Confianza científica: Alto
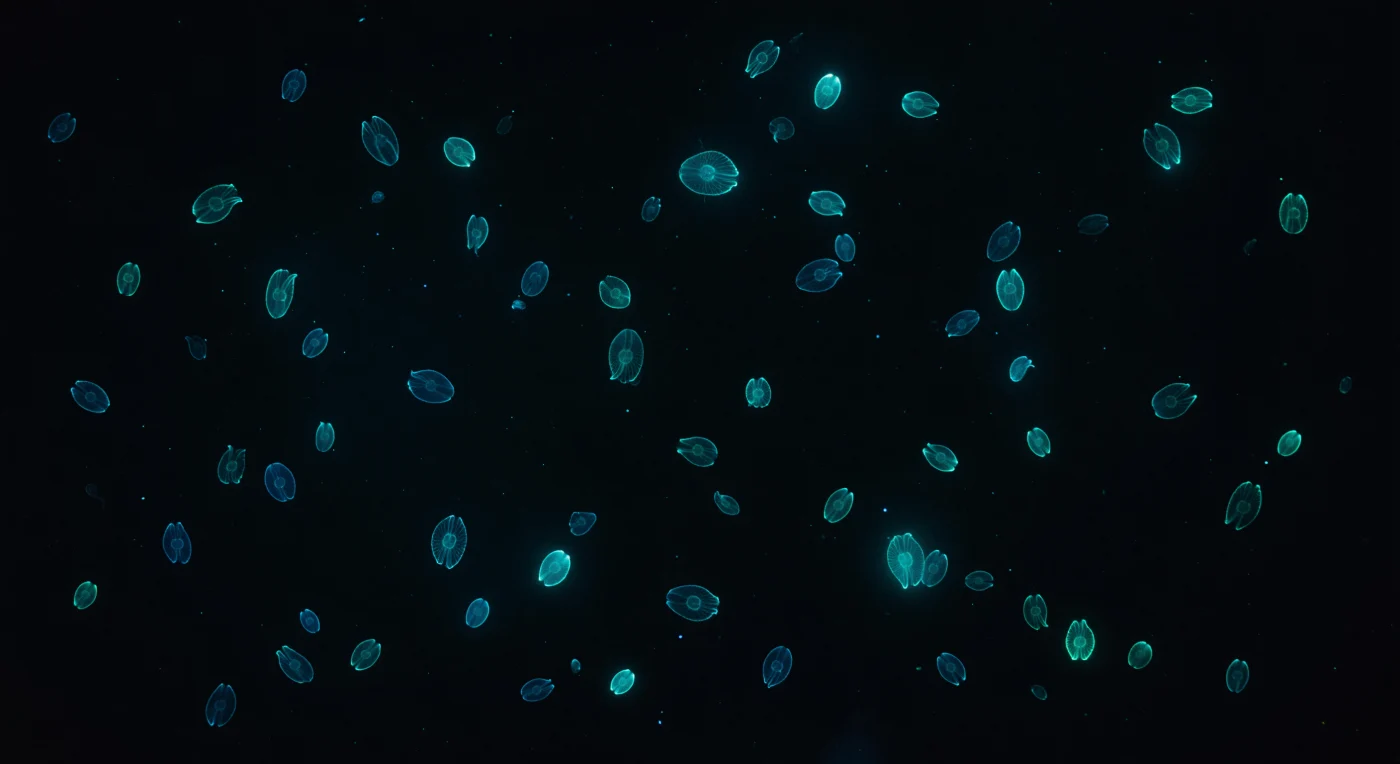
Suspendido en la oscuridad absoluta a dos metros bajo la superficie sin luna, el mundo se ha reducido a negro puro interrumpido por constelaciones vivas: decenas de ctenóforos del género *Mnemiopsis leidyi* derivan en todas direcciones, sus cuerpos ovalados casi invisibles salvo por ocho cintas luminosas de fuego azul-verdoso frío que trazan sus contornos contra el vacío a exactamente 490 nanómetros, esa tonalidad cercana al verde azulado que no existe en ningún paisaje terrestre. Cada fila de peines emite ondas de bioluminiscencia que viajan continuamente desde el polo aboral hacia el oral —un pulso lento e hipnótico como una respiración hecha de fotones—, mientras que la mesoglea, perfectamente equiparada en índice de refracción con el agua de mar, hace desaparecer el cuerpo mismo: solo las ocho líneas luminosas lo definen, una catedral de luz construida con tejido vivo cuya matriz extracelular es noventa y cinco por ciento agua. Los animales más distantes se reducen a halos difusos donde la nieve marina y los microorganismos en suspensión dispersan cada fotón antes de extinguirlo, confiriendo al agua negra una textura volumétrica y una profundidad que de otro modo no tendría. La sensación es la de flotar dentro de una nebulosa oscura cuyos astros están todos vivos, todos a la deriva en corrientes imperceptibles, todos portando su propia luz fría a través de un océano tan inmóvil y oscuro que cada fotón parece precioso y deliberado.

