Confianza científica: Alto

Suspendido en el corazón de la floración, cada línea de visión queda interrumpida antes de los cincuenta centímetros por otro barril de gelatina viva: *Salpa thompsoni*, cien individuos por metro cúbico, convertidos colectivamente en una niebla nacarada que transforma el océano subantártico gris-verde en algo parecido a un enorme fanal de luz opaca. Cada zoide es un cilindro muscular perfectamente transparente —del tamaño de un puño o de un antebrazo— cuyas bandas estriadas solo se revelan como un leve temblor refractivo, mientras el intestino color ámbar-dorado y la gónada rosa coral flotan en su interior como joyas suspendidas, encendidas por la luz difusa que desciende desde arriba como una catedral polar sin dirección precisa. A partir de tres metros los individuos se disuelven en la opalescencia colectiva hasta que el campo lejano se convierte en una niebla nácar impenetrable, punteada únicamente por cientos de pequeños corazones carmesí que laten a dos pulsos por segundo —señales de vida interior en un mundo compuesto casi enteramente de agua y gel. Por debajo del plano horizontal, cruzando el campo visual en procesión continua, caen los pellets fecales: cilindros compactos de color verde oliva oscuro, densos y lastrados, que descienden con propósito balístico a cien metros por día, transportando hacia el abismo gris-azulado el carbono reempaquetado de millones de picoplancton —la floración filtra, pulsa y exhala indiferente arriba mientras su producto más sólido se aleja hacia la oscuridad, una nevada orgánica en sentido inverso que fertiliza el fondo oceánico con los residuos comprimidos de este vasto e inacabable resplandor lechoso.

Suspendido en la oscuridad absoluta del océano profundo, el ojo se encuentra a apenas unos centímetros de una estructura que casi no existe: una esfera de mucopolisacáridos tan perfectamente adaptada ópticamente al agua marina que se percibe únicamente como una leve distorsión del negro, una curvatura fantasmal en el límite de la nada. Entonces el haz oblicuo la encuentra, y la casa entera se enciende en cascadas de color interferencial —azul ártico que se derrama en ámbar dorado que se funde en verde espuma de mar— bandas que se reorganizan y respiran sobre la membrana curva como una pompa de jabón estirada hasta el tamaño de una naranja, pintadas directamente sobre el vacío con una luminosidad tan fina que parece imposible. En el interior de esa penumbra translúcida late un pulso rojizo e insistente —el corazón de la larvácea ardiendo como una brasa vista a través de la niebla— mientras la notocorda vítrea traza sus arcos sinusoidales lentos y casi meditativos, y cada batida de la cola hace temblar los tabiques internos de mucus estructurado que revelan por un instante, antes de que la oscuridad los reclame, la arquitectura nanoscópica de una catedral de gel en pleno funcionamiento. Copos de nieve marina derivan a través del haz en su descenso gravitacional silencioso, y la sensación que lo domina todo es la de una fragilidad imposible ejerciendo una autoridad tranquila: esta burbuja de casi-nada de tres centímetros es una planta filtrante, una bomba de carbono, una vida de cordado completa, y lleva funcionando perfectamente desde hace quinientos millones de años.

Te encuentras inmóvil en columna de agua abierta, a la altura de los ojos con una cadena viva de *Thalia democratica* que se extiende desde pocos centímetros frente a ti hasta un infinito cobalto tan saturado que parece el interior de una estrella azul. Cada zoide de la cadena cuelga en el agua como un barril de vidrio soplado no más ancho que una uva grande — y desde esta proximidad puedes resolver con nitidez toda la arquitectura interna a través de la pared corporal casi invisible: ocho bandas musculares estriadas de blanco fantasmal envuelven cada cilindro como sugerencias de estructura más que tejido sólido, mientras en el centro geométrico un intestino enrollado de ámbar dorado brilla como si contuviera un filamento cálido, el fitoplancton acumulado durante una mañana de filtración continua comprimido en un núcleo luminoso suspendido en gel perfecto. La transparencia de estos animales no es vacío sino física precisa — su índice de refracción se separa del agua de mar por la fracción más mínima, lo suficiente para que la luz solar reestructurada en quince metros de océano proyecte encajes geométricos de azul y blanco sobre sus cuerpos y los atraviese, completando sus patrones en la nada cobalto del más allá. En el polo anterior de cada zoide, un corazón del tamaño de una cabeza de alfiler late con pulso visible, lanzando destellos de carmesí a través de la pared del barril, mientras la cadena entera — conectada por tejido estolónico fino como seda de araña — ocupa apenas el dos por ciento de tu campo visual total, y sin embargo absorbe cada átomo de atención porque es la única estructura organizada en lo que de otro modo sería una oscuridad azul perfecta e infinita.

