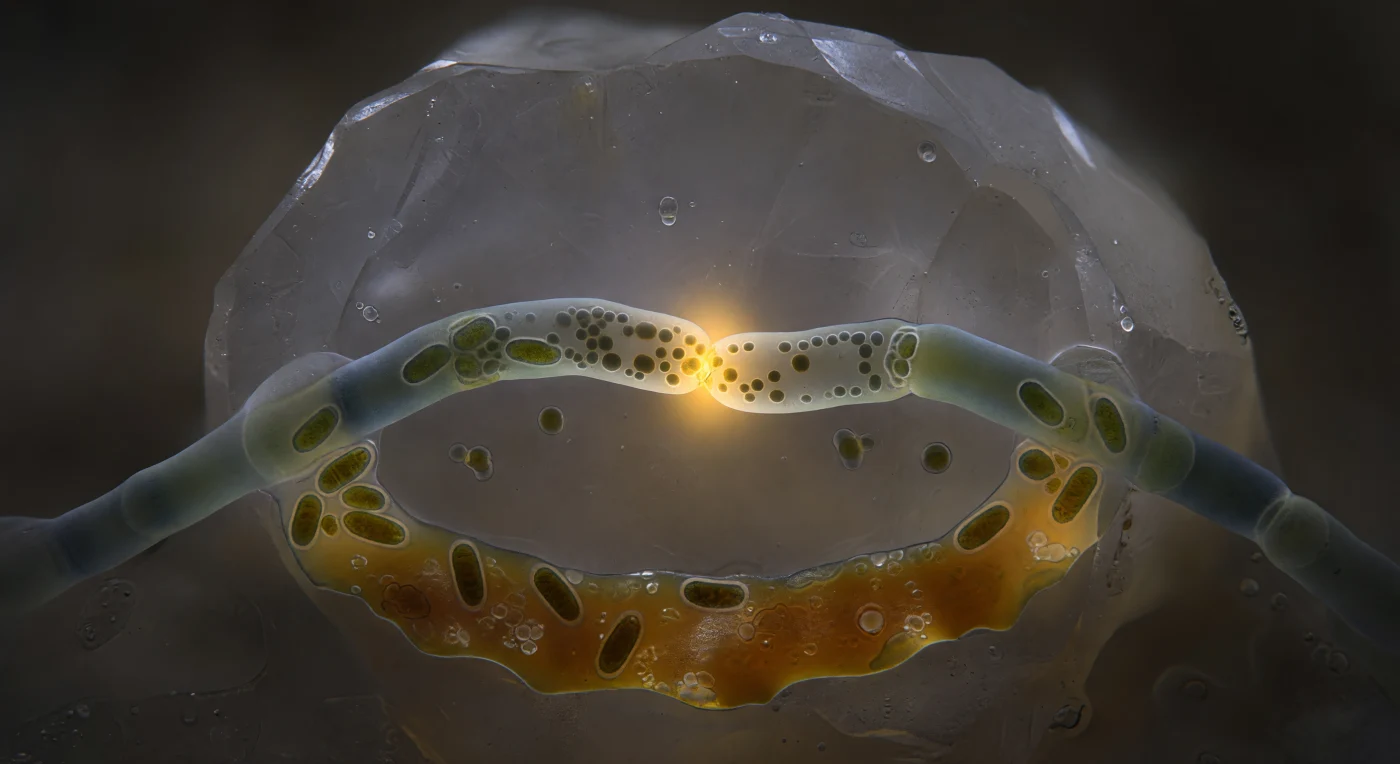
In questo istante sospeso all'interno di un poro del suolo largo una quarantina di micrometri, l'occhio viene catturato da un evento straordinario: due ife di fungo micorrizico arbuscolare si curvano l'una verso l'altra come cavi tesi da una forza invisibile, le loro pareti di chitina e glucani — traslucide come vetro marino appannato, appena tinte di blu-verde — che si dissolvono nel punto di contatto per formare un unico poro di fusione di appena un micrometro e mezzo di diametro, ardente di un ambra-oro intenso e pulsante. Questa apertura luminosa, l'unica fonte di calore cromatico in un mondo altrimenti assoluto nel buio, rivela al suo interno un traffico biologico serrato: mitocondri dalla forma allungata, densi e ramati, si accalcano ai margini del poro mentre granuli lipidici e flussi di citoplasma scivolano in una corda viscosa dall'ifa sinistra verso quella destra, la densità del materiale che cambia percettibilmente nell'attraversamento, come merce scaricata in un porto sottomarino. Sullo sfondo, la parete cristallina di un grano di feldspato sale come una scogliera levigata dai ghiacciai — pallida, appena traslucente agli spigoli, fredda di una luce interna diffusa che non viene da nessuna sorgente esterna — mentre il film d'acqua che riveste ogni superficie cattura il bagliore ambrato del punto di fusione e lo disperde in un'aureola morbida nel grigio-bruno del suolo circostante, trasformando questa anastomosi cellulare in qualcosa che assomiglia meno a un campione di laboratorio e più a una sorgente idrotermale oceanica vista da vicinissimo: un nodo irreducibile di energia e scambio biologico nell'oscurità assoluta della terra viva.