En la oscuridad absoluta a cuatrocientos metros de profundidad, el haz azul del ROV corta el agua como un bisturí de luz de 470 nanómetros y, en un instante, el vacío se puebla: los cuerpos lobulados de *Bathocyroe fosteri* emergen de la nada, masas de mesoglea pigmentada de color vino oscuro que absorben la luz ambiental hasta volverse agujeros negros gelatinosos, pero bajo la excitación azul sus lóbulos orales carmesí arden con una saturación que parece imposible a esta profundidad, formas de terciopelo plegado suspendidas en la columna de agua como linternas encendidas por brasas internas. A lo largo de sus ocho filas de peines, las placas ciliares compuestas fragmentan el haz en pulsos iridiscentes —violeta, verde azulado, ámbar— que recorren cada hilera en ondas metacrónicas, no bioluminiscencia sino física óptica pura: redes de difracción vivientes construidas de cilios que descomponen la luz en cintas espectrales fugaces. Todo el volumen iluminado está texturizado por nieve marina, agregados de mucus, colonias bacterianas y detritos fecales de uno a cinco milímetros que descienden a poco más de un metro por minuto hacia el sedimento remoto, los más cercanos nítidos y blancos en el haz, los más lejanos fantasmas grises que se disuelven en una oscuridad que no es un color sino la negación de todo color. La quietud total —sin turbulencia, a cuatro grados Celsius bajo una presión que aplasta el tiempo mismo— convierte la escena en un instante congelado: geometría carmesí contra vacío eléctrico azul, criaturas diseñadas para nunca ser vistas ofreciéndose brevemente a la luz prestada de una máquina intrusa.

Suspendidos sin peso en la columna de agua mediterránea, a dos metros bajo la superficie, nos hallamos frente a frente con una de las formas de vida más etéreas del planeta: un *Bolinopsis infundibulum* cuyo cuerpo de mesogleacasi perfectamente equiparado en índice de refracción al agua circundante —un gel viscoelástico de colágeno, glicoproteínas y agua, un noventa y cinco por ciento líquido— no existe tanto como materia visible sino como una distorsión óptica apenas perceptible, un fantasma de refracción que flota inmóvil en el espacio azul-verde. Lo que lo delata con certeza son sus ocho hileras de ctenos: cintas arqueadas de placas ciliares compuestas que propagan ondas metacrónicas antipléticas desde el polo oral hacia el aboral a quince o más ciclos por segundo, transformando cada placa en una rejilla de difracción móvil que descompone la luz en cascadas espectrales de carmesí a ámbar, de verde ácido a violeta eléctrico, como auroras boreales corriendo en cadencia escalonada sobre un espejo mojado. Los lóbulos orales del animal cuelgan abiertos en pliegues suaves, levemente rosados allí donde los canales gastrovasculares ramificados se vuelven visibles como capilares contra la luz cenital, mientras debajo, a cuatro metros, el suelo arenoso recibe encajes de cáusticas doradas —cuerdas sinuosas de luz concentrada que se deslizan sobre la sedimentación ripada y los restos de posidonia—, convirtiendo toda la columna de agua intermedia en una nave catedralicia de luz filtrada y azul profundo. Esta criatura sin cerebro, sin corazón, sin sistema circulatorio convencional, es sin embargo un motor biológico preciso: sus placas ciliares baten en fases independientes a cada lado del cuerpo, sus coloblastos esperan pegajosos en tentáculos que pueden extenderse veinte veces la longitud del cuerpo, y toda su existencia transcurre en este medio de salinidad y presión calibradas, deriva y transparencia, donde ser invisible es la estrategia evolutiva suprema.