Desde cinco metros por debajo, la mirada asciende hacia una estructura que llena el campo visual como una medusa fantasma en colapso: la casa larvaciana abandonada desciende girando lentamente, su membrana mucosa —hace apenas horas una esfera perfecta de unos cuarenta centímetros de diámetro— se pliega ahora en crestas asimétricas que atrapan y liberan la luz índigo del crepúsculo mesopelágico en arcos iridiscentes de violeta, oro y verde azulado que viajan por su superficie como colores en una pompa de jabón. Donde las partículas se han acumulado en el interior, la transparencia cede a una opacidad lechosa, crema y ámbar pálido, la huella biológica de cadenas de fitoplancton y pellets fecales que manchan la membrana desde dentro como condensación sobre cristal frío. Un halo difractivo de luz blanco-azulada corona el perímetro completo de la estructura, el borde de la membrana refractando los últimos rayos descendentes en un anillo luminoso espectral que hace que la casa parezca una luna caída hundiéndose lentamente hacia las tinieblas. En la rejilla de entrada, aún visible como una malla geométrica de hilos mucopolisacáridos que se comba bajo el peso del detrito acumulado, copépodos de apenas un milímetro —cuerpos tan transparentes que el agua azul los atraviesa, sus ojos compuestos como dos esferas de cobre cálido— sondean el filtro colmatado mientras la nieve marina, una precipitación de copos de mucus y diatomeas individuales, desciende a velocidades distintas llenando la columna de agua de profundidad y presencia.

