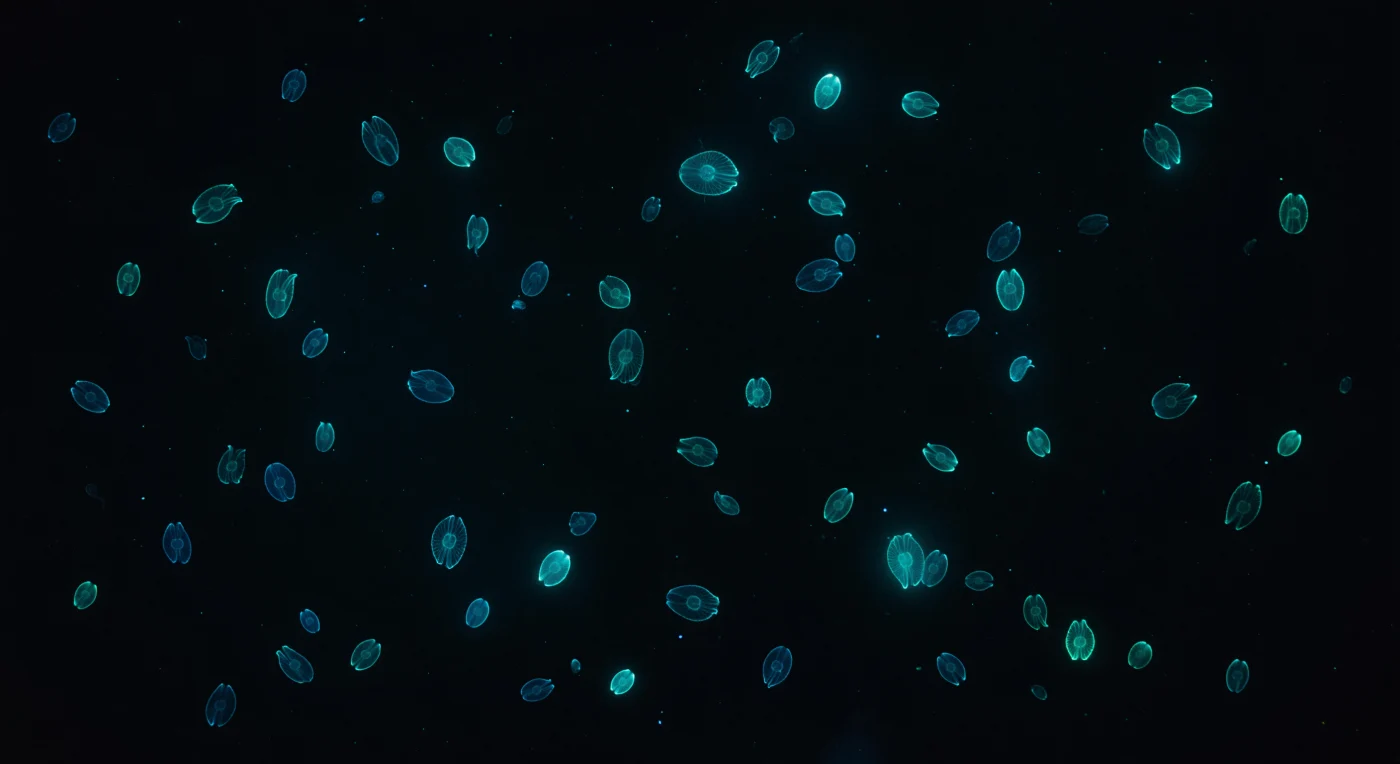
In absoluter ozeanischer Dunkelheit, zwei Meter unterhalb einer mondlosen Meeresoberfläche, verdichtet sich die Welt auf reines Schwarz – durchbrochen von lebenden Konstellationen. Dutzende von *Mnemiopsis leidyi* treiben in alle Richtungen, ihre gallertigen Körper nahezu unsichtbar, fast perfekt im Brechungsindex mit dem umgebenden Salzwasser übereinstimmend, sodass nur acht leuchtende Bänder kalten blaugrünen Feuers bei exakt 490 Nanometern ihre ovalen Umrisse gegen das Nichts zeichnen. Wellen biolumineszenter Aktivierung wandern kontinuierlich vom stumpfen Aboralpol zur Mundöffnung – ein langsamer, hypnotischer Puls, erzeugt durch Photocyten tief im Mesoglea-Gewebe, wo Luciferin-Luciferase-Reaktionen in Echtzeit ablaufen und jeden Kammplattenstreifen für etwa eine Sekunde erleuchten, bevor die Reihe erlischt und sich von Neuem entzündet. Die entfernteren Tiere lösen sich in weiche zyanfarbene Höfe auf, da schwebende Meeresschneepartikel jeden Photon zerstreuen und dem ansonsten leeren Wasserkörper eine fast greifbare Textur und Tiefe verleihen. Man schwebt inmitten eines dunklen Nebels, dessen Sterne alle leben, alle auf kaum wahrnehmbaren Strömungen treiben und ihr eigenes kaltes Licht durch ein Meer tragen, in dem jedes Photon kostbar und bewusst gesetzt wirkt.

