Affidabilità scientifica: Alto
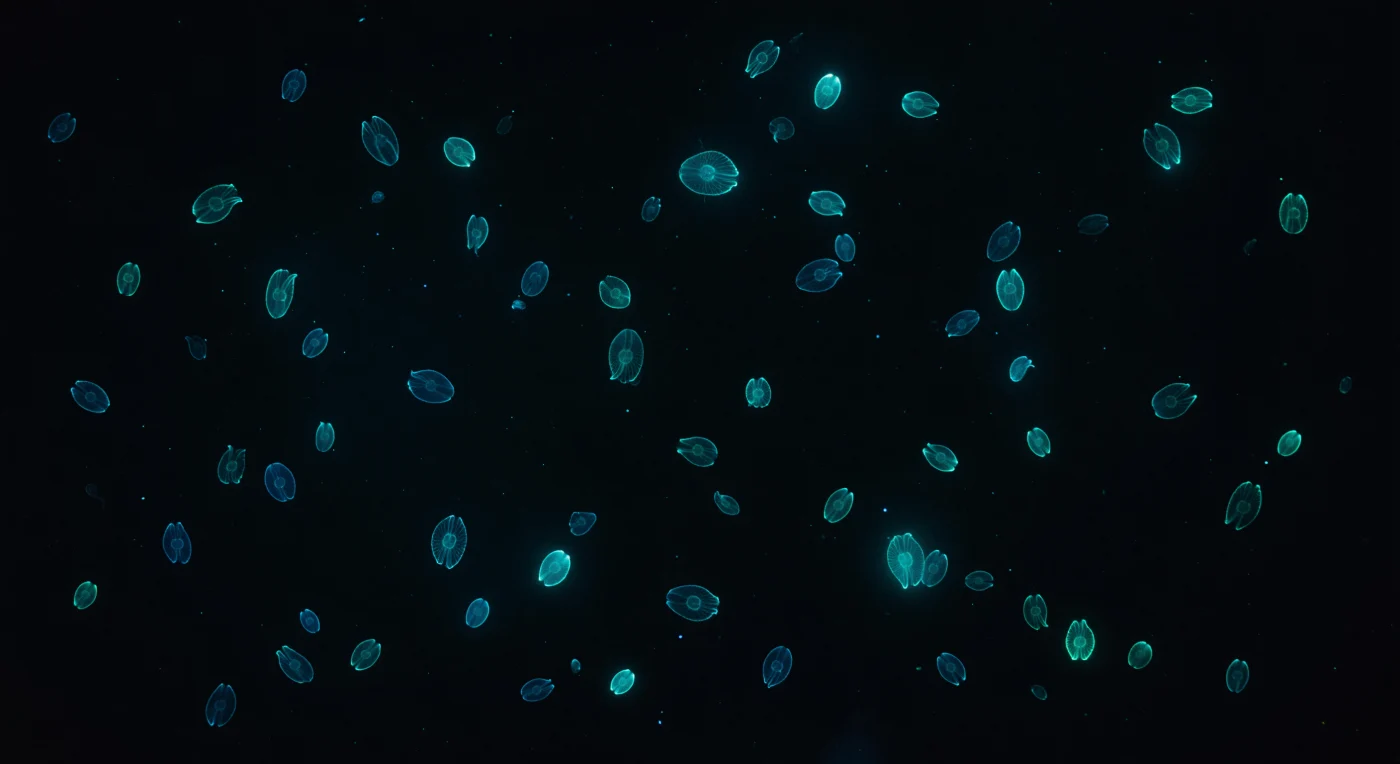
Sospesi nell'oscurità assoluta a due metri sotto una superficie senza luna, l'oceano si è ridotto al nero puro interrotto da costellazioni viventi: decine di *Mnemiopsis leidyi* derivano in ogni direzione, i loro corpi ovali quasi invisibili tranne per otto nastri luminosi di fuoco azzurro-verde freddo che ne tracciano i contorni nel vuoto. Ogni fila di pettini emette esattamente a 490 nanometri — quella specifica tonalità teal che non esiste da nessuna parte sulla terraferma — e onde di attivazione bioluminescente viaggiano continuamente dal polo aborale verso quello orale, un impulso lento e ipnotico come un respiro fatto di fotoni, ogni onda impiegando un secondo intero ad attraversare la lunghezza del corpo prima che la fila si spenga e si riaccenda dall'apice. La mesoglea, il gel extracellulare che costituisce il 97% della massa dell'animale, è così perfettamente accordata all'indice di rifrazione dell'acqua marina da risultare completamente assente alla vista — solo le otto linee luminescenti ne definiscono l'esistenza, una cattedrale di luce costruita da tessuto vivente e colloblasti, strutture adesive specializzate per la caccia, invisibili nel buio. Più lontano, quaranta o cinquanta organismi in aggregazione libera riducono le loro file bioluminescenti a corone soffuse, aloni ciano che sanguinano nell'acqua dove la neve marina e i microrganismi in sospensione catturano e diffondono ogni fotone prima di spegnerlo, conferendo alla colonna d'acqua buia una qualità di profondità e texture volumetrica altrimenti impossibile.