Suspendido en la oscilación quieta de la corriente faríngea, el ojo no encuentra paredes ni suelo: solo la malla. En todas las direcciones se despliega una catedral ortogonal de fibras de gel mucopolisacárido, cada hebra del diámetro aproximado de una bacteria, tensa entre sus puntos de anclaje como hilo de vidrio ámbar fundido, bañada en una película viscosa casi invisible que atrapa la luz dorada irradiada desde el tejido de la salpa y la devuelve como fulgor de miel traslúcida. La geometría es implacable en su regularidad: horizontales y verticales se cruzan en ángulo recto creando ventanas de apenas un micrómetro de apertura, filtros biológicos capaces de retener células de *Prochlorococcus* de 0,6 micrómetros —como la esfera carmesí oscura encajada ahora mismo en la intersección más próxima, una roca de color borgoña presa en el entramado— junto a la vara bacteriana de amarillo pálido que adhiere su flanco a la fibra vecina mediante química de superficie. La red se repite en profundidad plano tras plano, cada capa sucesiva un poco más difusa que la anterior, disolviéndose en una niebla cálida de oro y ocre donde la geometría se convierte en pura luminosidad, mientras una corriente invisible y paciente arrastra el agua y su carga de vida celular hacia la digestión de la salpa que lo contiene todo.
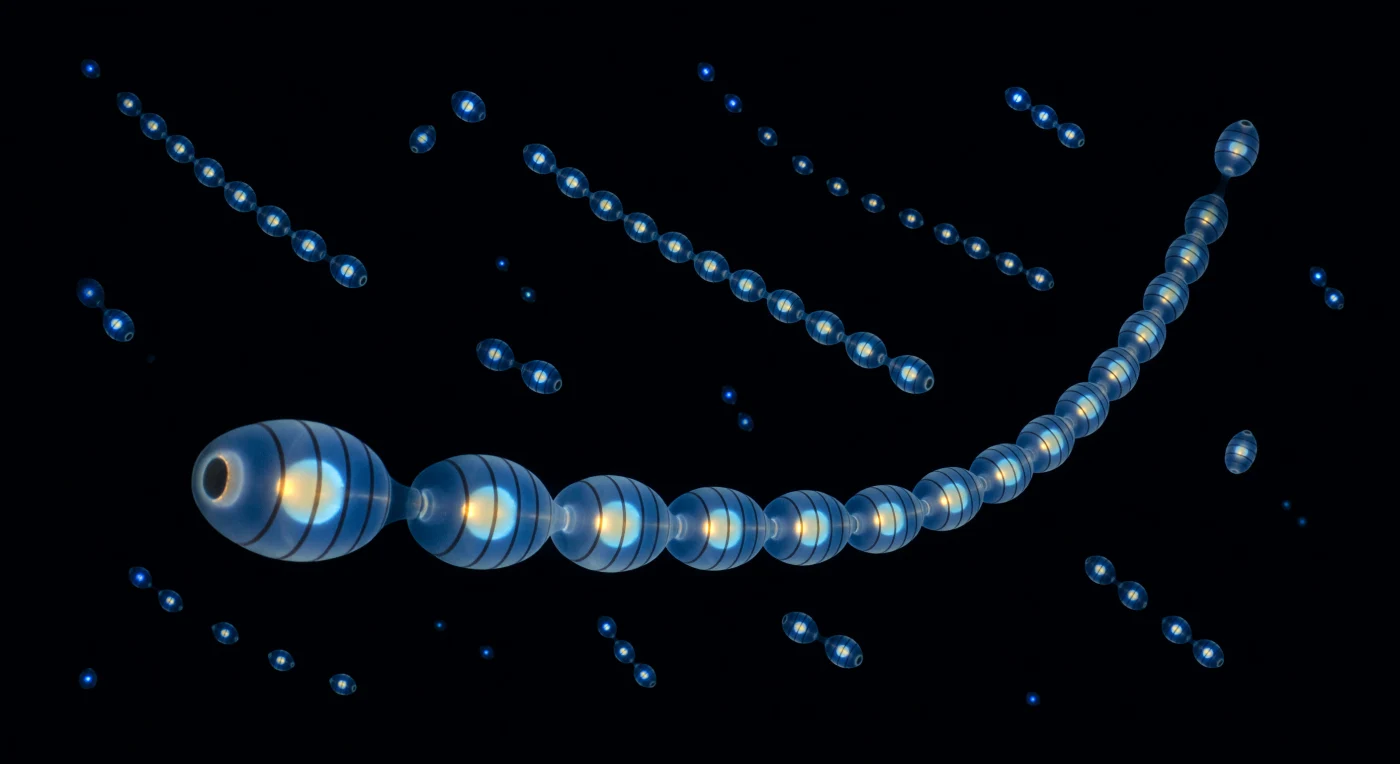
A dos metros bajo la superficie, en una oscuridad tan densa que parece material, la cadena de sálpas estalla en luz. Veinticinco zooides se extienden a lo largo de todo tu campo visual como un candelabro viviente, cada uno del tamaño de un pulgar, y el pulso bioluminiscente recorre la cadena de anterior a posterior en una ola secuencial de azul-blanco frío, encendiéndose y extinguiéndose cada barril en trescientos milisegundos: dentro de cada zooide iluminado, las bandas musculares circulares aparecen como aros oscuros apilados contra el resplandor cerúleo, mientras el contenido intestinal flota como una silueta ámbar suspendida, oro cálido contra azul quirúrgico, una brasa sellada dentro del hielo. La pared corporal no es una superficie sino un medio —gel y agua y casi nada—, de modo que la luz no se refleja sino que irradia desde la propia sustancia del animal, cuyo índice de refracción roza tan de cerca el del agua de mar que el organismo existe menos como objeto que como diferencia: una arquitectura de vacío organizado que se delata únicamente por sus sombras internas y su propio fuego frío a 476 nanómetros. Detrás y alrededor de la cadena más próxima, otras cadenas pulsan en dispersión tridimensional a distancias de uno a varios metros en todas las direcciones, reduciéndose progresivamente desde hilos de cuentas hasta puntos de chispa pura, hasta que las más lejanas son indistinguibles de estrellas; y entre ellas no hay niebla ni difusión, solo la absorción absoluta del agua salada tragándose la luz antes de que pueda regresar, de manera que la profundidad del espacio no se ve sino que se siente, un diagrama de dispersión volumétrico dibujado en fotones fríos contra un vacío perfecto e irreversible.