Der ROV-Strahl aus blauem LED-Licht schneidet durch das Wasser wie eine Klinge, und aus dem ontologischen Nichts materialisieren sich die Formen von *Bathocyroe fosteri* — dunkelweinrote Rippenquallen, deren pigmentierte Mesoglea so dicht und lichtabsorbierend ist, dass sie im Dunkel schlicht nicht existieren, bis die 470-Nanometer-Wellenlänge ihre Flanken trifft und die karmesinfarbenen Orallappen aufglühen wie glimmende Kohlen in einer geschlossenen Faust. Fünf Tiere treiben in loser Gemeinschaft durch den Lichtkegel, ihre gelatinösen Körper zu gut neunzig Prozent aus Wasser, ihre Mesoglea ein viskoelastisches Gel aus Kollagenfasern und Glykoproteinen mit einem Brechungsindex so nah am Meerwasser, dass sie bei Fehlen von Licht schlicht verschwinden. Entlang ihrer acht Kammreihen brechen die Ziliatenplatten — jede aus Hunderttausenden fusionierter Geißeln gebaut — das einfallende blaue Licht in metachrone Spektralwellen: Violett, Blaugrün, ein flüchtiges Bernsteinleuchten, das als reine Beugungsoptik über lebendige Strukturen wandert, kein Biolumineszenz-Ereignis, sondern physikalische Unausweichlichkeit. Das Meeresschnee füllt den gesamten Lichtkegel mit seiner langsamen Textur — Schleimflocken, Kotpellets, bakterielle Aggregate, die mit etwa einem Meter pro Minute sinken und dem Blick räumliche Tiefe geben wie umgekehrte Sterne in einem schwarzen Universum.

In den oberen zwei Metern des mediterranen Wassers hängt man schwerelos vor einem Wesen, das kaum als Materie existiert: *Bolinopsis infundibulum*, eine lebende Laterne aus nahezu reiner Gallerte, deren Körper so perfekt den Brechungsindex des umgebenden Meerwassers angleicht, dass er nur als zarter optischer Schimmer verraten wird. Was hingegen mit unbändiger Realität brennt, sind die acht Kammreihen – breite Bögen aus zusammengesetzten Ziliatenplatten, die in antiplektischen metachronen Wellen aufleuchten: Tiefkarmin fließt in Bernstein, dann in giftiges Grün, dann in elektrisches Violett, jede Farbe der nächsten nachjagend wie langsames Neon über einen nassen Spiegel, ein biologisches Beugungsgitter, das dreißig Mal pro Sekunde pro Platte neu kalibriert wird. Die oralen Lappen des Tieres fallen in weichen Falten nach vorn, ihr Gewebe zart rosa geflusht von den verzweigten gastrovaskulären Kanälen, die durch die Mesogloea ziehen – ein feines Geäder, das nur im Gegenlicht der Oberfläche sichtbar wird wie Kapillaren in einer ans Licht gehaltenen Hand. Vier Meter tiefer rollt mediterranes Sonnenlicht als goldenes Kaustikengeflecht über sandigen Meeresboden und Posidonia-Debris, das durch die Wassersäule zu etwas Impressionistischem weichgezeichnet wird – einem warmen Untergeschoss, das dem Mittelwasser die Qualität eines lichtdurchfluteten Kirchenschiffs verleiht, offen und enorm relativ zu dem Phantom in seiner Mitte.