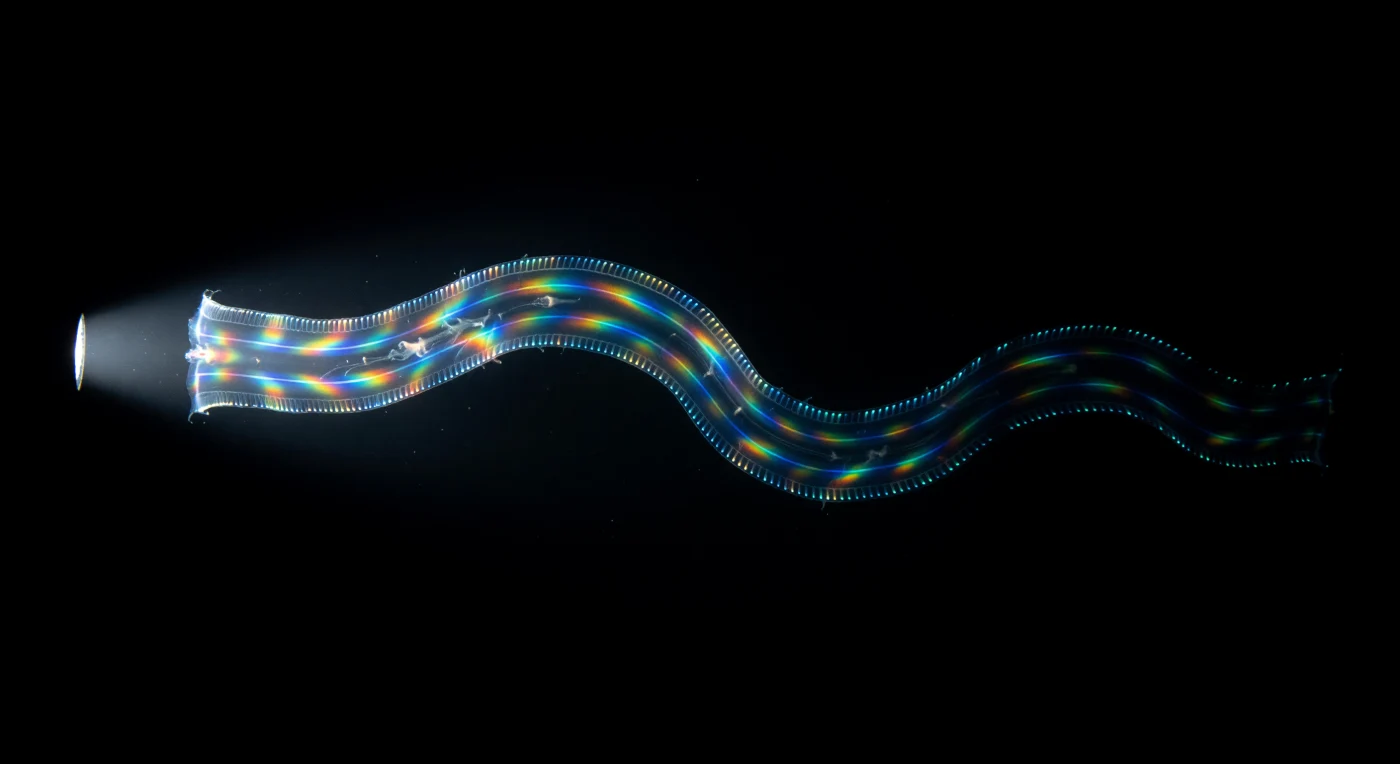
Frente a ti, materializándose desde la nada en la oscuridad absoluta a tres metros de profundidad, una cinta de tejido gelatinoso de sesenta centímetros ondula con gracia sinusoidal lenta, tan transparente que la percibes casi solo como una leve curvatura del vacío, una membrana que separa una oscuridad de otra. Sus cuatro filas de peines —estructuras compuestas de miles de cilios fusionados en placas que baten en ondas metacrónicas a entre quince y treinta y cinco hercios— corren a lo largo de toda su extensión como costuras luminosas, emitiendo una bioluminescencia azul verdosa fría de 490 nanómetros producida por fotocitos en la mesoglea subyacente, trazando cada flexión del cuerpo en luz de zorro contra el negro impenetrable del Atlántico tropical. Cuando el haz de tu linterna golpea la cinta de lado, los mismos peines que actuaban como discretas velas ahora explotan en difracción espectral plena: los espacios mecánicos entre miles de cilios en movimiento funcionan como redes de difracción vivientes, y rojo, naranja, oro, verde, cobalto y violeta se despliegan y enrollan en cascada estroboscópica a lo largo del ancho satinado del animal, colores saturados más allá de lo que el agua diurna permitiría. *Cestum veneris* —un depredador voraz construido en un noventa y siete por ciento de agua de mar, con una matriz mesogleica viscoelástica de colágeno cuyo índice de refracción casi iguala al del océano circundante— cruza el cono de luz y regresa a la oscuridad, y sus cuatro bordes bioluminescentes se curvan en lentas curvas serpentinas hasta desvanecerse por completo, dejando solo agua salada tibia y el recuerdo de algo que no tenía ningún derecho obvio a existir.

En las aguas heladas de un loch escocés en febrero, a quince metros de profundidad, docenas de Pleurobrachia pileus derivan en la columna de agua como canicas de cristal viviente, sus cuerpos esféricos de apenas dos centímetros construidos casi en su totalidad de mesoglea —una gelatina coloidal de colágeno y agua cuyo índice de refracción apenas difiere del mar que los rodea, volviéndolos casi invisibles salvo por el suave desplazamiento óptico que trazan contra la turbidez verdosa. Cada individuo porta ocho filas de peines ciliares que barren la esfera como los meridianos de un mundo diminuto, y la batida metacrónica de sus miles de cilios fusionados genera ondas de