Nell'oscurità assoluta a quattrocento metri di profondità, dove la pressione schiaccia il silenzio in qualcosa di fisicamente percepibile, il fascio LED blu del ROV taglia l'acqua come una rivelazione improvvisa e raduna il mondo dal nulla: i *Bathocyroe fosteri* si materializzano uno a uno, lanterne di velluto bordeaux e cremisi sospese nel vuoto, i loro lobi orali accesi di un rosso saturo che non dovrebbe esistere in questo luogo, la mesoglea pigmentata così densa di colore da assorbire il blu circostante e trasformarsi in volumi quasi opachi, voragini scure che si stagliano contro un nero ontologico. Lungo le otto file di pettini, le placche ciliari diffrattono la luce in sequenze metronomiche di violetto, turchese e ambra fugace — non bioluminescenza, ma fisica pura, reticoli di diffrazione biologici che trasmettono segnali spettrali lungo il corpo come impulsi nervosi rallentati fino alla contemplazione. Attorno agli animali, la neve marina riempie l'intero cono illuminato di texture fine e paziente — aggregati di muco, materiale fecale e colonie batteriche da uno a cinque millimetri, ciascuno che precipita verso il fondale lontano a circa un metro al minuto, i granuli più vicini bianchi e nitidi, quelli più lontani fantasmi grigi che svaniscono nella transizione tra luce e assenza di luce. Questi organismi, composti per il novantasette per cento di acqua e quasi perfettamente isorifrattivi con il mare che li contiene, vivono nell'invisibilità come condizione evolutiva primaria, e solo il dono temporaneo di un fascio artificiale permette di vedere cosa la pressione, il freddo a quattro gradi Celsius e l'oscurità eterna custodiscono in questo intervallo dimenticato della colonna d'acqua.

Sospesi nella colonna d'acqua mediterranea, a due metri sotto la superficie, ci troviamo di fronte a una creatura che esiste appena come materia: la *Bolinopsis infundibulum*, il cui corpo di mesogleaè così perfettamente accordato all'indice di rifrazione dell'acqua circostante da dissolversi in essa, tradito solo da un lieve tremolio ottico ai suoi margini. Ciò che brucia con realtà inconfutabile sono le otto file di pettini — larghi nastri di ciglia composte che percorrono il corpo in onde metacrone antiplettiche, dal polo orale a quello aborale, dipingendo cascate di carminio, ambra, verde acido e violetto elettrico in una sequenza spettrale continua, come un reticolo di diffrazione biologico che gira a trenta battiti al secondo per placca. I lobi orali dell'animale si aprono in morbide pieghe, appena velate dal rosa pallido dei canali gastrovascolari che ramificano attraverso la mesogleae diventano visibili solo dove la luce di superficie li attraversa come capillari in controluce. Quattro metri più in basso, la luce solare si frantuma in una raggiera di caustics dorate che scorrono sul fondale sabbioso, trasformandolo in un piano impressionistico di ambra e ombra cobalt, mentre la colonna d'acqua intermedia si gonfia di una traslucenza blu-verde satura di fotoni, come la navata di una cattedrale liquida. In questo spazio al tempo stesso intimo e oceanico, la creatura pulsa nella sua iridescenza asimmetrica — un fantasma di vetro e arcobaleno che non emette suono, non produce turbolenza, e che è composto per il novantasette per cento dell'acqua stessa che lo circonda.