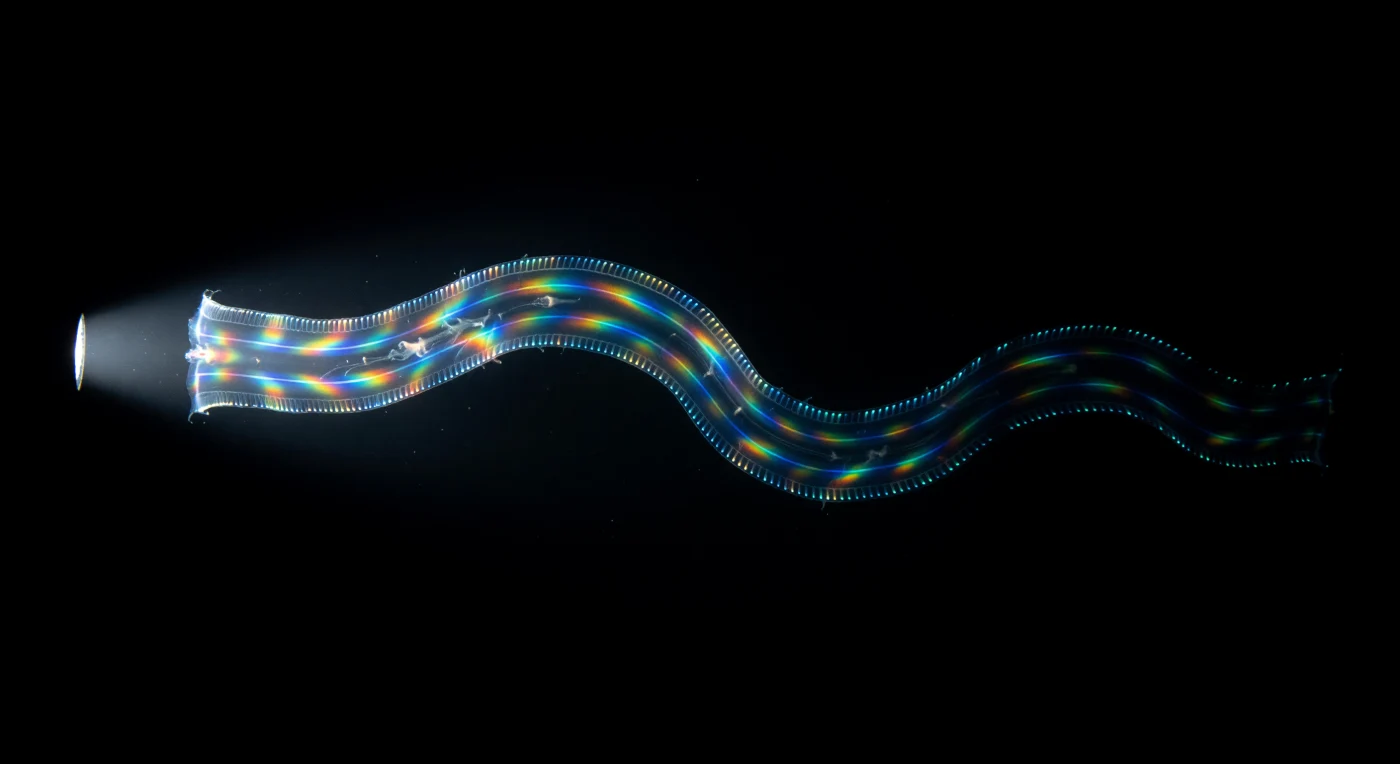
In absoluter ozeanischer Dunkelheit, drei Meter unter der Oberfläche des tropischen Atlantiks, materialisiert sich aus dem Nichts ein flaches, transparentes Band aus Gelatine – *Cestum veneris*, der Venusgürtel, sechzig Zentimeter pures gelöstes Tier, das sich in einer einzigen, langsamen Sinuswelle durch das körperwarme Salzwasser schiebt. Das Tier ist optisch kaum vom Wasser zu unterscheiden, denn sein Mesoglea-Gewebe besitzt nahezu denselben Brechungsindex wie das umgebende Meer; wahrnehmbar bleibt es fast ausschließlich durch seine vier Kammreihen, deren Photocyten ein kaltes Blaugrün bei 490 Nanometern pulsieren lassen – schmale, sequenziell aufflammende Lichtsäume, die die gesamte Kontur des Tieres wie lebendige Neonlinien nachzeichnen und jede Biegung der Sinuswelle als glühendes Atemzeichen im Schwarz sichtbar machen. Als der Taucherscheinwerfer die Fläche trifft, bricht das latente Spektrum auf: Die Kammplatten verwandeln sich in Beugungsgitter in Bewegung, ihre mit 15 bis 35 Hertz schlagenden Zilien zerlegen das weiße Licht in eine rollende Kaskade aus Rot, Gold, Grün und Violett, die in Wellen vom Mundpol zum Aboral-Pol jagt, ein kontinuierlich umschreibendes Farbband, gesättigt und blendend gegen das undurchdringliche Schwarz. Dann gleitet das Tier zurück in die Dunkelheit, und nur die vier biolumineszenten Linien verraten noch seine Existenz, bis auch sie erlöschen und das warme Salzwasser sich wieder schließt, als habe es nie etwas enthalten.