color estructural —rosa apagado, ámbar cálido, un hilo fugaz de violeta dorado— que recorren cada fila con una cadencia casi respiratoria, sin luz propia sino como un efecto de interferencia en el escaso resplandor que desciende filtrado por el agua teñida de turba. Detrás de cada esfera se extienden tentáculos de quince centímetros, filamentos de gossamér prácticamente invisibles cubiertos de coloblastos adhesivos que atrapan copépodos y larvas de zooplancton al menor contacto, retrayéndose luego hacia la boca en cuestión de segundos. Al fondo del encuadre, el margen oscuro de una fronda de kelp —pardo umbra, coriáceo, sólido— ofrece el único material denso en un mundo por lo demás compuesto de agua, gel y luz escasa, mientras la suspensión de fragmentos de diatomeas dispersa el tenue resplandor superficial en una niebla ambiental sin dirección que envuelve esta congregación invernal de predadores translúcidos.

En el corazón de la corriente del Golfo, suspendidos en una columna de agua que oscila entre el azul cobalto y el índigo del abismo, dos arquitecturas de gelatina llevan a cabo la transacción más silenciosa del océano abierto: un *Beroe cucumis* de color salmón-coral ha dilatado su macrostoma en un óvalo improbable y ha engullido más de la mitad de un *Mnemiopsis leidyi* que todavía sigue vivo dentro de él, mientras sus cuatro filas de paletas ciliadas posteriores continúan ejecutando sus programas metacrónicos en el agua libre, disparando cascadas de color estructural —violeta, ámbar, verde— que se desvanecen desde el extremo oral hacia afuera como una señal que aún no sabe que la fuente se ha perdido. El *Beroe*, cuya mesoglea no es verdaderamente tejido sino un gel viscoelástico de colágeno, glicoproteínas y agua con un índice de refracción casi idéntico al del mar, se ilumina desde dentro: sus canales meridionales ramificados trazan una delta fluvial de venación rosa-apricot en perfecta simetría bilateral, pulsando suavemente mientras la digestión comienza a disolver al depredador desde adentro. La luz de la tarde tardía, fragmentada por la superficie agitada en redes de cáusticos dorados, se curva alrededor de los cuerpos translúcidos como si ambos organismos fueran lentes vivientes, concentrando y dispersando destellos que convierten todo el episodio —noventa y cinco por ciento agua, cero huesos, cero sangre— en algo que parece más visión que depredación.

