Wissenschaftliche Zuverlässigkeit: Hoch

Der Blick durch das Herz einer *Salpa thompsoni*-Blüte eröffnet eine Welt, die jeder vertrauten Vorstellung vom Ozean widersteht: Hundert Individuen pro Kubikmeter füllen jeden Sichtstrahl mit pulsierenden Glaszylinder, deren Gewebe so nah am Brechungsindex des Meerwassers liegt, dass sie weniger als feste Körper erscheinen denn als Verdichtungen des Wassers selbst, jede mit einem warm-bernsteinfarbenen Eingeweideschimmer im Zentrum, der das gesamte Volumen in eine perlmuttfarbene, diffuse Leuchte verwandelt. Das kalte, blaugraue Licht der subantarktischen Oberfläche verliert jede Richtung, bevor es wenige Meter weit reicht – gestreut von Hunderten durchsichtiger Linsen, bis die Ferne zu einem gleichmäßigen, nacreous Nebel verschwimmt, aus dem keine einzelne Form mehr heraustritt. Durch dieses Gewebe aus lebendem Glas fällt beständig, mit ballistischer Ruhe, ein Regen dunkeloliv-brauner Kotpillen: dichte, membranbewehrte Zylinder, jedes ein komprimiertes Paket aus umgewandelter Biomasse, das die Energie der Oberfläche mit hundert Metern täglich in die Tiefe trägt. Die Blüte filtert und pulst und atmet unablässig um Sie herum, gleichgültig und endlos, ihr kollektiver Stoffwechsel ein planetarer Kreislauf in langsamer Bewegung – der gesamte Ozean hier verwandelt in ein Organ des Kohlenstoffexports.

In absoluter ozeanischer Schwärze hängt vor dir eine Struktur, die kaum existiert – eine drei Zentimeter große Kugel aus Muzopolysacchariden, so nah am optischen Nichts, dass sie sich nur als eine kaum wahrnehmbare Verzerrung der Dunkelheit abzeichnet, ein linsenartiges Verbiegen der Leere. Dann trifft sie der schräge Lichtstrahl: Die kaltweiße Lichtbahn streift die Membran des frisch aufgeblasenen Gehäuses von *Oikopleura dioica* unter flachem Winkel, und die gesamte Oberfläche entfacht sich in arktisches Blau, Bernsteingelb und Meeresgrün – Interferenzbänder aus Muzinschichten von kaum einem Mikrometer Dicke, die sich wie Seifenblasenirisieren über die Wölbung verschieben, als wäre Licht direkt auf Vakuum gemalt. Dahinter, in der abgedunkelten Innenkammer, pulsiert ein diffus rötlicher Herzschlag durch die Rumpfwand des Larvaceens, während der glasartige Notochordstab in langsamen sinusförmigen Schwingungen durch die transparente Muskulatur des Schwanzes gleitet und die gesamte Strömungsmaschinerie des Gehäuses antreibt. Was zunächst wie leerer Raum zwischen Körper und Außenmembran wirkt, ist in Wahrheit eine architektonierte Welt aus gossamer-feinen Schleimfiltern, deren Porengitter weit unterhalb der optischen Auflösung liegen, deren kollektive Streuung jedoch einen opaleszenten Dunst im Filtrationsraum erzeugt – eine Nanokonstruktion in Gel, für einen Herzschlag lang lesbar im Licht, bevor die Dunkelheit sie zurückfordert.