Ciò che riempie l'intero campo visivo è un mondo a sé: un singolo aggregato di suolo occupa l'inquadratura come un pianeta oscuro, la sua superficie di materia organica umificata e melanine fungine compressa in una geologia antica color espresso, punteggiata da inclusioni minerali — granuli di quarzo e feldspato — che affiorano come blocchi di ghiaccio pallido immersi nell'humus scuro. Una pellicola di glomalina riveste ogni faccia esposta in una lacca ambra-dorata sottilissima, conferendo all'intera superficie un lieve carattere idrofobico che si manifesta in una goccia d'acqua perfettamente sferica, quasi mercuriale, appoggiata su un piano anteriore con un angolo di contatto ripido e orgoglioso. Dalle fessure che attraversano la cintura equatoriale dell'aggregato emergono filamenti ipeali bianchi — più sottili del vetro filato a questa scala — che si arcuano in parallelo come punti di sutura di seta attraverso cuoio scuro, tenendo coesa la struttura contro le forze di disgregazione fisica che altrimenti sbriciolerebbero questo mondo compresso. La glomalina non è solo un rivestimento estetico: è una glicoproteina recalcitrante prodotta dalle ife delle micorrize arbuscolari, capace di sequestrare carbonio per decenni e di cementare particelle minerali e organiche in aggregati stabili che resistono all'erosione e sostengono la porosità del suolo — nell'angolo in basso a destra, sotto luce ultravioletta, tutta questa architettura invisibile esplode in fluorescenza giallo-verde incandescente, tracciando ogni contorno e ogni cresta come una mappa di fuoco freddo su un vuoto che sembra spazio cosmico.
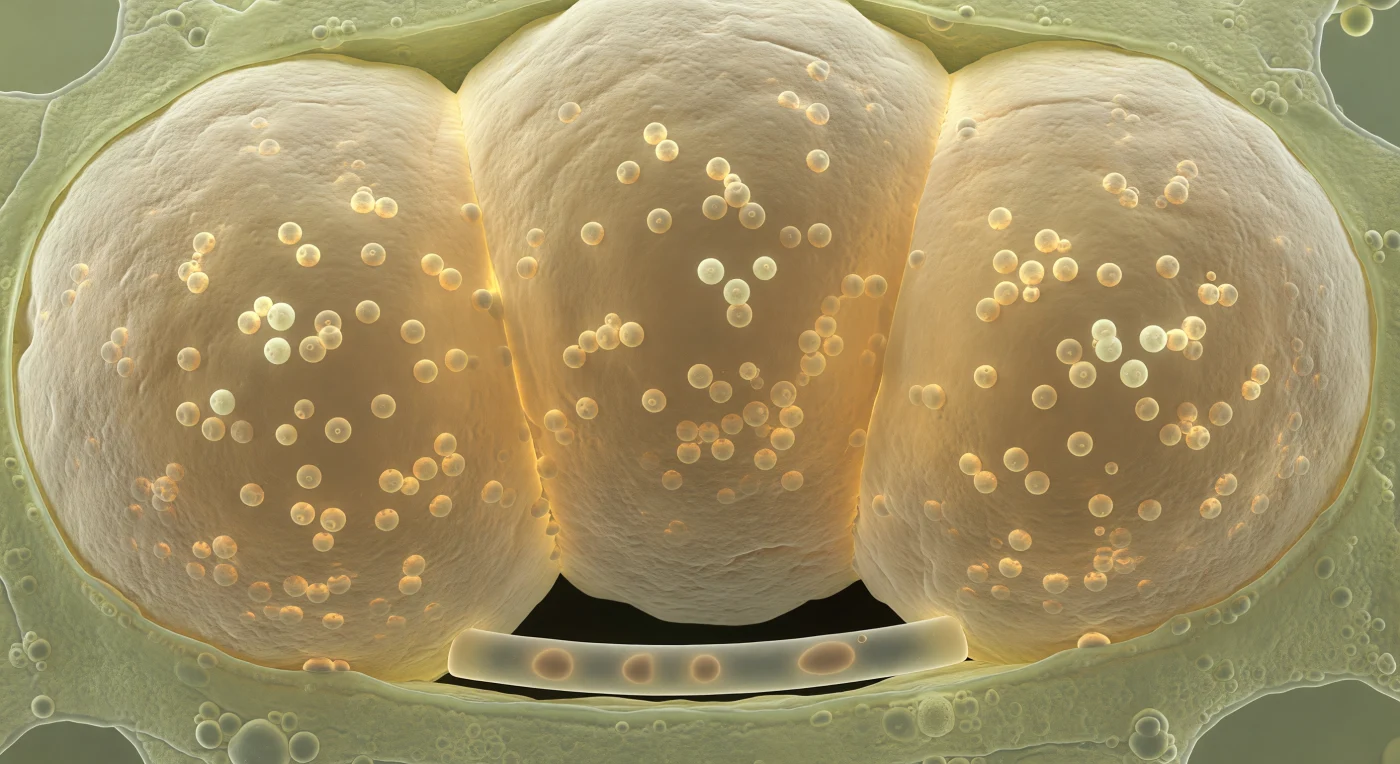