Ante ti se despliega una esfera de sesenta centímetros que parece labrada en cristal soplado, suspendida en la oscuridad absoluta de las aguas mesopelágicas a trescientos metros de profundidad, donde ningún rayo solar ha llegado jamás: la membrana de mucopolisacáridos atrapa la luz lateral de los LEDs del ROV y la dispersa en nácar iridiscente —blanco hielo, azul pálido, rosa fugaz— gracias a la interferencia de película delgada de una pared biológica estirada hasta un grosor casi imposible. En su interior, visible a través de capas concéntricas de filtros mucosos casi perfectamente igualados al índice de refracción del agua, el tronco ámbar-anaranjado de *Bathochordaeus charon* late con un ritmo lento y poderoso, su notocorda caudal brillando como una varilla de vidrio mientras cada undulación lateral comprime el agua hacia los filtros de alimentación anteriores, generando ondas de presión que recorren la membrana exterior como respiraciones de un pulmón translúcido. Sobre la rejilla de entrada en el polo anterior —hexágonos de mucus de apenas sesenta micrómetros de apertura, cada uno una lente de luz azul-blanca— tres copépodos flotan en suspensión cautelosa, sus ojos compuestos como rubíes oscuros, atraídos por la corriente pero sin atreverse a cruzar el umbral de esa arquitectura efímera y perfecta. Alrededor, en los límites más lejanos del alcance lumínico, zooides solitarios de salpas derivan en peristáltica lenta, cada uno una linterna de biología disuelta contra el frío sin fondo del abismo.

Desde el fondo del abismo, con la mejilla apoyada sobre la ooze biogénica gris-beige a 3.000 metros de profundidad, la mirada se eleva hacia una oscuridad que no tiene techo ni horizonte: una catedral negra y silenciosa cuyo único registro de color es un frío resplandor bioluminiscente de azul casi monocromático, tan tenue que parece el recuerdo de la luz más que la luz misma. En ese vacío aparente desciende una procesión vertical de materia con paciencia geológica: cilindros oscuros de color oliva-marrón de apenas medio milímetro a dos milímetros de longitud —pellets fecales de salpa— giran con lentitud infinita mientras caen, sus superficies rugosas por biofilms bacterianos que les confieren una textura aterciopelada y viva, y sus bordes recogen ese azul ambiental como un halo rimlight casi imperceptible. Entre ellos, carcasas de salpas colapsadas descienden como pañuelos de tejido translúcido, sus bandas musculares ya disueltas, arrastrando halos difusos de materia orgánica disuelta que enturbian el agua a su alrededor como vaho sobre cristal frío. Esta lluvia oscura es la cámara terminal de la bomba biológica oceánica: cada pellet es un paquete sellado de carbono fijado en la zona fótica hace semanas, completando una travesía vertical de miles de metros para inscribirse como un microimpacto sobre el sedimento pálido donde, en los márgenes del encuadre, las siluetas blancas de holoturias permanecen inmóviles, casi lunares, testigos mudos de una nieve que nunca cesa.

Te encuentras suspendido en el corazón de un universo que ningún ojo humano podría alcanzar por sus propios medios: el océano oligotrófico subtropical visto desde adentro, a la distancia de un solo diámetro celular del organismo más cercano. Ante ti, en todas direcciones hasta donde la sapiente oscuridad azul lo permite, una constelación de esferas rubí arde con la lentitud de brasas que nadie avivó —cada una es una célula de *Prochlorococcus*, cargando en su interior los pigmentos que capturan la luz empobrecida que desciende desde cincuenta metros más arriba, y que la devuelven transformada en ese rojo encendido, cálido y obstinado, contra el frío azul del medio. El agua no es oscuridad sino translucencia activa: fotones dispersados por decenas de metros de columna oceánica llegan aquí sin dirección precisa, construyendo una iluminación de catedral sin nave ni vitrales, un azul que lo impregna todo y que hace que cada emisión clorofílica parezca una pequeña hoguera interior. Entre los puntos rojizos más abundantes flotan cilindros levemente más grandes de *Synechococcus*, de un rojo más anaranjado y con un interior levemente granular donde los pigmentos antena —ficobilisomas— distribuyen la energía captada antes de cederla a la fotosíntesis, y más allá, casi invisibles, varillas bacterianas heterotróficas se deslizan en arcos brownianos, denunciadas únicamente por el prisma de su membrana al atravesar el fondo azul. Este es el verdadero motor del océano productivo: no las ballenas, no los peces, sino este cosmos rubí de células que miden menos de un micrómetro y que, en su densidad aparentemente vacía —una cada decenas de micrómetros—, suman la mayor biomasa fotosintética del planeta y sostienen, sin saberlo, todo lo que vendrá después, incluidos los animales gelatinosos que filtran este mismo universo diminuto para construir sus propios cuerpos de agua y luz.