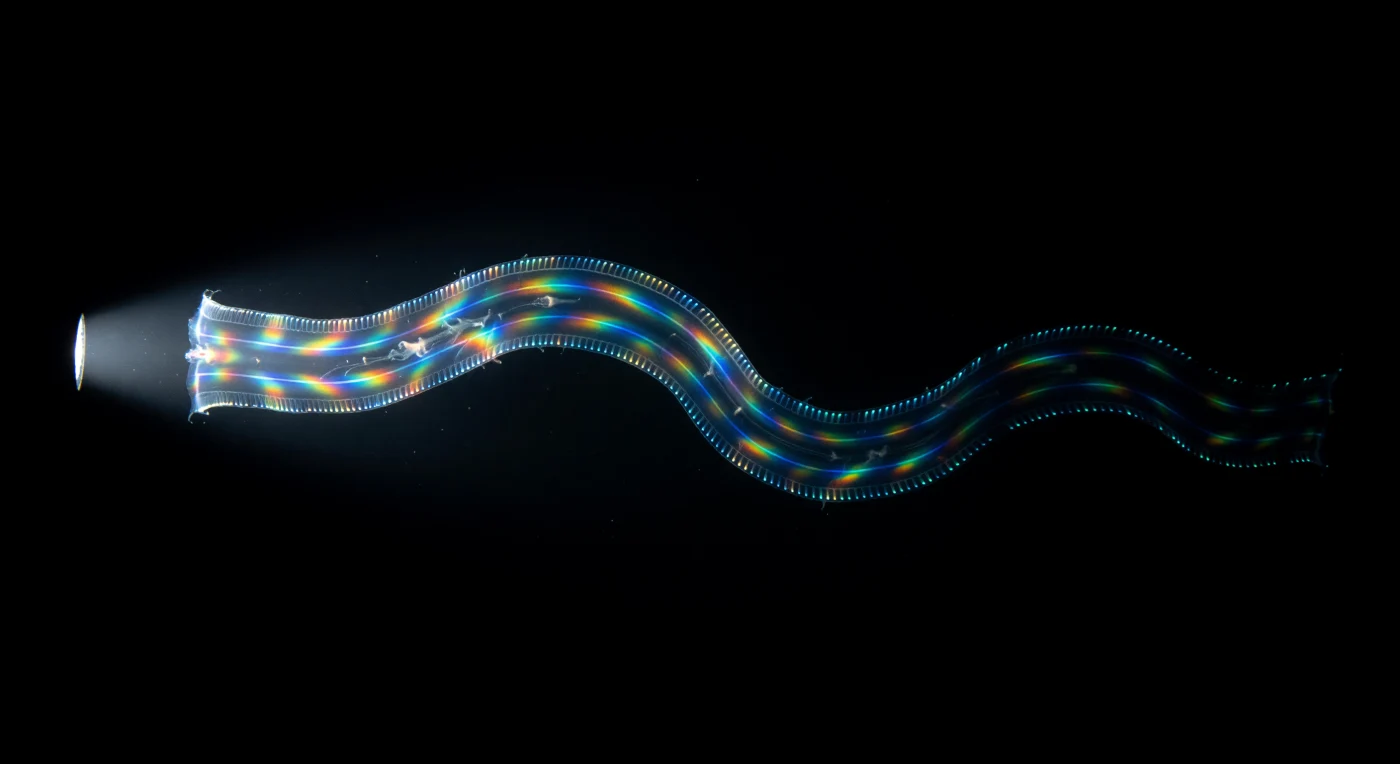
Nell'oscurità assoluta di tre metri sotto la superficie dell'Atlantico tropicale, un nastro di tessuto gelatinoso trasparente emerge dal nulla con la lentezza irreale di un'allucinazione: è *Cestum veneris*, la cintura di Venere, sessanta centimetri di mesoglea così perfettamente accordata all'indice di rifrazione dell'acqua da rendersi quasi invisibile, tradita soltanto da quattro linee di bioluminescenza blu-verde che corrono lungo i suoi bordi — i filari di pettini ciliari che sparano luce a 490 nanometri in onde sequenziali, fotociti che si accendono uno dopo l'altro come un circuito in cascata tracciando l'intera architettura dell'animale nell'oscurità con l'autorità silenziosa di un tubo al neon. L'organismo ondeggia in un'unica sinusoide lenta e continua, l'intero corpo piatto che respira come seta abbandonata a mezz'aria, spinto da battiti ciliari a 15-35 Hz in onde metacrone che percorrono ogni filare in decine di millisecondi. Poi il fascio della torcia lo colpisce di traverso, e i pettini — reticoli di diffrazione in movimento — esplodono in un'iridescenza strutturale che scorre da rosso ad arancio a verde a cobalto a violetto lungo tutta la larghezza del nastro in un'unica cascata cromatica pulsante, colori saturi e impossibili che avanzano in onde mentre la materia gelatinosa del corpo cattura la luce bianca e rivela per un istante la rete dei canali interni, l'architettura fantasma di un essere che è per il 97% acqua di mare e che, quando rientra nel buio, lascia dietro di sé soltanto quelle quattro linee fredde e sinuose che si allontanano fino a scomparire nel nero dell'oceano come se non fossero mai esistite.