Sospeso all'interno di questa cellula corticale radicale, sei circondato da tre vesciole di funghi micorrizici arbuscolari che riempiono ogni centimetro dello spazio disponibile come palloni aerostatici di ambra pallida gonfiati oltre il limite — la loro superficie esterna, ruvida e stratificata come avorio consumato, transisce verso una levigatezza vitrea nell'interno, luminosa di una luce che non proviene da nessuna fonte esterna ma sembra emergere dalla chimica stessa delle membrane compresse. Queste strutture sono organi di accumulo lipidico: serbatoi temporanei in cui il fungo concentra trigliceridi e acidi grassi sintetizzati a partire dal carbonio fotosintetico ceduto dalla pianta ospite, riserve energetiche dense che alimenteranno la crescita del micelio esterno nel suolo. All'interno di ciascuna vesciola fluttuano decine di globuli lipidici sferici — cinque, dieci micrometri di diametro, ciascuno che rifrange la luminescenza ambrata in un proprio alone dorato, come gocce di olio freddo sospese in resina e congelate a metà caduta, alcune che si sfiorano, altre leggermente separate in un mezzo di viscosità appena percettibile. Il citoplasma della cellula ospite è stato schiacciato ai margini come cera liquida sul bordo di uno stampo, ridotto a una pellicola traslucida e granulare di ribosomi e organelli, mentre la parete cellulare — un reticolo crema-verdastra di microfibrille di cellulosa — preme in silenzio con la tensione quieta di una struttura sotto carico. Quasi invisibile nella penombra tra due vesciole, un singolo ifa intraradicale largo cinque micrometri — non più spesso del tuo polso — si allunga nel buio assoluto oltre la cellula, unico filamento che connette questo vault di lipidi a tutto l'esterno.