Te encuentras suspendido en el interior líquido de una casa de mucopolisacáridos casi invisible, con el campo visual copado por una sola estructura de tres milímetros: la cola de *Oikopleura dioica* detenida a mitad de batida en una perfecta curva en S, iluminada desde arriba por la luz azul-verde difusa que se filtra a través de las paredes del casa como una luminosidad catedralicia sin sombras duras. La cola se presenta como una cinta de vidrio translúcido cuyo eje central alberga el notocordio, una varilla brillante formada por células vacuoladas apiladas en disco, cada una un lente biconvexo de citoplasma comprimido que concentra la luz ambiental en un trazo lineal de blanco-azul frío, flanqueada a ambos lados por delgadas láminas de músculo estriado cuyas bandas transversales se insinúan como líneas de orden disciplinado. Los extremos de la cinta se desvanecen en las membranas alares, cada una reducida a una sola franja de difracción con interferencia violeta que se apaga hacia plata, existiendo en el límite absoluto de la resolución óptica. Detrás de la curva, el agua del compartimento interior no está en reposo: partículas de ámbar y oro describen trayectorias circulares residuales, fantasmas de estelas vorticales que el último batido dejó antes de que la imagen quedara congelada, testimonio visible de que este organismo, apenas mayor que el grosor de una uña, mueve el océano a su alrededor.