Ci si trova sospesi a quindici metri di profondità nelle acque scure e torbide di un sea loch scozzese in febbraio, immersi in una luce verde giada che filtra dall'alto senza mai formare ombre, diffondendosi invece in una nebbia ambientale densa di frammenti diatomacei e particolato in sospensione. Attorno a noi, una dozzina di Pleurobrachia pileus derivano nella colonna d'acqua come sfere di vetro soffiato del diametro di due centimetri, i loro corpi di mesogle quasi indistinguibili dall'acqua stessa se non per una sottile distorsione rifrangente lungo l'equatore e per il bagliore verdino-salvia dei contenuti intestinali visibili al loro interno come un nucleo luminescente sospeso nel nulla. Sull'individuo più vicino, otto file di piastre ciliari percorrono la superficie sferica come i meridiani di un piccolo mondo, ciascuna animata da un'iridescenza metaronale che scorre in sequenza dal rosa antico all'ambra pallido e al viola dorato, colori soffusi e organici come riflessi di una bolla di sapone sotto cielo coperto, ritmati con la cadenza quasi respiratoria del battito ciliare che propelle lentamente l'animale attraverso l'acqua gelida. Dietro ogni sfera si prolungano tentacoli gossamer di quindici centimetri, invisibili se non come sottilissimi fili d'argento che svaniscono nella torbidità, i loro colloblasti pronti a catturare la preda per contatto adesivo; sul margine inferiore del campo visivo, il bordo cuoioso e brunissimo di una fronda di kelp offre l'unica presenza materiale solida in un mondo altrimenti composto di acqua, gel e luce morente.