Im fahlen Jadegrün eines schottischen Lochs im Februar treibt man inmitten einer stillen Ansammlung gelatinöser Kugeln, jede kaum so groß wie eine Glasmurmel, deren mesoglea-Körper so vollkommen transparent sind, dass sie sich nur durch eine hauchdünne Brechungsgrenze gegen das trübe, partikelgesättigte Wasser abzeichnen. Die acht Kammreihen des nächstgelegenen Tieres ziehen sich wie Meridiane über die Kugeloberfläche, und wo die Zilienplatten ihren metachronalen Schlag vollführen, läuft ein gedämpftes Schillern hindurch – von altrosa über warmes Bernstein bis zu einem flüchtigen Violettgold –, strukturelle Farbe, die nicht aus Pigmenten, sondern aus dem Lichtspiel Tausender synchron schlagender Zilienpaddel entsteht. In den blassgrünen Leuchtkern, den der Darminhalt hinter der fast unsichtbaren Mesoglea-Wand wirft, laufen feine Tentakeln aus, die sich in fünfzehn Zentimeter lange Gossamer-Fäden auflösen und erst im Partikelschleier verschwinden, ihre colloblastenbesetzten Tentillen nur als silbriger Hauch erkennbar. Vier oder fünf weitere Individuen hängen in verschiedenen Tiefen als blasse, kaum greifbare Formen in der Wassersäule, manche mit einem kurzen rosa Flimmern verraten, andere so nah an der optischen Auflösung des Wassers, dass sie bloß als leichte Dichteanomalie im dunkelgrünen Nichts erscheinen. Am unteren Bildrand gibt ein ledrig-glänzendes Kelpblatt den einzigen materiellen Anhaltspunkt in einer Welt, die fast vollständig aus Wasser und gefrorenem Licht besteht.