Vor dir hängt eine lebende Kette aus *Thalia democratica* im offenen Blau, jedes Einzeltier ein gläsernes Fässchen von der Größe einer großen Weintraube, durch dessen wasserhelle Wand du die gesamte innere Architektur auflösen kannst: acht geisterhafte Muskelreifen, ein aufgerollter bernsteinfarbener Darm, der das Licht eines Morgens voller Phytoplankton-Filterung in sich trägt, rosafarbene Gonaden wie winzige Rosenquarztropfen und ein kirschrotes Herz, das sichtbar pulsiert. Das Gewebe dieser Tunikaten besitzt einen Brechungsindex, der kaum von jenem des umgebenden Meerwassers abweicht – die Körper sind buchstäblich aus dem Ozean selbst gemacht, zu neunzig bis siebundneunzig Prozent Wasser, und ihre Sichtbarkeit verdankt sich einzig den pigmentierten Organen und dem Spiel der Sonnenkaustics, die von fünfzehn Metern Wassersäule in geometrische, blau-weiße Netzwerke umgeformt werden und unablässig über die Gallertoberflächen wandern. Jede Verbindung zwischen den Einzeltieren besteht aus stolonischem Gewebe, kaum dicker als ein Spinnenfaden, und doch bildet diese gesamte Struktur eine kollektive Superorganismus-Einheit, die im oligotrophen Freiwasser Picoplankton-Zellen von weniger als zwei Mikrometern Durchmesser filtert – Organismen, die den meisten anderen Zooplanktern schlicht entgehen. Im absolut gesättigten Kobaltblau ohne Boden, ohne Substrat, ohne jede Grenze fühlt sich diese Glaskette wie die einzige Ordnung in einem ansonsten vollkommenen Nichts an.

Der Blick nach oben aus fünf Metern Tiefe öffnet sich in ein Gewölbe aus tiefem Indigo, das letzte gefilterte Tageslicht zu einem schmalen blauen Fenster zusammengedrängt, das die absteigende Struktur von unten wie eine erlöschende Laterne umrahmt. Das verlassene Larvaceen-Haus – eben noch eine transparente Schleimkugel von vielleicht vierzig Zentimetern Durchmesser – knickt und faltet sich in langsamen, eleganten Plissees, während der innere Druck nachlässt: eine Hemisphäre kollabiert nach innen, die andere hält noch ihre Kugelgeometrie, und wo die Membran gespannt bleibt, zerbricht sie das schräg einfallende Licht in irisierenden Bändern aus Violett, Gold und Türkis, während die mit eingeschlossenen Partikeln beladenen Zonen zu cremig-ockerfarbenem Opak verblassen wie beschlagenes Glas. Ein diffraktiver Lichtkranz aus blauweißem Schimmer umringt den gesamten Rand des sinkenden Hauses, dort wo die Membran die letzten Indigostrahlen bricht und streut – ein kurzzeitiger optischer Heiligenschein, der die Struktur wie einen versinkenden Mond erscheinen lässt. Am oberen Einlassgitter, einem durchhängenden hexagonalen Netz aus Mukopolysaccharid-Fäden, schweben bernsteinfarbene Copepoden von kaum einem Millimeter Länge, ihre Körper so durchsichtig, dass das Blau des Wassers durch sie hindurchscheint, nur die kupferfarbenen Komplexaugen und der dunkle Darmkanal verraten ihre Anwesenheit, während sie mit unsichtbar feinen Mundgliedmaßen die verstopfte Filtermembran abtasten und das kohlenstoffreiche Wrack auf seinem Weg in die Dunkelheit begleiten.

Der Blick geht in alle Richtungen gleichzeitig — ein orthogonales Gitter aus schimmernden Gelsträngen, das sich in warmes Bernsteinlicht auflöst, als hätte jemand das Innere einer Kathedrale aus flüssigem Honig gesponnen. Diese Fasern sind das Schleimfilternetz einer Salpe, sekretiert vom Endostyl aus Muzopolysacchariden, einem biologischen Polymer, das Wassermoleküle bindet und dabei eine Maschenweite von kaum einem Mikrometer erzeugt — präzise genug, um Bakterien und Picophytoplankton aus dem vorbeiströmenden Meerwasser herauszusieben. An einer Kreuzung zweier Stränge sitzt eine einzelne *Prochlorococcus*-Zelle wie ein dunkelroter Edelstein im Gitterwerk, ihr Durchmesser von 0,6 Mikrometern lässt sie hier wie einen Felsbrocken wirken, während ein stäbchenförmiges Bakterium träge an einer benachbarten Faser haftet, gehalten durch Oberflächenchemie allein. Das gesamte Netz bewegt sich kaum, doch ein unsichtbarer Strom zieht Partikel unaufhörlich durch diese leuchtende Geometrie — ein stiller, kontinuierlicher Erntevorgang, der tief im Ozean tonnenweise Kohlenstoff aus dem Oberflächenwasser in die Tiefe transferiert.