Suspendidos en la columna de agua negra de una estuarina cálida a las dos de la madrugada, percibimos tres cuerpos de *Mnemiopsis leidyi* que se yerguen a nuestro alrededor como bóvedas catedralicias de gelatina translúcida, sus paredes de mesoglea tan finamente ajustadas en índice de refracción al agua marina que las distinguimos menos como materia sólida y más como distorsiones orgánicas del espacio, mientras sus ocho hileras de ctenas pulsan en secuencia metacrónica y desencadenan cascadas de bioluminiscencia —cobalto profundo virando a verde azulado a 490 nm— que recorren cada placa ciliar como un hilo de fibra óptica viva. Desde las hendiduras gonopóricas que flanquean los canales meridionales emergen exhalaciones lechosas de esperma, plumas viscosas que derivan con la microcorriente y capturan la luz emitida por el animal vecino hasta volverse brevemente plateadas con bordes azul frío, creando donde se superponen una niebla reproductiva suspendida en media agua de una delicadeza comparable al cirro atmosférico. Entre esas nubes pelágicas ruedan libremente los óvulos —esferas refractivas de apenas 120 micrómetros que actúan como microlentes, concentrando la emisión bioluminiscente en un punto brillante interior rodeado de un halo difractivo violeta-anaranjado—, y docenas de ellos fulguran simultáneamente a distintas profundidades de campo, componiendo una constelación tridimensional de chispas frías que se disuelven en la oscuridad absoluta. Todo el escenario —paredes gelatinosas irradiando su marea luminosa, nubes de gametos a la deriva, la lluvia de huevos como lentes esféricas— constituye una aurora submarina enteramente biológica, sostenida únicamente por la energía química de los seres vivos en el silencio salino y tibio de la bahía.

Desde doce metros de profundidad en el Hardangerfjord, la mirada dirigida hacia arriba revela una nave catedralicia de cuerpos gelatinosos suspendidos en toda la columna de agua: cientos de *Bolinopsis infundibulum* que, con sus ocho a doce centímetros de mesoglea casi perfectamente transparente, se confunden con el propio mar y solo se delatan como leves distorsiones refractivas cuando los rayos oblicuos del sol nórdico de agosto los atraviesan. Cada individuo lleva ocho hileras de peines que recorren su cuerpo de polo a polo —bandas de cilios fusionados que baten en ondas metacrónicas a quince o treinta y cinco ciclos por segundo— y al difractar la luz producen una secuencia continua de rojo, naranja, verde y violeta que se desplaza placa a placa como fuego prismático lento; como ningún animal sincroniza su fase con sus vecinos, el conjunto oscila en una hoguera policromática asíncrona que transforma la columna de agua en un candelabro vivo. Varios de los ejemplares más cercanos muestran en sus canales gastrovasculares un rubor cálido de color albaricoche —la señal translúcida de copépodos recién ingeridos, visible como una yema dentro de un huevo sostenido a contraluz—, mientras el espejo de mercurio plateado de la superficie, surcado por redes de cáusticas luminosas, flota lejano sobre toda esta multitud silenciosa y gélida. Entre los cuerpos deriva nieve marina en suspensión y copépodos que huyen en arcos bruscos, y la luz se estrecha hacia abajo en hilos azul frío donde la población se adelgaza y los individuos más profundos son apenas óvalos pálidos con ocho franjas tenuemente luminosas sobre un fondo oscuro.