In den oberen zwei Metern des Golfstroms, wo spätnachmittägliches Licht die bewegte Oberfläche in ein flimmerndes Netz goldener Kaustiken verwandelt, vollzieht sich eine der stillsten Gewalttaten des offenen Ozeans: Ein *Beroe cucumis* in Lachsrosa und Rosenkoralle hat seinen unwahrscheinlich weit gedehnten Macrostome-Mund um mehr als die Hälfte eines noch lebenden *Mnemiopsis leidyi* geschlossen, dessen freiliegende hintere Kammreihen weiterhin ihre metronomischen Metachronal-Programme abspulen — jede Ktenplatten-Reihe ein schmales Glasruder, das das Abendsonnenlicht in rollende Kaskaden struktureller Farbe bricht, Violett über Bernstein zu Grün in Sekundenbruchteilen, ein unwillkürliches Regenbogenflackern, das nicht weiß, dass sein Signal bereits verloren ist. Durch das nahezu wasserklare Mesoglea des Räubers zeichnen sich die verzweigten Meridionalkanäle wie ein leuchtendes Flussdelta von oben ab — rosafarbene Tributäre in bilateraler Symmetrie, die schwach pulsieren, während die Verdauung beginnt, und das Licht von innen heraus wärmen, als wäre das Gewebe eher getönte Luft als Zellmasse. Die kaustischen Muster der Wasseroberfläche gleiten wie goldene Interferenzwellen über beide Körper, biegen sich um die geschwungene Mesoglea wie um lebende Linsen, und im bläulich-indigofarbenen Wasserraum dahinter treiben transparente Flocken marinen Schnees in kontemplativer Langsamkeit — einziges Maß für die Stille einer Begegnung, die auf keiner anderen Bühne als dem offenen Ozean möglich wäre.

In der samtschwarzen Wassersäule einer warmen Sommernacht in der Chesapeake Bay schweben drei *Mnemiopsis leidyi* wie langsam atmende Kathedralen aus Gallerte, ihre Mesoglea-Wände so dünn und brechungsgleich zum umgebenden Wasser, dass sie weniger als feste Körper erscheinen denn als organische Verzerrungen des Raums. Entlang ihrer acht Kammreihen rollt biolumineszentes Licht in metachronen Wellen — tiefes Kobaltblau, das in Türkis und Aquamarin übergeht — als würden glühende Finger über lebende Lichtleiterkabel streichen, Photozytenmission bei 490 nm, die sich durch Hydrogelwände diffundiert und die verästelten Gastrovaskulärkanäle als gespenstische dunkle Spitzen im Innenlicht freilegt. Aus den Gonoporenspalten entlang der Meridionalkanäle quellen langsame, milchweiße Spermienwolken hervor, die im Licht der eigenen Ctenophoren-Emission kurz silbrig aufleuchten, sich träge mit der Mikroströmung drehen und dort, wo zwei Wolken benachbarter Tiere sich überschneiden, einen perlmuttfarbenen Fortpflanzungsnebel formen, der chemische Signale trägt, die für uns unsichtbar bleiben. Durch diesen pelagischen Schleier taumeln Eier als fast perfekte, hochbrechende Kugeln, die das biolumineszente Umgebungslicht in winzige innere Kaustiken bündeln — gleißende Miniaturgestirne, jedes von einem prismatischen Beugungshalo aus Violett bis Orange umkränzt — und so ein dreidimensionales Universum kalter biologischer Funken bilden, das in der warmen, salzigen Stille dreißig Fuß unter der Oberfläche langsam in die Schwärze auflöst.

Wer von zwölf Metern Tiefe im Hardangerfjord nach oben blickt, sieht eine Kathedrale aus treibendem Glas: Hunderte von *Bolinopsis infundibulum* hängen wie Ornamente im Wasser, ihre ovalen Körper aus Mesoglea – einem viskoelastischen Gel mit nahezu demselben Brechungsindex wie Seewasser – kaum als Materie erkennbar, eher als sanfte Linsenwölbungen, die das Licht um sich herum verbiegen. Acht Kammbänder verlaufen meridional entlang jedes Körpers, und während die metachronen Ziliarwellen Platte für Platte abrollen, erzeugt die Beugung an den fusionerten Cilien ein kontinuierliches Prismenspiel, das von Rubinrot über Goldgelb bis zu tiefem Meeresgrün und weitem Violett kreist – jedes Tier in einer anderen Phase versetzt, sodass das gesamte Ensemble wie ein lebendiger Kronleuchter aus asynchron flammenden Farbstreifen flimmert. Die breiten Volumenstrahlen des norwegischen Sommerlichts, schräg durch das phytoplanktongetrübte Wasser fallend, werden von jedem Kammband spektral gestreut und zerfächern sich in prismatische Fächer, die sich zwischen den Körpern überschneiden und die Wassersäule mit einem diffusen chromatischen Dunst füllen. Einzelne Tiere tragen einen warmen Bernsteinschimmer durch ihre Gastrovaskulalkanäle – der durchscheinende Apricotton kürzlich verschluckter Copepoden, sichtbar wie ein Eigelb hinter einer Eierschale, die ans Licht gehalten wird. Weit oben liegt die Oberfläche als silbernes, von Kaustiknetzmustern gekräuseltes Spiegelglas, und in der Tiefe darunter werden die Tiere zur blauen Andeutung: blasse Ovale mit acht schwach leuchtenden Streifen, ihr kollektives Schimmern kaum noch vom Dunkel zu trennen.