Suspendido a dos centímetros de su cara anterior, el *Pegea confoederata* no oculta nada: la túnica —una pared de gel tan próxima al índice de refracción del agua marina que solo se delata como un menisco curvo, una piel de cristal pulido— convierte al organismo entero en una linterna anatómica iluminada por la luminancia difusa del Atlántico tropical filtrada a través de cuarenta metros de columna de agua. En ese interior suspendido flotan las estructuras como inclusiones en ámbar vivo: la cesta faríngea resplandece con un parpadeo dorado-miel donde la red de mucosa —filamentos biológicos de seda secretados por el endostilo— dispersa la luz en iridiscencia cálida mientras filtra bacterias y picoplancton de apenas décimas de micrómetro; el endostilo mismo traza una franja ventral de resina encendida a lo largo de todo el barril, densa y luminosa. A tres milímetros de diámetro, el corazón late en ondas peristálticas perfectamente visibles cada medio segundo —rojo arterial en la sístole, rosa translúcido en la diástole— enviando pulsos de color a través del gel circundante, y junto a él el lóbulo gonadal naranja-rosado exhibe ovocitos individuales como esferas pálidas engastadas en tejido cálido, perlas en ámbar. El océano circundante es un vacío azul-índigo infinito sin referencia sólida, punteado únicamente por copos de nieve marina que derivan a un centímetro de distancia y capturan la luz de fondo como motas brillantes, mientras en el borde curvo de la túnica un destello de interferencia de película delgada —verde pálido, violeta— recuerda que este barril de agua organizada por la vida es, en su perfección geométrica, indistinguible de una burbuja de jabón estirada hasta la transparencia absoluta.
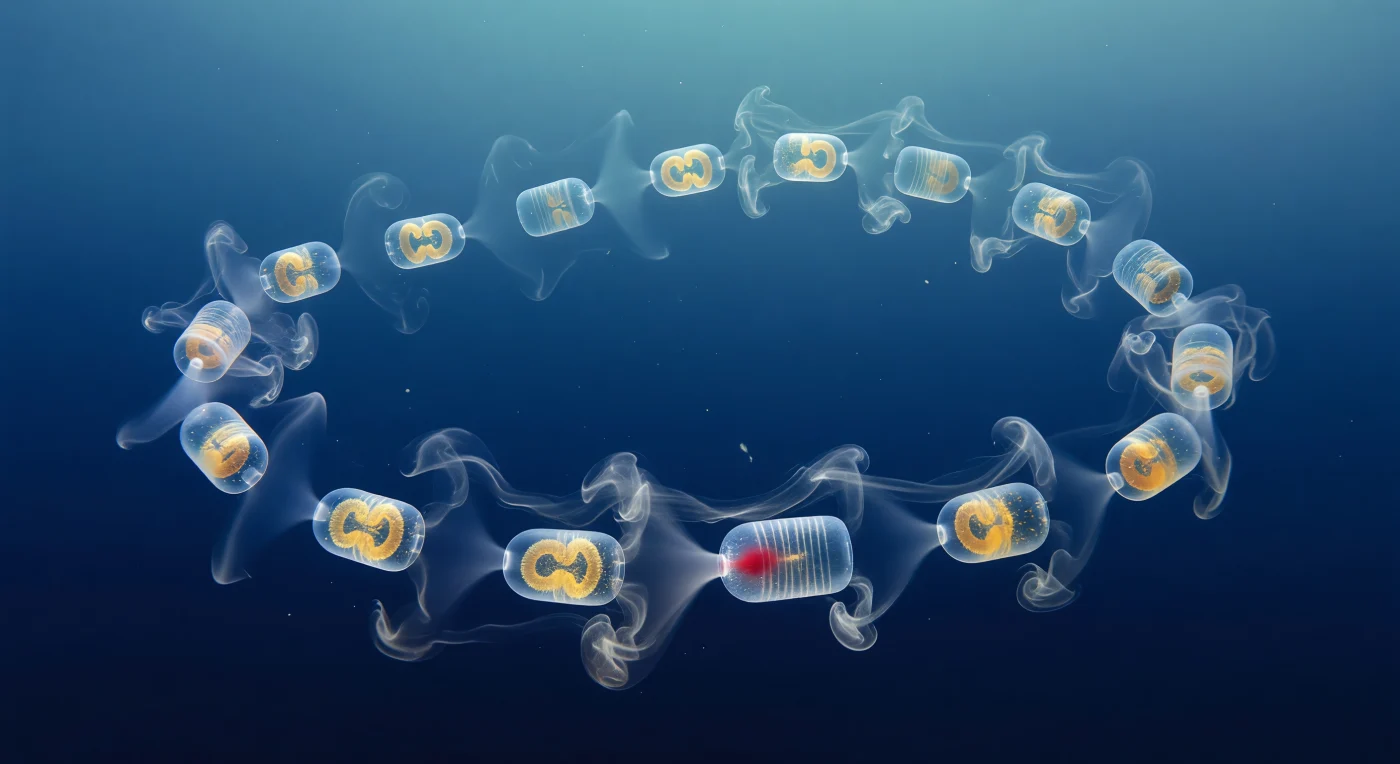
Suspendido frente a ti como una guirnalda de faroles de ámbar en el interior de un océano silencioso, el anillo de *Cyclosalpa affinis* pulsa con una cadencia lenta y casi hipnótica: cada barril cilíndrico, apenas más largo que una falange, deja pasar el azul del agua casi sin alterarlo, traicionando su presencia solo por el halo prismático que dibuja la pared de túnica y por el nudo dorado-incandescente del intestino cargado de fitoplancton que flota dentro como una brasa dentro de un frasco de cristal. La óptica Schlieren convierte el agua misma en un registro visible de la mecánica del fluido: de cada sifón atrial emerge una pluma de agua exhalada ligeramente más cálida y de menor densidad que el entorno, resuelta por los gradientes de índice de refracción como humo de plata que se ensancha, se deshilacha en láminas y se entrelaza con el chorro del zoide vecino para formar una estela compuesta de filamentos trenzados, iridiscentes donde la luz descendente los atraviesa en ángulo oblicuo. En el extremo opuesto de cada barril los sifones orales generan el negativo exacto de ese fenómeno: una sombra cóncava y tenue, una lente invertida de agua levemente más densa que converge hacia la abertura de entrada y delata, solo por diferencia óptica, el mecanismo de filtración que captura bacterias de menos de dos micrómetros sin ninguna estructura rígida. La cadena completa se arquea en una hélice suave a través del campo de visión, los zoidos más cercanos nítidos como vidrio tallado y los más lejanos disueltos en el azul índigo, mientras copos de nieve marina derivan entre ellos como testigos mudos de un proceso que, multiplicado por los millones de individuos de un bloom oceánico, bombea carbono orgánico hacia el fondo a una tasa que los modelos climáticos aún intentan cuantificar con precisión.