Te ciernes en la oscuridad absoluta del Cañón de Monterey, a mil metros de profundidad, mientras el cono azul del LED del ROV atraviesa el vacío y encuentra algo que parece imposible: *Lampocteis cruentiventer*, una ctenófora lobada del color de la sangre arterial, suspendida inmóvil en el negro perfecto como una llama carmesí sin fuente. Su mesoglea —ese gel viscoelástico de colágeno, glicoproteínas y agua que constituye más del noventa y cinco por ciento de su masa— absorbe la luz azul y la devuelve únicamente en su propio escarlata profundo, pues ninguna longitud de onda por debajo de los seiscientos nanómetros sobrevive al contacto con ese pigmento, haciendo que el animal parezca autoiluminado en un espectro que el ROV no puede suministrar. Los lóbulos orales cuelgan abiertos y relajados como los pétalos de terciopelo de un tulipán en plena floración, sus bordes superiores capturando el haz azul en un destello cian preciso que traza la curvatura exacta del margen antes de desvanecerse en la sombra color vino oscuro, mientras ocho hileras de ctenios —esas paletas de cilios fusionados que definen al filo Ctenophora— imprimen sutiles costillas topográficas sobre los flancos del cuerpo y proyectan microsombras bajo la luz rasante. La nieve marina deriva en todas direcciones a través del cono de iluminación, partículas blancas de tamaño variable moviéndose en vectores ligeramente distintos según la corriente ambiente, convirtiendo el agua negra en un espacio medible y otorgando al animal, de apenas unos pocos centímetros de diámetro, la escala justa de su soledad en el abismo.

Suspendido sobre el polo aboral de un *Mnemiopsis leidyi* vivo, el ojo percibe ante todo una cúpula hemisférica de vidrio biológico puro —unos doscientos micrómetros de diámetro— que emerge de una planicie de tejido transparente como una catedral construida a partir de una sola gota de agua, su pared tan ópticamente homogénea que solo se traiciona por una delgada caustica de luz azul-blanca que traza su ecuador donde la iluminación oceánica transmitida se dobla levemente a través del colágeno y las glicoproteínas. En el interior de esa cúpula flota, sin soporte visible, un agregado cremoso de gránulos de carbonato cálcico —el estatolito—, una masa de blanco mate y marfil cálido que descansa en equilibrio sobre cuatro abanicos de cilios balanceadores dispuestos en posiciones cardinales, cada uno de ellos convertido por su frecuencia de batido en un velo vibratorio translúcido que dispersa la luz transmitida en aureolas de luminiscencia perlada. Cuatro surcos ciliados irradian desde la base de la cúpula como las costuras de una rosa de los vientos, sus superficies cubiertas de cilios más finos cuya difracción estructural arranca destellos de verde y plata a los ángulos rasantes, y se prolongan hasta donde las primeras filas de peines comienzan sus trayectorias arqueadas, perceptibles apenas como cintas luminosas que se curvan fuera de campo con un tenue fulgor azul-verdoso de bioluminiscencia residual. Todo el conjunto está bañado por la iluminación difusa del océano profundo, que penetra desde todas las direcciones a través de la mesogolea translúcida —un gel viscoelástico casi idéntico en índice de refracción al agua de mar— de manera que no existen sombras duras, solo gradientes que van del azul-blanco en el vértice de la cúpula al cobalto profundo en los recesos de tejido más alejados, mientras que más allá del perímetro del animal el océano abierto se extiende como un vacío luminoso de azul absoluto que hace que esta arquitectura de precisión submilimétrica parezca simultáneamente íntima e infinitamente suspendida en el espacio.