Nelle acque tiepide della Corrente del Golfo, a pochi metri sotto una superficie che spezza il sole pomeridiano in reti di caustica dorata, un *Beroe cucumis* ha dilatato il suo macrostoma fino a inglobare più della metà di un *Mnemiopsis leidyi* ancora vivo: una predazione silenziosa che procede per lenta interpenetrazione di mesoglee, due architetture gelatinose — ciascuna composta per oltre il novantacinque per cento di acqua — che si fondono in un'unica forma translucida color albicocca e rosa corallo. Attraverso il corpo del predatore, i canali meridionali ramificati brillano come un delta fluviale visto dall'alto, tributari rosa-carnicino che si biforcano in simmetria bilaterale e pulsano debolmente mentre inizia la digestione, la loro luce filtrata da una mesoglea così trasparente da sembrare aria tinta piuttosto che tessuto. La metà posteriore della preda sporge ancora nell'acqua aperta, e le sue quattro file di pettini esposti continuano il loro programma metacronale con indifferenza meccanica: ogni placca ciliare — centinaia di migliaia di ciglia fuse in palette di vetro larghe poche centinaia di micrometri — converte la luce radente in cascate di colore strutturale che scorrono dal violetto all'ambra al verde in frazioni di secondo, un arcobaleno involontario che si ripete e si affievolisce man mano che il ritmo si perde dall'estremità orale verso l'esterno. Le caustics della superficie scivolano in interferenze dorate su entrambi i corpi come se i loro profili curvi fossero lenti viventi, mentre la neve marina deriva nel blu cobalto sullo sfondo, offrendo all'intera scena la sua unica misura di profondità e silenzio.

Ci troviamo sospesi nella colonna d'acqua di un estuario estivo, immersi in un'oscurità quasi assoluta che viene spezzata soltanto dai corpi di tre *Mnemiopsis leidyi*, le cui pareti di mesolea trasparente si ergono come navate di gel vivo, talmente accordate all'indice di rifrazione dell'acqua da apparire non come forme solide ma come distorsioni organiche dello spazio. Lungo i loro fianchi, otto file di piastre ciliari pulsano in sequenza metacrona, e dove la bioluminescenza accende ciascun ctene in successione, un'onda di luce blu-cobalto che scivola verso il turchese si propaga lentamente lungo ogni filare come una fiamma che percorre una fibra ottica vivente, emettendo a 490 nm una radiosità fredda e diffusa che illumina i canali gastrovascolari come pizzo scuro nell'interno incandescente dell'animale. Dalle fessure dei gonopori lungo i canali meridionali, lente esalazioni di materiale riproduttivo si dispiegano nell'acqua come nubi perlacee, i pennacchi di sperma viscosi e debolmente luminescenti mentre catturano la luce dei vicini, formando una nebbia pelagica di urgenza riproduttiva che si addensa nelle zone di sovrapposizione in un alone argenteo orlato di blu freddo. Liberi in questo scenario tumultano le uova, sfere rifrangenti di circa 120 micrometri che si comportano come micro-lenti, concentrando la luce bioluminescente circostante in una caustica interna brillante — una stella compressa in ciascun vitello trasparente, circondata da un alone di diffrazione che separa la luce in un breve arcobaleno violetto-arancio — finché decine di queste sfere incandescenti a profondità differenti compongono nell'oscurità una costellazione tridimensionale di scintille fredde, interamente biologica, interamente silenziosa, sospesa nelle acque salmastre e calde della Chesapeake nel cuore della notte.

Guardando verso l'alto dai dodici metri di profondità dell'Hardangerfjord, il visitatore si trova immerso in una cattedrale liquida abitata da centinaia di *Bolinopsis infundibulum*: corpi ovali di mesoglea quasi perfettamente trasparente — un gel viscoelastico composto per il novantacinque percento di acqua, il cui indice di rifrazione è così prossimo a quello dell'acqua di mare da renderli leggibili solo come lievi distorsioni ottiche — sospesi in ogni strato della colonna d'acqua fino allo specchio argentato della superficie, che galleggia lontano come un soffitto di mercurio increspato da reti di luce caustica. Ciascun animale porta otto file di piastre ciliari — le cteni, strutture composte da centinaia di migliaia di ciglia fuse in palette battenti a frequenze di quindici-trentacinque hertz in onde metacronali sfasate — che diffrangono la luce nordica estiva in strisce prismatiche continue, ciclando dal rosso all'arancio al verde al violetto e di ritorno, mentre la fase ciliari di ogni individuo è sfasata rispetto ai vicini, così che l'intera massa pulsa di un fuoco policromatico lento e asincrono, un lampadario vivente di arcobaleni rotanti. Alcuni corpi mostrano una sfumatura ambra-arancio diffusa lungo i canali gastrovascolari — la traccia traslucida di copepodi recentemente ingeriti, visibile come un tuorlo d'uovo tenuto controluce attraverso la mesoglea incolore — mentre tra un animale e l'altro l'acqua conserva la propria microtexture di neve marina in lenta caduta, copepodi in fuga e sottili veli di muco che trascinano dagli lobi orali. La percezione della scala si impone non attraverso la dimensione del singolo organismo, di appena otto-dodici centimetri, ma attraverso la densità collettiva di centinaia di corpi che trasformano un fiordo norvegese in qualcosa di simile a una navata di vetro colorato, completamente silenziosa, gelida, continuamente luminosa.