Im Lichtkegel der blauen ROV-Scheinwerfer taucht aus absolutem Schwarz ein unmögliches Tier auf: *Lampocteis cruentiventer*, die Blutrote Rippenqualle, hängt reglos drei Körperlängen vor der Kamera in tausend Metern Tiefe des Monterey-Canyons, ihr scharlachrotes Mesoglea-Gel so gesättigt, dass es fast chirurgisch wirkt gegen die Finsternis um sie herum. Die breiten Mundlappen hängen geöffnet und entspannt nach vorn, wie die samtigen Blütenblätter einer Tulpe in voller Blüte, ihre Oberränder fangen das blaue LED-Licht und werfen einen zyanfarbenen Glanzstreifen, bevor die Helligkeit in weinroten Schatten versinkt. Acht Kammreihen zeichnen sich als kaum dunklere Längsrippen über den roten Körper – die einzelnen Ctenophoren-Platten, jede ein Bündel aus Hunderttausenden fusionierter Geißeln, bleiben bei diesem eingefrorenen Blick unsichtbar, hinterlassen aber eine zarte, gerippte Textur, die das schrägfallende Licht in Mikroschaten verwandelt. Mariner Schnee – Partikel organischen Trümmermaterials von staubfein bis reiskorngroß – treibt stetig durch den Lichtkegel auf unterschiedlichsten Vektoren, gibt dem schwarzen Wasser messbare Tiefe und erinnert daran, dass dieser Körper, der zu 97 Prozent aus Wasser besteht und kaum schwerer als das Meer selbst ist, in einer Strömungslandschaft aus fallendem organischen Material existiert, in der er als lautloser, gallertiger Räuber auf Beute wartet.

Der Blick fällt auf eine Kuppel aus biologischem Glas, kaum zweihundert Mikrometer im Durchmesser, die sich wie ein Dom aus einem einzigen Wassertropfen über dem aboralen Pol von *Mnemiopsis leidyi* erhebt — ihre Wandung so optisch homogen, dass sie sich nur durch einen hauchdünnen Kaustikbogen aus konzentriertem Blau-Weiß verrät, dort wo das Meerlicht sich minimal durch Kollagen und Glykoprotein bricht. Im Innern schwebt eine cremefarbene Masse aus Kalziumkarbonat-Granulat — der Statolith — in vollkommener Stille, matt-weiß wie pulverisierte Kreide gegen die umgebende Transparenz, von vier Fächern schlagender Balancierciilien umgeben, die als schimmernde Schleierschleier vibrieren, ihre Schlagfrequenz weit jenseits jeder eingefrorenen Momentaufnahme. Von der Kuppelbasis strahlen vier zilienbesetzte Furchen wie die Nähte einer Kompassrose nach außen, ihr kollektives Flimmern erzeugt ein kaum wahrnehmbares irisierendes Aufleuchten — flüchtige Grüns und Silbers an den Rändern, wo Beugungslicht im Streifwinkel gebrochen wird. Das Mesoglea-Substrat darunter erscheint als weite Ebene aus wasserklarem Silikongelée, ihre Tiefe in blauem Dunst verloren, während der offene Ozean jenseits des Tierkörpers als leuchtend tiefblaues Nichts rahmt, das diese kristalline Architektur gleichzeitig intim und in unendlichem Raum schwebend erscheinen lässt.