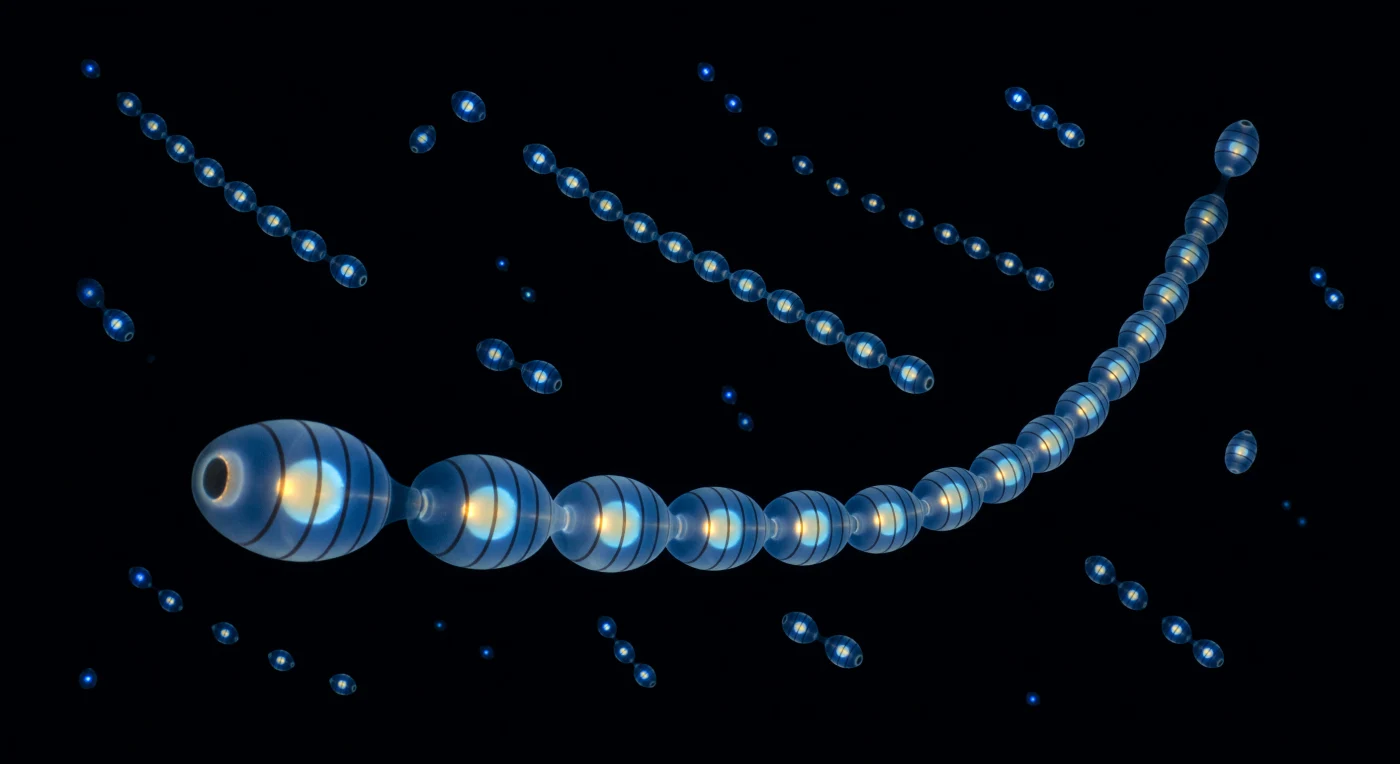
Zwei Meter unter der Oberfläche eines mitternächtlichen Meeres existiert keine Welt mehr – nur Schwärze mit physischer Präsenz –, bis die Kette sich entzündet: Eine Salpen-Aggregation aus fünfundzwanzig Zoiden erstreckt sich als lebender Lüster quer durch das gesamte Gesichtsfeld, jeder Einzelorganismus ein glaserner Zylinder von Daumenbreite, und der biolumineszente Impuls wandert in einem kalten, sequenziellen Blau-Weiß-Wellenzug von Vorder- zu Hinterglied, jeder 300-Millisekunden-Blitz das eigene Innenleben enthüllend – acht dunkle Muskelreifen wie Rippenschatten in einer Laterne, der Darm als warm-goldene Silhouette gegen das chirurgisch reine 476-Nanometer-Cyan, ein Glutkern versiegelt in Eis. Die Körperwand ist keine Oberfläche, sondern ein Medium aus Gel und Wasser und Beinahe-Nichts, sodass das Licht nicht reflektiert, sondern aus der Substanz des Tieres selbst herausstrahlt, als wäre jeder Zoid ein selbstgenügsamer kalter Stern. Im Raumtiefenfeld dahinter pulsen weitere Ketten in loser dreidimensionaler Streuung – nahe als perlenschnurartige Fäden mit erkennbarer Struktur, mittlere als leuchtende Gebilde, ferne als bloße Funken, die vom Salzwasser verschluckt werden, bevor ihr Licht zurückkehren kann –, und zwischen ihnen herrscht keine Dunstschicht, kein Meeresschnee, nur das vollkommene, unwiderrufliche Schwarz des Ozeans, der alles Licht in sich begräbt.

In dreihundert Metern Tiefe, wo kein Sonnenstrahl mehr eindringt, schwebt vor dir eine Kugel, die der Ozean nicht hätte erschaffen dürfen: sechzig Zentimeter Durchmesser, aus einer einzigen kontinuierlichen Membran aus Mukopolysaccharid geblasen, schimmernd in perlmuttfarbenem Blau und zartem Rosa, wo das seitliche LED-Licht ihre Dünnfilm-Interferenz entzündet. Dabei handelt es sich um das Haus von *Bathochordaeus charon*, einem Riesenappendicularie, der dieses Bauwerk innerhalb weniger Stunden aus eigenen Drüsenzellen sekretiert hat – ein mehrschichtiges Baffle-System aus konzentrisch angeordneten Schleimfiltern, deren