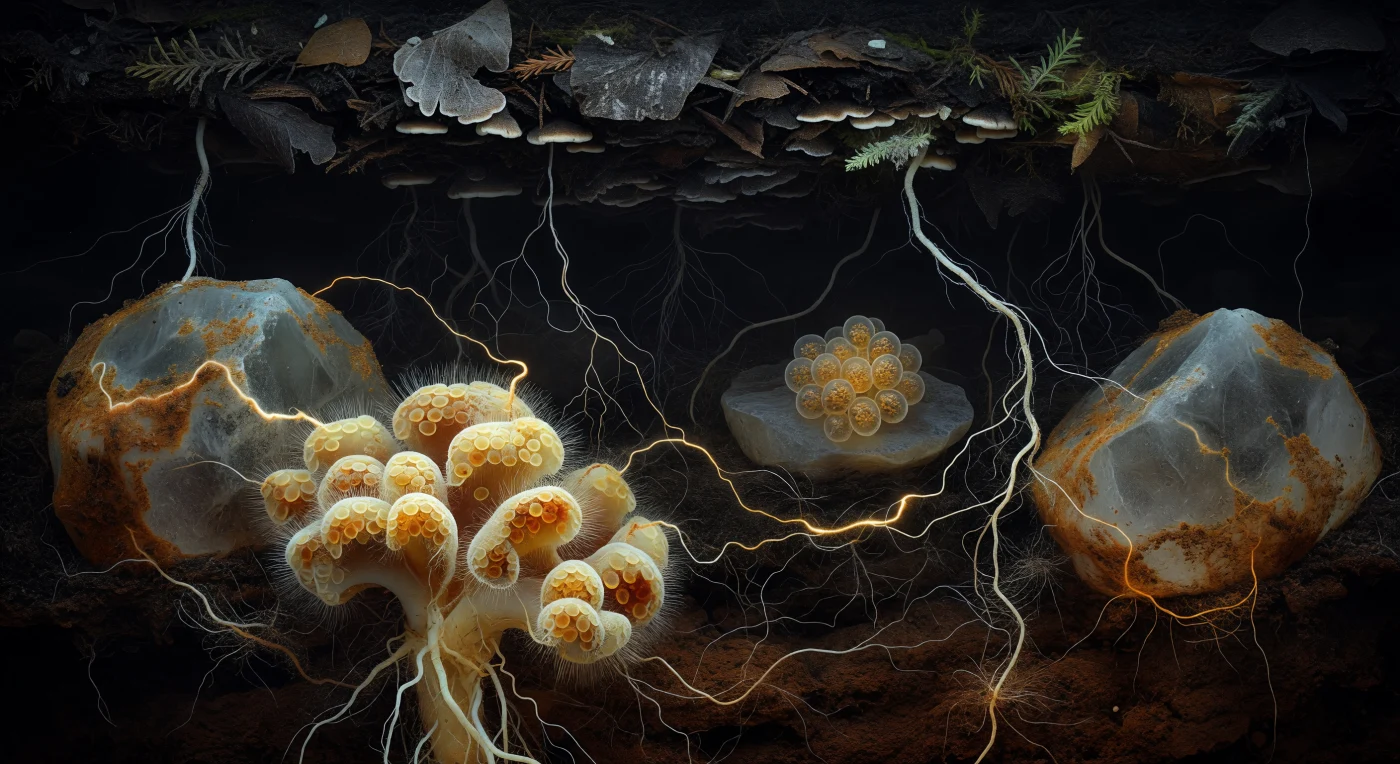
Sospesi nell'oscurità assoluta di un poro del suolo forestale, ci troviamo a derivare tra due massicci granuli di quarzo — le loro superfici incrostate di una patina ambrata di materia organica che sembra trattenere un calore antico, simile alla resina vecchia di secoli. In primo piano, un cluster di punte radicali ectomicorrizali domina la scena come una formazione corallina barocca: il mantello fungino avvolge la radice in strati di crema, giallo burro e ocra arrugginita, con ife extraradicali che si irradiano verso l'esterno come capelli in una corrente lenta, mentre l'argento pallido della corteccia radicale affiora negli interstizi tra gli involucri fungini. Nella mezzadistanza, la Rete Micotica Comune — la cosiddetta Wood Wide Web — si rivela come la trama definitiva di questo mondo sotterraneo: fili bianchi impalpabili, ciascuno non più spesso di un tratto di penna, tessono una maglia tridimensionale attraverso la matrice del suolo oscuro, trasportando zuccheri fotosintetici da albero ad albero in un flusso citoplasmatico appena percettibile come una luminescenza ambrata interna. Un ammasso di spore color ambra riposa contro una superficie di feldspato come uova di vetro traslucido, i loro globuli lipidici che rifrangono la luce disponibile in oro mieloso, mentre le radici fini di faggio e abete rosso scendono dall'orizzonte di humus sovrastante come cavi d'argento e crema, uniti da questa rete gossamer che svanisce progressivamente nell'argilla scura e minerale del sottosuolo.