Nella totale oscurità a mille metri di profondità nel Canyon di Monterey, il fascio di LED blu del ROV taglia il vuoto e colpisce qualcosa di inverosimile: *Lampocteis cruentiventer*, una ctenofora lobata la cui mesoglea è di un rosso arterioso così saturo da sembrare quasi chirurgico contro il nero assoluto che la circonda. Il corpo gelatinoso — viscoelastico come un gel collagene, composto per il novantasei percento di acqua fredda salata — trattiene la luce blu all'interno dei propri strati traslucidi e restituisce solo il suo scarlatto profondo, incapace di riflettere alcuna lunghezza d'onda inferiore ai seicento nanometri, così che l'animale sembra irradiare da solo in uno spettro che il ROV non può fornire. I lobi orali aperti pendono in avanti come petali di velluto di un tulipano in piena fioritura, i loro bordi superiori percorsi da riflessi ciano nitidi e precisi che ne tradiscono la vera fermezza gelatinosa, mentre otto file di pettini — file di piastre ciliari capaci di battere in onde metacronate a trenta hertz — tracciano costolature appena più scure lungo i fianchi del corpo, troppo ravvicinate per essere risolte individualmente ma abbastanza presenti da catturare la luce radente in micro-ombre. La neve marina deriva continuamente attraverso il cono luminoso in una nevicata particolata a traiettorie divergenti, dando profondità misurabile all'acqua altrimenti invisibile e ricordando all'osservatore che questo corpo rosso e silenzioso è sospeso nel mezzo di un oceano che si estende per chilometri in ogni direzione.

Sospesi appena sopra il polo aborale di un *Mnemiopsis leidyi* vivente, lo sguardo cade su una cupola emisferica di biologica trasparenza assoluta — circa duecento micron di diametro — che si erge da una pianura di tessuto gelatinoso con la geometria silenziosa di una cattedrale costruita da una singola goccia d'acqua. All'interno della cupola, l'organo apicale custodisce il suo statociste: un aggregato compatto di granuli di carbonato di calcio, bianco avorio opaco, sospeso nell'invisibile al di sopra di quattro ventagli di ciglia bilanciatrici che battono a frequenze irrapresentabili, risolte solo come aloni perlacei e tremolanti di luce diffusa. Questi ventilatori vibratili occupano le posizioni cardinali attorno allo statociste, e da essi si dipartono quattro solchi cigliati che si irradiano verso l'esterno come i meridiani di una bussola, la loro superficie coperta di ciglia più fini il cui movimento collettivo produce un fugace iridescente lavaggio di verdi e argenti — diffrazione strutturale della luce oceanica che sfiora il tessuto a incidenza radente. La mesoglea sottostante, vasta pianura di silicone biologico trasparente percorsa da fibre di collagene che si tradiscono solo come un fremito rifrattivo, ospita l'intera architettura su uno sfondo di blu oceanico profondo, visibile attraverso lo spessore del corpo come un vuoto luminoso e infinito che trasforma questa struttura minuta in qualcosa di cosmico.