Maschenweite bis in den Submikrometerbereich reicht und so selbst kleinste Bakterien aus dem vorbeiströmenden Tiefenwasser siebt. Durch die fast wasserklare Wand hindurch erkennt man den kompakten, bernsteinorangen Rumpf des Tieres im Innern, sein Herz in langsam kreisendem Pulsschlag, und den langen, bandförmigen Schwanz, dessen rhythmische Schläge den Wasserstrom durch das gesamte Filterwerk treiben. Am anterioren Pol drängen sich zögernde Copepoden und Amphipoden an das Einlassgitter – angezogen vom Sog, doch nicht bereit, den Schritt ins Innere zu wagen – und machen so spürbar, wie dieses scheinbar zerbrechliche Gebäude aus Schleim in Wirklichkeit eine der effizientesten Filtrationsmaschinen des mesopelagischen Ozeans ist.

Der Blick richtet sich von der abyssalen Ebene aus senkrecht nach oben in eine Dunkelheit, die jede Vorstellung von Raum auflöscht – nur ein diffuses, bläuliches Schimmern, das keiner Quelle zugeordnet werden kann, modelliert die Konturen blass-knochenfarbener Holothurien an den Bildrändern, halb versunken im grau-beigen biogenen Schlamm aus Foraminiferen-Schalen und Radiolarien-Gespenstern. Aus dieser Schwärze fallen, in spiralförmigen Bahnen und mit einer Geduld, die an geologische Prozesse erinnert, zylindrische Kotpellets von Salpen herab – dunkel olivbraun, ihre Oberflächen rau von lebenden Biofilmen aus Bakterien, die den gepressten Inhalt von Picoplankton noch während des Abstiegs abbauen. Dazwischen treiben kollabierte Salpen-Kadaver, deren einstige Tonnenkörper sich in formlose, halbdurchsichtige Gewebefetzen aufgelöst haben und zarte Aureolen aus gelöstem organischem Material hinter sich herziehen – kaum sichtbare Schleier, die sich wie Atem auf Winterglas in die umgebende Wassersäule auflösen. Diese Pellets sind versiegelte Kohlenstoffpakete, die vor Wochen an der sonnenbeschienenen Oberfläche entstanden und nun hier ankommen, um sich in winzige Einschlagkrater im Sediment einzuschreiben: das terminale Empfangszimmer der biologischen Pumpe, gerendert in einer einzigen kalten Frequenz blauen Lichts.