Sospesi nell'oscurità assoluta del suolo, ci troviamo di fronte alla sezione trasversale di un rizomorfo ectomicorrizale come davanti alla facciata tagliata di un pilastro sepolto: il bordo esterno è un anello di cellule melanizzate così compresse e ispessite da sembrare ossidiana carbonizzata, un'armatura biologica che assorbe ogni fotone e protegge la struttura dai patogeni e dalla disidratazione. Attraversando quella soglia, il passaggio verso la midolla è netto come un confine architettonico — si entra in una cattedrale di cellule-tubo pallide e a parete sottilissima, i cui lumi larghi fino a cinquanta micrometri appaiono come finestre traslucide, i vacuoli centrali pieni di una luminescenza diffusa simile a carta di riso bagnata illuminata dall'interno. Al centro dell'intera struttura si apre il canale assiale, un tunnel cavo e leggermente umido che corre nell'asse del rizomorfo, superficie interna madida e riflettente, vero corridoio di trasporto idrico capace di muovere acqua e soluti attraverso la rete del bosco. All'esterno, ife satellite di pochi micron si irradiano nella matrice di suolo compresso, scivolando tra grani di quarzo — che premono contro il rizomorfo come ciottoli contro un cavo interrato — e film organici ambrati che rivestono i minerali: fili di fibra ottica biologica che svaniscono nell'oscurità terrestre, portando carbonio, fosforo e segnali chimici tra alberi lontani.

Nel buio assoluto della matrice del suolo, un fronte miceliare di funghi micorrizici arbuscolari avanza come un delta fluviale vivente: dall'ifa centrale corrono ramificazioni sempre più sottili, le punte più giovani opache e densamente citoplasmatiche, luminescenti di un bianco-azzurro spettrale che si intensifica nei punti di costrizione dove il filamento si insinua a forza tra due granuli minerali, deformandosi in sezione ovale sotto la pressione della roccia. I segmenti più maturi assumono l'aspetto di catene di perle di vetro torbido, le vacuole lipidiche che si ripetono ogni venti-quaranta micrometri come nodi lungo una corda, mentre aloni arancio-giallastri di ossalato si allargano sulle superfici calcitiche dove la chimica dissolve lentamente il minerale — un calore sulfureo che contrasta con il freddo biologico delle ife. Sullo sfondo, appena intuita attraverso gli strati di particelle e film organici, la parete curva di un pelo radicale si erge come una scogliera pallida di cellulosa, enormemente più grande di tutto ciò che la circonda, destinazione silenziosa che orienta ogni singola punta crescente con un'urgenza che non ha bisogno di frecce per essere percepita.