Vor uns schwebt eine kaum sichtbare Glaskugel — die zwei Millimeter kleine Cydippid-Larve eines Rippenquallen, so optisch dem Meerwasser angeglichen, dass sie eher als zarte Linsenverzerrung erscheint als als festes Wesen, während das blassgoldene Dämmerlicht durch die Oberflächenmikroschicht herabfiltert und sich in acht rudimentären Kammreihen zu flüchtigen prismatischen Blitzen bricht, die in Rosé und Violett aufleuchten und wieder verschwinden. Aus unserer auf Sandkorngröße geschrumpften Perspektive treibt eine Kette von Chaetoceros-Diatomeen vorbei wie ein langsamer Güterzug aus lebendem Glas — jede Siliziumzelle so groß wie eine Melone, bernstein- und ockerfarben leuchtend dort, wo Morgenlichtstrahlen auf chloroplastgefüllte Frusteln treffen, verbunden durch glasklare Dornen, die das Licht in prismatische Nadeln zersplittern. Ein Copepoden-Nauplius rotiert in der Ferne auf seiner Achse wie ein fremdes Raumschiff aus kupferrotem Chitin, während überall Meeresschnee driftet — transparente Schleimflocken, Detrituspartikel und Phytoplanktonzellen in Gold, Olivgrün und warmem Bernstein, von oben sanft beleuchtet in jenem diffusen aquatischen Licht, das keine harten Schatten kennt. Die Larve hängt genau an der Grenze zwischen der erleuchteten Oberflächenwelt und der sich verdunkelnden Tiefe, ihre beiden haarfeinen Tentakel bereits ausgestreckt in diese lebendige Suspension hinein, taktend nach einer Beute, die in diesem Stadium ihres Lebens kaum kleiner sein könnte als sie selbst.

Vor dir hängen drei *Mnemiopsis leidyi* reglos im offenen Wasser, ihre ovalen Körper aus nahezu reiner optischer Transparenz genau entlang jener horizontalen Schimmerlinie aufgespannt, die die gesamte Sichtweite von Wand zu Wand durchzieht: die Thermokline, sichtbar als ein flimmerndes, wärmeflirrendes Linsenband, das warmes, ceruleanblaues Wasser über sich von kaltem, grünlich-dämmrigem Wasser darunter trennt. Jede Qualle stradelt diese Dichtegrenze mit der Präzision eines Auftriebsmeisters — ihre obere Hälfte badet in gefilterten, schräg einfallenden Goldlichtkegeln, die vierzig Meter Meerestiefe zu breiten Scheiben weichen Lichts aufgeweicht haben, während ihre untere Hälfte bereits in jenem kühlen, partikelreichen Zwielicht versinkt, das das Licht nicht mehr vollständig durchdringt. Entlang jeder der acht Kammreihen rollen in langsamen Wellen Strukturfarben aboralwärts: Rubinrot gleitet in Bernstein, Bernstein in Säuregrün, Grün in Indigo, die Sequenz das gesamte sichtbare Spektrum in unter einer Sekunde durchlaufend, da jede Ktenplatte — ein aus Tausenden verschmolzener Cilien gefügtes Beugungsgitter — das einfallende Licht in seine Bestandteile aufbricht. Die weichen Oralloben hängen offen in die kältere Zone hinein, ihre Cilien ziehen unsichtbare Mikroströmungen aufwärts, und in der grünlichen Tiefe darunter treiben Dutzende orangeroter Copepodenpünktchen wie Glut in einem Ozean aus Kälte und Stille.