A un centímetro del tronco de *Oikopleura longicauda*, el cuerpo del animal se presenta como una esfera de gel casi perfecta, apenas del tamaño de un grano de arroz, suspendida en el azul difuso de la columna de agua: en su interior, un crescent de intestino color ámbar y un núcleo rosado de gónada flotan dentro de lo que parece vidrio viviente, mientras la cola se aleja en un bastón de cristal esmerilado que late con suaves y rítmicas ondulaciones. Desde la superficie del tronco, docenas de células glandulares oikoplásticas extruden en tiempo real filamentos de mucus plateado, cada uno más fino que una telaraña, que al contacto con el agua marina absorben fluido y se hinchan formando membranas semitransparentes: la casa incompleta, de apenas tres milímetros, pende como una burbuja de jabón hecha de materia viva, con su cuarto faltante todavía abierto al océano. Donde dos capas de mucus se han fusionado y adelgazado hasta el espesor óptico preciso, la interferencia de película delgada deposita colores que no están pintados sino tejidos en la física de la luz: un oro pálido que vira a aguamarina y se desvanece en violeta espectral hacia el frente de crecimiento, tonos que se deslizan y reordenan cada vez que la membrana flexiona, igual que el aceite sobre agua inmóvil. En ese frente activo, una encaje de microburbujas de mucus aún sin fusionar captura el azul ambiental como puntos de luz especular, y más allá una tenue opalescencia —mucopolisacáridos disueltos difundiéndose en gradientes brownianos— empaña suavemente el volumen circundante, como si el agua misma hubiera sido levemente escarcha en torno a esta arquitectura molecular que se levanta, filamento a filamento, en el corazón del mar abierto.

A tres centímetros del fondo, el campo visual queda enteramente ocupado por la bolsa desinflada de una salpa muerta, extendida sobre la ooze foraminiferífera como un pañuelo de vidrio arrugado: su mesoglea translúcida ha perdido la turgencia, y las antiguas bandas de músculo circular se adivinan aún como líneas gris-plateadas casi borradas, vestigios de la arquitectura contráctil que en vida impulsaba el animal por propulsión a chorro filtrando picoplancton de menos de dos micrómetros. Manchas de biofilm bacteriano blanco-mate colonizan la membrana glistening, primer eslabón de la cadena de remineralización que convierte este pulso de carbono orgánico —caído desde la zona fótica en semanas de descenso lento— en nutrientes devueltos al ciclo bentónico a 1 500 metros de profundidad. Tres poliquetos de cuerpo nacarado rastrean con palpos tentaculares el tejido en descomposición, un brazo de ofiura articula sus osículos calcáreos desde el borde del sedimento hacia el margen del cadáver, y cinco anfípodos translúcidos —con ojos compuestos de un rojo granate casi fosforescente— raspean la superficie bajo una luz bioluminiscente azul-verdosa difusa que hace de este resto pálido el único foco cálido en la oscuridad fría. La ooze circundante revela, a esta escala íntima, tests individuales de foraminíferos —esferas perforadas y formas espiraladas de calcita— incrustados en un manto de nieve marina sedimentada, recordando que cada gramo de ese lodo es historia comprimida del plancton gelatinoso que alguna vez pobló la columna de agua sobre este abismo silencioso.

Te encuentras suspendido en plena columna de agua a cincuenta metros de profundidad, dentro de un gradiente que asciende desde el azul oscuro más denso del abismo hasta una neblina jade-dorada que difumina la luz solar descendente en conos de resplandor difuso, casi lácteo. A treinta centímetros de tu cara, la casa larvaceana ocupa su propio universo óptico privado: una arquitectura de jabón iridiscente —violeta, oro, verde fantasma— que apenas existe como objeto sólido y sin embargo está construyendo en tiempo real el puente invisible entre picoplancton de medio micrón y tejido animal, atrayendo hacia sus embudos de filtración la turbidez dorada suspendida en el agua como si destilara luz disuelta. Más a la derecha, el salpón solitario pulsa con una contracción lenta y cristalina, su intestino encendido en ámbar cálido, mientras dos pequeñas bolitas oscuras de material fecal comienzan su viaje gravitacional hacia las profundidades —cápsulas densas de carbono empaquetado que transportan hacia el fondo lo que fue primero energía solar—. Y desde ese azul tan saturado que se convierte en oscuridad propia, ascendiendo con lentitud de planeta, un enorme disco plateado del tamaño de una mesa de comedor se aproxima con la boca abierta: el *Mola mola*, cuya improbable cadena alimentaria desde la cianobacteria hasta los trescientos kilos de músculo y cartílago solo se hace visible gracias a estas linternas traslúcidas que cuelgan en la neblina verde-dorada como arquitecturas de vidrio suspendidas entre el sol y el abismo.