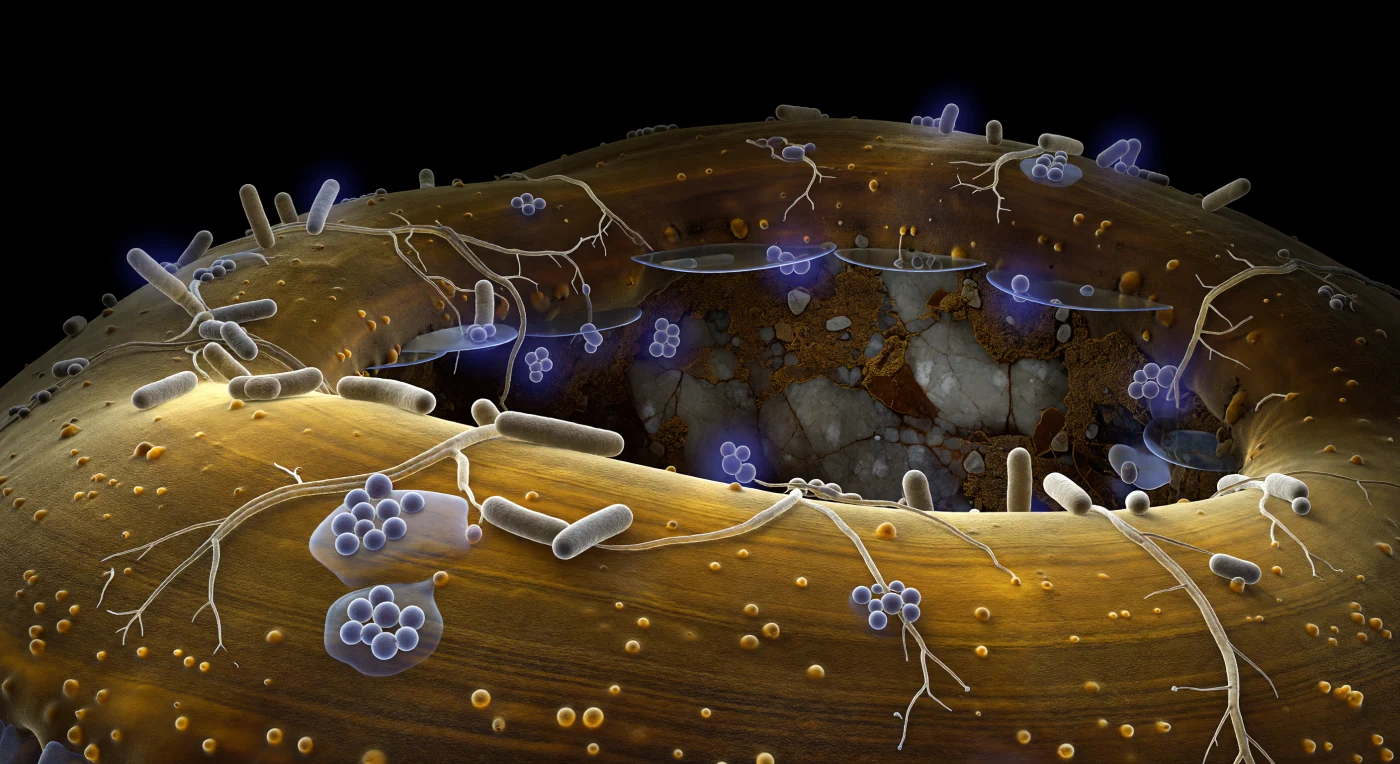
La superficie della ipha ectomicorrizale si distende davanti come il fianco di una scogliera di resina ambrata, incurvata e seminascente, la sua parete chitinosa retroilluminata dal freddo bagliore citoplasmatico che filtra dall'interno come luce attraverso il guscio di una tartaruga levigata — unica fonte di calore luminoso in questa notte permanente sotto il pavimento della foresta. La texture della parete è finemente granulare a questa distanza, stratificata in bande laminari di ambra più scura e più chiara, punteggiata di protuberanze glicoproteiche arrotondate come bolle di resina solidificata, e su questa topografia collinare si aggrappano e si distendono le forme batteriche: cilindri grigio-beige dal lucore ceroso, tre volte la tua altezza apparente, alcuni isolati e eretti, altri riuniti in piccole colonie dove un gel di esopolisaccaridi traslucido si accumula tra le cellule come glicerina versata, rifrangendo il bagliore ambientale in aloni prismatici. Sottili filamenti di Streptomyces ramificati tracciano reti irregolari sulla superficie dell'ipha, ancorati da trefoli adesivi quasi invisibili che appaiono solo come brevi lampi gelatinosi quando la luce li coglie nell'angolo giusto; alcuni individui irradiano aloni violetti in falso colore — molecole segnale che diffondono verso l'esterno in gradienti sferici entro pochi corpi batterici di distanza, facendoli apparire come fioche lanterne tenute vicino alla scogliera ambrata. Oltre la curvatura dell'ipha, il poro del suolo si apre nell'oscurità assoluta, la sua parete lontana una superficie di feldspato fratturato con guizzi micacei, e tra i due mondi sottili pellicole d'acqua tracciano menischi a specchio che catturano il riflesso citoplasmatico attraverso il vuoto — tutta la misura e tutta la scala si percepiscono unicamente attraverso i corpi viventi, la biologia che si legge come geologia.