Die Welt, die sich hier auftut, ist ein dreidimensionales Universum aus rubinroten Lichtpunkten, die in tiefblauem Wasser schweben wie Glut in Saphirglas – jede dieser winzigen leuchtenden Kugeln eine einzelne *Prochlorococcus*-Zelle, deren Chlorophyll das kalte blaue Streulicht der Oberfläche in warmes rotes Eigenlicht verwandelt. In regelmäßigen, fast kristallinen Abständen verteilen sich diese photosynthetischen Prokaryoten durch die gesamte oligotrophe Deckschicht des subtropischen Ozeans, ergänzt von den etwas größeren, orange-rot glühenden Zylindern der *Synechococcus* und den kaum sichtbaren, gläsern-geisterhaften Konturen heterotropher Bakterienstäbchen, die nur durch ihre minimale Brechungsindexdifferenz zum umgebenden Wasser verraten, dass sie existieren. Gelegentlich erscheinen größere Picoeukaryoten als kleine Laternen mit komplexen Chloroplastmembranen – konzentrische grün-goldene Kammern, die die schlichteren Prokaryoten ringsum an Strukturreichtum weit übertreffen. Die Leere täuscht: In jedem Milliliter dieses scheinbar klaren Wassers leben zehntausende photosynthetischer Zellen, die zusammen mehr Kohlenstoff binden als alle Regenwälder der Erde – und genau dieses unsichtbare, grenzenlose Gewimmel bildet die Nahrungsgrundlage für die galertigen Filter jener Organismen, deren hauchzarte Körper wenige Zentimeter über diesem Rubinkosmos durch das Wasser gleiten.

Im Inneren des gallertigen Hauses von *Oikopleura dioica* füllt ein einziges, kaum drei Millimeter breites Gebilde das gesamte Blickfeld: der Schwanz des Tiers, stroboskopisch eingefroren in einer vollkommenen organischen S-Kurve, schwebt wie ein durchsichtiges Glasband in dem von diffusem blaugrünem Meereslicht durchfluteten Innenraum. Entlang der zentralen Achse dieser Kurve zieht sich die Chorda dorsalis als leuchtend heller Stab gestapelter, vakuolisierter Scheibenzellen, von denen jede einzelne wie eine bikonvexe Wasserlinse bricht und das einfallende Licht zu einem kühlen, blauweiß glänzenden Streifen bündelt – flankiert von hauchdünnen Muskellamellen, deren regelmäßige Querstreifung kaum sichtbar, aber geometrisch präzise ist. Die seitlichen Flossenränder lösen sich als einzelne Beugungssäume irisierender Interferenzfarben auf, an der absoluten Grenze optischer Auflösung, und hinter dem gekurvten Schwanz beschreiben winzige Nahrungspartikel leicht verschwommene Kreisbahnen – stille Zeugen der Wirbelschlepper, die jeder Schlag in der Flüssigkeit hinterlässt und die nun langsam in Ruhe verfallen. Man schwebt in einem Organismus, der zu neunzig Prozent aus Wasser besteht und dessen Architektur allein durch Brechungsindexunterschiede sichtbar wird – eine gläserne Maschine im Maßstab des kaum Fassbaren.