En el límite exacto entre la superficie iluminada y la oscuridad que se abre hacia el abismo, una larva cydippida de apenas dos milímetros flota como una esfera de vidrio soplado, tan ópticamente transparente que se percibe más como una distorsión leve del agua que como un cuerpo sólido, su índice de refracción casi idéntico al del medio que la rodea. La luz del amanecer desciende en láminas doradas y azul pálido desde la microcapa superficial, doblándose a través del cuerpo gelatinoso y encendiendo sus ocho filas de peines rudimentarias en destellos prismáticos fugaces —rosa, violeta, blanco— que no son bioluminiscencia sino color estructural puro, la firma óptica de cilios en movimiento difractando fotones reales. A su alrededor, el mundo acuático es cualquier cosa menos vacío: una cadena de diatomeas Chaetoceros deriva como un tren de carga de vidrio silíceo dorado, cada frústulo facetado tan grande en esta escala que sus espinas hialinas proyectan agujas de luz dispersa sobre la larva entera, mientras un nauplio de copépodo de color cobre anaranjado rota sobre su eje como una nave mecánica impulsada por apéndices que baten el agua con ritmo deliberado. Dos tentáculos filiformes ya se extienden desde el cuerpo larvario hacia la nieve marina circundante —agregados de mucus, células fitoplánctonicas y detritos en ámbar, verde oliva y oro difuso— sus puntas adhesivas probando un océano que, para esta criatura de mesoglea casi pura, es simultáneamente hogar, alimento y cuerpo propio.

Suspendido en la columna de agua a media tarde, el observador flota sin peso en un azul que se transforma gradualmente de cerúleo cálido en lo alto a un verde frío y sombrío en lo profundo, y exactamente a la altura de los ojos, la termoclina se manifiesta como un espejo horizontal tembloroso: una lente continua de distorsión refractiva que ondula a través de todo el campo visual, revelando el límite entre dos masas de agua de diferente densidad y temperatura como una película de calor suspendida en el océano abierto. Tres *Mnemiopsis leidyi* —cada uno un óvalo de cinco a ocho centímetros de mesoglea casi perfectamente transparente, un gel viscoelástico de índice de refracción casi idéntico al del agua marina— se mantienen inmóviles precisamente en esa frontera, con sus lóbulos orales extendidos hacia la zona más fría y rica en presas, donde decenas de copépodos de color naranja-rojo forman una constelación de brasas suspendidas en la penumbra verdosa. A lo largo de los flancos de cada ctenóforo, las ocho filas de peines —formadas por miles de cilios fusionados en placas que baten en ondas metacrónicas a entre quince y treinta y cinco hercios— despliegan en secuencia lenta un arcoíris de color estructural: rubí deslizándose hacia ámbar, ámbar hacia verde ácido, verde hacia índigo, una difracción de la luz que recorre el cuerpo de la boca al polo aboral en menos de un segundo, el único color vívido en un universo construido de gradientes azules y sombra verde. La línea refractiva de la termoclina atraviesa cada cuerpo transparente como una regla trazada sobre vidrio, dividiendo su mitad superior bañada en oro filtrado de su mitad inferior sumergida en el crepúsculo frío, mientras los movimientos ciliares de sus lóbulos generan remolinos invisibles que espiralizan los copos de nieve marina a su alrededor.

Te encuentras suspendido en el interior de una larva cydippid de *Mnemiopsis leidyi*, un cosmos sellado de dos milímetros cuya oscuridad absoluta no es la del océano sino la de un sistema óptico hermético, perforado por puntos azules de DAPI que flotan en el volumen de la mesoglea como núcleos celulares a distintas profundidades, cada uno una esfera cerúlea de bordes nítidos, sin patrón, sin gravedad aparente. Desde esa nada emerge una red de filamentos magenta —el sistema nervioso difuso marcado con FMRFamida— ramificándose en todas direcciones sin jerarquía ni centro discernible, cada fibra emitiendo un halo rosado suave que sugiere luminiscencia propia antes que reflexión, un entramado que a cualquier plano focal parece escaso pero que, al profundizar la mirada, revela una densidad extraordinaria. Ocho arcos de verde ácido barren el espacio superior como los nervios de una bóveda catedralicia: las bandas serotoninérgicas de las hileras de peines, estructuras que imponen una gramática espacial sobre la anarquía de los filamentos, y donde el magenta las cruza brevemente nace un ámbar cálido, el único acorde térmico en una paleta dominantemente fría. En el cénit, el órgano apical arde como un halo circular perfecto —la co-localización de ambos fluoróforos en el estatocisto produce un anillo de blancura casi pura, magenta en el borde exterior y verde hacia el interior— irradiando una penumbra violácea que ilumina las últimas ramificaciones del plexo nervioso como si fuera el ojo quieto de una tormenta de luz. Todo el conjunto posee la cualidad de una astrofotografía de nebulosa, donde la profundidad se comunica únicamente por el gradiente de nitidez entre filamentos a distintos planos virtuales, y el animal entero —este depredador gelatinoso cuya masa es un noventa y siete por ciento agua— se experimenta desde adentro con la escala sentida de una galaxia a la deriva.