Ci troviamo sospesi alla superficie dell'oceano all'alba, rimpiccioliti alla scala di un granello di sabbia, immersi in un mondo liquido dove la gravità si percepisce appena e ogni direzione si dissolve in una foschia traslucida. Davanti a noi fluttua la larva cydippid come una biglia di vetro soffiato — due millimetri di gelatina quasi invisibile, la sua sfera così perfettamente accordata otticamente all'acqua di mare da esistere più come una lieve distorsione-lente che come un oggetto solido, mentre cattura la pallida luce dorata dell'alba che filtra dall'alto in raggi acquosi che si piegano e si diffondono attraverso il microstrato superficiale. Otto rudimentali file di pettini tracciano lievi linee meridiane lungo il corpo della larva, ciascuna un nastro evanescente che emette il più tenue scintillio prismatico — una fugace scia dal rosa al violetto mentre i fotoni dell'alba si diffrangono sui proto-cteni in battito, pura colorazione strutturale prodotta dalle ciglia in movimento piuttosto che bioluminescenza in questa luce d'aurora. Due tentacoli sottili come capelli si estendono nell'acqua circostante come fili di seta lasciati cadere al rallentatore, mentre attorno a loro una catena di diatomee *Chaetoceros* deriva come un treno merci di lanterne silicee dorate, ciascuna cellula grande come un melone alla nostra scala ridotta, e un nauplio di copepode color rame-arancio ruota su se stesso come un'astronave aliena nella sospensione densa di neve marina, particelle ambrate e frammenti verdastri che popolano questo crepuscolo acquatico illuminato dall'alto in morbidi raggi causali.

Sospeso a quaranta metri di profondità, l'occhio incontra qualcosa di impossibile: una superficie orizzontale che non è superficie, una distorsione rifrattiva continua che attraversa l'intera colonna d'acqua come un vetro tremante — la termoclina, confine tra due masse d'acqua a densità e temperatura diverse, resa visibile dalla variazione dell'indice di rifrazione che incurva e ondula ogni forma al di là di essa. Tre *Mnemiopsis leidyi* stazionano con precisione assoluta su questa soglia invisibile, i loro corpi ovoidali di cinque-otto centimetri quasi perfettamente trasparenti — mesogleiche gelatinose al novantacinque percento d'acqua, con un indice di rifrazione allineato a quello del mare circostante — tanto che la riga lucente della termoclina li attraversa e riemerge dall'altro lato appena distorta, come se passasse attraverso lenti viventi. Le otto file di pettini cileari lungo ciascun fianco battono in onde metacrone che si propagano dalla bocca al polo aborale, e la luce pomeridiana filtrata — già smorzata e azzurrata dai quaranta metri di colonna soprastante — si diffrange su ciascuna placca ctenale in archi cromatici che scorrono dal rosso rubino all'ambra, al verde acido, all'indaco, al violetto, un arcobaleno strutturale che non emette luce propria ma la scompone e la restituisce in meno di un secondo per ogni ciclo. I lobi orali, ampi e opalescenti, pendono aperti nella zona più fredda e verde sotto la termoclina, dove decine di copepodi arancio-rossi — concentrati alla soglia di densità come flottiglie di semi in un campo gravitazionale sottile — pulsano con traiettorie scosse nell'oscurità verdognola del basso, mentre le correnti ciliari degli animali creano vortici impercettibili nello strato di confine, portando quella costellazione di prede vive verso l'interno in un atto di predazione silenzioso quanto il peso dell'acqua stessa.