Zwei Zentimeter vor dir hängt ein *Pegea confoederata* in vierzig Metern Tiefe des tropischen Atlantiks — acht Zentimeter reines biologisches Glas, dessen Tunikawall so nahe am Brechungsindex des umgebenden Meerwassers liegt, dass er sich nur als zarteste Verdickung des Raums verrät, eine gespannte Gelmeniskus aus gefrorenem Ozean. Das diffuse Mittagslicht von der Oberfläche durchdringt den Körper vollständig und verwandelt ihn in eine lebende Laterne: Der Pharyngealkorb glüht in warmem Bernsteingold, sein Schleimnetze fängt das Licht in honigfarbener Irideszenz auf, während der Endostyl als leuchtend-ockerfarbener Längsstreifen die gesamte Ventralseite durchzieht wie eine Ader aus warmem Harz. Kaum drei Millimeter groß, schlägt das karmesinrote Herz in sichtbaren Systolen — dunkles Arterienrot bei der Kontraktion, zartes Rosa bei der Erschlaffung, jede halbe Sekunde ein vollständiger Wellenzug durch das umgebende Gel. Daneben liegt der Gonadenlappen in sattem Orange-Rosa, seine Oberfläche bei diesem Abstand körnig aufgelöst, einzelne blasse Oocyten als transluzente Kugeln in warmem Gewebe eingebettet wie Perlen in Bernstein — ein vollständiges anatomisches Stillleben, aufgehängt im blauen Nichts des offenen Ozeans und von oben durch den gesamten Himmel des Meeres beleuchtet.
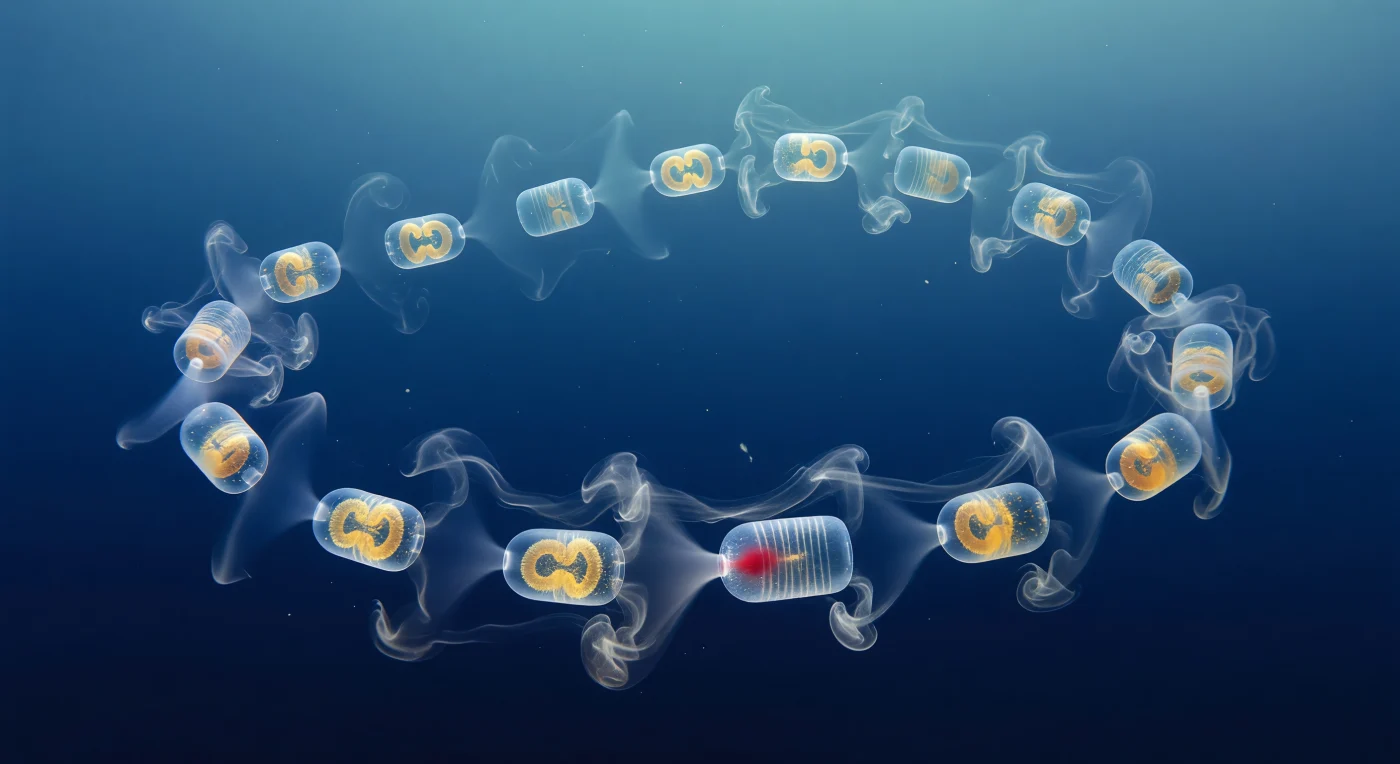
Auf Augenhöhe mit der geschwungenen Kette von *Cyclosalpa affinis* schwebt man mitten im offenen Ozean, umgeben von einem tiefen Kobaltblau, das nach unten ins fast Schwarze übergeht – jeder einzelne Zoid füllt das Blickfeld wie ein bernsteinfarbenes Windlicht aus reinem Glas, seine Wand so transparent, dass das Meerwasser hindurchscheint und sich nur durch einen zarten prismatischen Schimmer an der gewölbten Tunikakante verrät. Die Schlieren-Optik macht das Unsichtbare sichtbar: Aus jedem Atrialsipon quillt ein silbrig-weißer Atemzug warmen Ausatemwassers hervor, dessen Dichtegradient das Licht bricht und ihn wie Morgenrauch erscheinen lässt, der sich in feinen Lamellen auffaltet und nach drei bis fünf Zentimetern mit dem Nachbarstrahl zur geflochtenen Turbulenzwolke