Suspendido en la columna de agua azul del kreisel, el *Mnemiopsis leidyi* llena el campo visual como una nave de vidrio vivo: su mesoglea, casi perfectamente transparente, no es vacío sino un gel viscoelástico de colágeno y glucoproteínas cuyo índice de refracción iguala al del agua de mar, revelando su arquitectura interna a través de la refracción pura antes que por la opacidad. Las ocho hileras de peines se encienden desde abajo con la luz LED azul-blanca, y cada placa —formada por miles de cilios fusionados en un remo compuesto que bate en onda metacrónica antiplectica— irradia una secuencia de fuego espectral: rojo-ámbar en las placas superiores virando a verde eléctrico hacia la mitad y enfriándose en violeta-azul hacia el polo aboral, color generado no por pigmento sino por la geometría de difracción de los cilios. Flanqueando el estomodeo, dos franjas gonádales de coral pálido y ámbar cálido contrastan con el frío azul circundante, su textura granular íntima y orgánica frente a la translucidez del tejido que las rodea. Los lóbulos orales —labios de tejido gelatinoso texturado como vidrio esmerilado húmedo— se contraen y expanden en ritmo peristáltico lento, sus bordes orlados de una membrana gaseosa que el rayo ascendente hala en lavanda difusa. La pared curva del tanque se convierte a esta escala en un horizonte oscuro y perfectamente circular, y la punta de pipeta difuminada al borde del encuadre —mundana en el laboratorio, monolítica aquí— devuelve al observador a la realidad de un universo doméstico contenido en seis centímetros de vida suspendida.

En el corazón del océano mesopelágico, a media distancia entre la luz superficial y la oscuridad absoluta, el observador se encuentra suspendido junto a una estructura que apenas merece llamarse sólida: un tentillum de *Pleurobrachia*, rama lateral de un tentáculo no más grueso que un hilo de vidrio estirado, cuya translucidez azul-hielo revela tenues estrías citoesqueléticas internas mientras la hebra entera vibra con la tensión arrestada de una retracción a medio completar. A lo largo de su longitud se apiñan los coloblastos, orgánulos adhesivos exclusivos de los ctenóforos, cada uno una cúpula hemisférica de diez a cincuenta micrómetros de diámetro, empaquetados de gránulos refractivos que devuelven destellos blanco-dorados al ambiente azul-negro profundo como una constelación de perlas pegajosas —algunos todavía intactos y esféricos, otros aplastados contra la quitina del crustáceo en discos de contacto irreversible donde los filamentos en espiral, cual resortes semidesplegados, anclan la cabeza adhesiva al núcleo del tentillum mediante bioquímica que no admite retracción. Irrumpiendo desde el ángulo superior del encuadre, una antena de *Calanus* —viga articulada de quitina ferruginosa y siena cálida, esculpida en crestas transversales y sétulas doradas— ha rozado el tentillum y activado la descarga de varios coloblastos, cuyas cabezas granulares se han extendido en halos adhesivos apenas visibles contra la superficie del exoesqueleto, mientras una pata natatoria translúcida aún forcejea inútilmente contra la hebra, imprimiendo en su curvatura infinitesimal la única evidencia visible de la fuerza de una presa que ya ha perdido. Esta escena de depredación congelada revela la estrategia fundamental de los ctenóforos: sin aguijón ni mandíbula, solo la paciencia gelatinosa de un animal que es noventa y siete por ciento agua atrapando a uno de los crustáceos más veloces del plancton oceánico mediante adhesión molecular, en el silencio azul de un mundo donde la violencia no hace ruido.