Nell'oscurità assoluta di una crepa nel suolo, il seme di *Dactylorhiza* si staglia come un dirigibile traslucido sospeso contro pareti di granuli neri di basalto e quarzo — la sua cuticola è un'unica membrana di iridescenza spettrale, sottile come un film di sapone, che rifrange la chimica impercettibile dell'acqua interstiziale in argento pallido e verde-azzurro, mentre all'interno l'embrione appare come un insieme di cellule incolori e quasi sferiche, ciascuna debolmente luminosa come una lanterna di carta, un organismo fermo sulla soglia della possibilità. L'ifa di *Rhizoctonia* che si avvicina da sinistra è, a questa scala, un cilindro imponente color oro-ambra — la sua parete di chitina spessa come bambù laccato, il citoplasma percorso da lenti fiumi di organuli granulari — e dove ha perforato la cuticola la membrana si raccoglie in un annulo serrato, sigillata dal tessuto vivente stesso. Dentro una cellula embrionale il pelotone riempie l'intero volume cellulare: una spirale incredibilmente fitta di ifa avvolta su se stessa decine di volte, ambra e oro e avorio pallido dove i giri si comprimono, illuminata dall'interno da una luce chimica e graduale che è insieme invasione e alleanza, la cellula stessa deformata in una lieve ellisse dalla pressione del groviglio. Le cellule embrionali adiacenti si stanno già modificando — leggermente dilatate, le membrane tese, la loro luminosità interiore che vira dal bianco-argento freddo verso un crema-dorato più caldo mentre la differenziazione muove i suoi primissimi passi molecolari. La palette dell'intera scena è prima di tutto buio, poi ambra: il grit minerale quasi nero di umidità, i film d'acqua pewter-bluastri, il seme come un'aureola argentea fantasmatica, e dentro tutto questo il filo ocra-dorato del fungo che arde lentamente come un carbone nell'oscurità assoluta.

Sospesi all'interno di un poro del suolo, a soli cinque micrometri di altezza, ci troviamo immersi in una cattedrale di oscurità quasi totale, rischiarata soltanto dal bagliore ambrato della glomalina che lacca ogni superficie minerale come una resina di miele antico. Sopra di noi, la corteccia radicale dell'albero appena morto si sfascia in rovine di cellulosa bruna e nerastra, liberando in caduta lenta spore AMF che alla nostra scala appaiono come massi e piccole case — sferiche, ellissoidali, dai toni ambra, rosso vino e ocra — alcune già incrinate lungo le suture, che trasudano fluidi lipidici nel film acquoso circostante. Attraverso l'intera profondità della scena corrono orizzontalmente i fili argentei della rete miceliale comune, cavi iphal ancora percorsi da un tenue flusso citoplasmatico che trasporta risorse verso le radici dei sopravvissuti lontani, mentre sopra di essi il micelio saprotrofico colonizza i resti corticali con ife opache e quasi doppie in diametro, urgenti e ramificate, impegnate nella demolizione biochimica della materia organica. Da sinistra avanza un velo di filamenti giallo pallido, da destra una rete crema più densa, entrambe convergendo su questa zona ricca di nutrienti in una competizione silenziosa e millimetrica che ridisegnerà, nei prossimi giorni e settimane, l'intera architettura del suolo forestale sottostante.