verwebt. Jeder Zoid ist zugleich ein Strahlantrieb und ein Filterapparat: Während die Atemzüge nach hinten treiben, zeigen die oral ausgerichteten Einströmsiphonen zarte konkave Schattendellen, wo das partikelreiche Wasser in den Schleimmukosfilter gesogen wird und dabei als schwache optische Linse im Gegenlicht erscheint. Die Kette als Ganzes zieht sich in einem dreidimensionalen Bogen durch das Bild – nahe Zooide scharf und kristallin, entfernte zu blaugrauen Geisterformen verblasst –, während einzelne Marineeschneeflocken träge vorbeitreiben und die lebendige Bernsteinglut der Verdauungsorgane in jeder Kammer still und gleichmäßig pulsiert.

Man schwebt reglos im blauen Nichts des offenen Wassers, eine Handbreite entfernt vom fast körperlosen Rumpf einer *Oikopleura longicauda* — einem durchsichtigen Gelkügelchen, das warme Bernstein- und Rosétöne der inneren Organe wie Einschlüsse in geschliffenem Glas trägt, während der kristalline Schwanz sich in langsamen, hypnotischen Schlägen seitlich ausstreckt. Von der körnigen Oberfläche des Rumpfes drängen in Echtzeit silberweiße Schleimfäden aus den Oikoplastendrüsen hervor, dünner als ein Spinnfaden, die sofort Seewasser aufnehmen und zu einer halbdurchsichtigen Membran aufquellen — dem Rohbau eines neuen Hauses aus Mucopolysacchariden, das sich bereits als schimmernde, unvollständige Ellipse von drei Millimetern Durchmesser abzeichnet. Wo zwei Schleimlagen verschmolzen sind und die Membran die richtige optische Dicke erreicht hat, erscheinen Dünnfilm-Interferenzfarben: ein blassgoldenes Schillern, das sich zu einem Aquamarinton und einem Hauch von Violett wandelt — nicht aufgemalt, sondern physikalisch erzeugt durch dieselbe Wellenlängenauslöschung, die Seifenblasen ihre Palette verleiht. Am Wachstumsrand zerfällt die Struktur in eine Spitze schäumender Mikrobläschen, die jeden Lichtpunkt des diffusen Blaus als Spekularpunkt fangen, während ein kaum sichtbarer Schleier aus gelösten Zuckermolekülen das umgebende Wasser leicht opalisierend trübt — eine lebendige Architektur, Molekül für Molekül errichtet im Herzen des Ozeans.