Sospeso nell'oscurità assoluta di un sistema ottico sigillato, il visitatore si trova immerso nell'interno di una larva cydippide di *Mnemiopsis*, dove una rete non gerarchica di filamenti magenta — le fibre nervose marcate con FMRFamide — si irradia in ogni direzione senza centro discernibile, ogni filo pulsante come corallo incandescente che proietta aloni rosa diffusi nel vuoto circostante. Otto archi di verde acido si curvano in alto come le costole di una volta cattedrale, le bande dei ranghi di pettini positive alla serotonina, strutture luminose che impongono una grammatica spaziale all'anarchia filamentosa sottostante, convergendo verso l'organo apicale aborale che arde al zenith come un'aureola circolare di magenta e verde sovrapposti — il sistema statolito che coordina l'orientamento gravitazionale dell'animale. Sparsi per tutto il volume della mesolea, punti ceruleo-freddi di DAPI segnano i nuclei cellulari come una costellazione sommersa, ciascuno a profondità diversa, comunicando la tridimensionalità di questo spazio attraverso la gradazione dal nitido al dissolto. Quello che la fisica dell'ottica a foglio di luce rivela è un sistema nervoso privo di cervello — distribuito, ridondante, evolutivamente antichissimo — il cui funzionamento emerge non da una gerarchia centrale ma dalla geometria stessa della rete, un modello di complessità senza comando che i ctenofori hanno conservato per oltre cinquecento milioni di anni.

Sospeso nel blu freddo del kreisel, un singolo esemplare di *Mnemiopsis leidyi* occupa l'intero campo visivo come un'architettura di vetro vivente: il corpo, largo sei centimetri, è quasi interamente trasparente, con la mesoglea — un gel viscoelastico composto per oltre il novantacinque percento d'acqua — che rifrange la luce dei LED sottostanti in caustics morbide e in continua deriva. Le otto file di pettini, ciascuna composta da decine di placche ciliari larghe fino a mezzo millimetro, bruciano di iridescenza strutturale mentre l'onda metacrona le percorre in sequenza: rosso-arancio al polo orale, verde smeraldo a metà corsa, violetto-azzurro verso il polo aborale, colori generati non da pigmenti ma dalla diffrazione geometrica di migliaia di assonemi fusi insieme. Due strie gonadali color albicocca-corallo fiancheggiano lo stomodeo, uniche note calde in un mondo dominato dal freddo blu-bianco dell'illuminazione artificiale, mentre i lobi orali — labbra di tessuto gelatinoso finemente traslucido — si contraggono e si distendono in un ritmo peristaltico lento che ricorda una respirazione. Sullo sfondo, la parete curva nera del serbatoio si dissolve in un orizzonte circolare che dà la sensazione di fluttuare all'interno di un universo privato, e un puntale di pipetta fuori fuoco emerge ai margini dell'inquadratura come un monolite ambrato, misura silenziosa dell'intimità di questa scala.

Sospesi nell'oscurità mesopelagica, ci troviamo a pochi decimi di millimetro da un tentillum di *Pleurobrachia* — un filo di vetro vivente che occupa quasi tutto il campo visivo, la sua traslucenza ghiacciata percorsa da sottili striature citoschelatrici e percossa da una tensione muscolare a metà della retrazione. Lungo il suo bordo si addensano i colloblasti: cupole emisferiche di adesione biologica, alcune ancora integre e gonfie come perle di rugiada, altre già schiacciate contro la chitina rugosa di un'antenna di *Calanus* — quella trave di ambra ferruginosa che entra nell'inquadratura come un arco architettonico, le sue scanalature trasversali orlate di luce fredda. Dove la superficie del crostaceo ha toccato il tentillum, due o tre teste di colloblasto si sono deformate in dischi piatti che diffondono un alone adesivo appena visibile: una reazione biochimica di ancoraggio irreversibile catturata nel suo mezzo atto, mentre i filamenti spiralati si srotolano come molle microscopiche nell'interno del filo. Una zampa natatoria del copepode — ambra traslucida, piumata di setole — spinge inutilmente contro il tentillum, e la lieve curvatura del filo tradisce il vettore di forza di un animale che non fuggirà. L'intera scena è un teatro di predazione miniaturizzata in cui la biologia del contatto — chimica, meccanica, geometria — si legge nella luce fredda a 480 nanometri che scende da una superficie lontanissima e incide di bianchi riflessi speculari ogni cupola adesiva nell'abisso blu-nero.