Der Blick fällt aus wenigen Zentimetern Höhe auf den zusammengesunkenen Kadaver einer Salpe, der wie ein zerknittertes Glasläppchen auf dem Sedimentboden in 1.500 Metern Tiefe liegt – ein schlaffer, blassweißer Gallertbeutel, dessen einstige Ringmuskulatur noch als zarte silbergraue Streifen durch die entleerte Mesoglea schimmert. Unregelmäßige Flecken mattweißen Bakterienbiofilms haben das Gewebe kolonisiert und verwandeln den Kadaver in eine ephemere Nahrungsressource, die in dieser Tiefsee als unverhoffter Energieschub gilt – ein Phänomen, das Wissenschaftler als „Tiefenopportunismus" bezeichnen, da organisches Material aus dem Pelagial selten und unvorhersehbar auf dem Abyssal auftrifft. Drei perlmuttartige Polychaetenwürmer tasten mit federförmigen Tentakelpalpen über die Biofilmflächen, während ein Ophiuridenarm in cremefarbenen Kalzitossikeln vom Sedimentrand herangreift und seine Saugfüße in die Gallerte drückt; fünf glasklare Amphipoden mit leuchtend roten Facettenaugen schaben entlang der Ränder an dem zerfallenden Material. Das umgebende foraminiferendurchsetzte Tiefseeooze aus feinstem biogenem Kalk rahmt die Szene in einer nahezu farblosen blauen Stille ein, in der der bleiche Kadaver wie eine Lichtinsel im ansonsten schwarzen Ozean erscheint und für kurze Zeit ein dicht gedrängtes Gemeinschaftstreffen des abyssalen Lebens ermöglicht.

Du schwebst reglos in fünfzig Metern Tiefe, eingeschlossen in eine Wassersäule, die sich von einem weichen Jadegrün über dir in ein so gesättigtes Mitternachtsblau unter dir verwandelt, dass es jenseits jeder Farbe zu liegen scheint – und zwischen diesen beiden Registern hängen zwei gläserne Lebewesen wie Laternen in einem Dom aus diffusem Licht. Das Larvaceen-Haus zu deiner Linken ist kaum als Körper erkennbar: eine schwebende Seifenfilm-Geometrie aus ellipsoiden Kammern und Einlassfunneln, die das Streulicht nur an ihren gekrümmten Oberflächen als dünne Interferenzfarben – Violett, Gold, ein Geisterhaftes Blaugrün – zurückwirft, während ihr Bewohner, ein glasklarer Tunikat-Rumpf von Traubengröße, mit seinem notochordalen Schwanz zwei- bis dreimal pro Sekunde schlägt und den goldgrünen Schimmer der unsichtbaren Picoplankton-Wolke durch Filtermeschen mit Porweiten von wenigen Hundertstel Millimetern in sich hineinsaugt. Daneben pulsiert eine solitäre Salpe in ruhigem Takt – acht parallele Muskelringe ziehen sich nacheinander zusammen und entlassen einen Wasserstrahl, der den Organismus vorwärtstreibt, während ihr Darm bernsteingolden leuchtet und am hinteren Siphon gerade zwei dichte, membranumhüllte Kotpillen in die Tiefe sinken, jede eine komprimierte Ladung organischen Kohlenstoffs auf dem Weg zum Meeresboden. Weit unter dir löst sich langsam aus dem Indigodunkel eine silbern-weiße Scheibe von der Größe eines Esstisches: die aufsteigende Mola mola, deren kleiner offener Mund genau auf diese glasernen Zwischeninstanzen gerichtet ist und damit die ganze unwahrscheinliche Nahrungskette sichtbar macht – von Cyanobakterien, die kleiner als ein halbes Mikrometer sind, bis zu einem Dreihundert-Kilogramm-Fisch, verbunden allein durch diese durchsichtigen Wesen, die im honigfarbenen Licht hängen wie Gedanken, die das Wasser selbst denkt.