Der Blick schweift durch einen absoluten Schwärze, die kein atmosphärisches Dunkel ist, sondern das geschlossene Vakuum eines optischen Systems – darin hängen vereinzelte blaue DAPI-Punkte wie kalte Sterne in einer fernen Galaxie, jeder Zellkern ein harter, klar begrenzter Himmelskörper, schwebend ohne Gravitation im gallertigen Mesoglea-Volumen einer zwei Millimeter kleinen Mnemiopsis-Larve. Von dieser Leere aus strahlt ein nicht-hierarchisches Netz magentafarbener FMRFamid-markierter Nervenfasern in alle Richtungen gleichzeitig, ohne Mittelpunkt, ohne Hierarchie – jede Faser glüht in sattem Korallenrot, verdickt sich an Kreuzungspunkten zu leuchtenden Knoten und wirft weiche Rosahöfe ins Dunkel, eine neuronale Architektur, die älter ist als jedes zentrale Nervensystem der Erde. Acht Bögen aus reinem Säuregrün wölben sich wie Kathedralen-Rippen über das Gesichtsfeld – die serotoninpositiven Kammreihen-Bänder, scharf begrenzte Meridiane aus Jade und Limette, die räumliche Ordnung in das anarchische Fadenwerk zwingen und sich in steilen Kurven zu einem einzigen Brennpunkt am Zenit zusammenfinden. Dort brennt das Apikalorgan als perfekter kreisförmiger Heiligenschein aus verschmelzendem Magenta und Grün, ein Statocyst-Rosett, dessen Gleichgewichtssinn die Schwerkraft für ein Tier erspürt, das zu neunzig sieben Prozent aus Wasser besteht und dessen gesamter Körper einen Brechungsindex besitzt, der sich kaum vom umgebenden Meer unterscheidet.

In der blauen Stille des Kreisel-Beckens schwebt ein einziges Tier wie eine gläserne Skulptur – *Mnemiopsis leidyi*, sechs Zentimeter lebendige Transparenz, deren Körper das kalte LED-Licht von unten so aufnimmt, dass das Mesogleawasser selbst zu leuchten scheint. Entlang der acht Kammreihen zieht ein irisierendes Flackern in metachronen Wellen: Jede Wimperplatte – ein gefedertes Paddel aus Zehntausenden synchron schlagender Cilien – bricht das Licht in ein kurzes Aufblitzen von Orangerot über Smaragdgrün bis zu kühlem Violett, reine Geometrie als Spektralfeuer. Die Mesoglea ist kein leerer Raum, sondern ein viskoelastisches Gel aus Kollagenfasern und Glycoproteinen, dessen Brechungsindex so nah an dem des Meerwassers liegt, dass das Tier erst durch die aprikosenrosa Gonadenstreifen entlang des Stomodäums sichtbar wird – warme, granulare Bänder organischer Dichte gegen das kalte Blau. Die Oralloben pulsieren in langsamer Peristaltik, ihre Ränder fransig und lichtdurchlässig wie gefrostetes Glas, während im Hintergrund die geschwungene Tankwand als dunkler Horizont das Tier in seiner privaten Welt einschließt.

In diesem eingefrorenen Augenblick tief im mesopelagischen Dunkel schwebst du unmittelbar neben einem einzelnen Tentillum von *Pleurobrachia* — einem lateralen Ast kaum dicker als ein Glasfaden, der sich wie ein Frostkristallarm in die schwarzblaue Wassersäule erstreckt. Entlang seiner eisblau-transparenten Länge drängen sich die Colloblasten: jede eine perfekte Halbkugel, deren refraktile Granulaköpfe das spärliche Umgebungslicht als kalte weißgoldene Funken zurückwerfen, während die spiralförmigen Filamente hinter jedem ausgelösten Kopf wie aufgezogene Miniaturfedern in das Innere des Strangs verschwinden — das zelluläre Fangsystem eines Tieres, das zu 97 % aus Wasser besteht und dennoch mit biochemischer Unerbittlichkeit zugreift. Oben rechts im Bild bricht die Antenne eines *Calanus*-Ruderfußkrebses ins Bild ein wie ein verrostetes Architekturrelikt aus Chitin: warm-bernsteinfarben, transversal gerippt, mit Setzulae besetzt — und dort, wo sie das Tentillum berührt hat, sind zwei oder drei Colloblasten zu flachen Scheiben zerdrückt, ihre Adhäsionsmoleküle bereits irreversibel mit der Cuticle verbunden. Ein Schwimmbein des Krebses fächert kraftlos gegen den Strang, die einzige Bewegung in dieser gestoppten Szene ist das infinitesimale Verbiegen des Tentillums unter dem Gewicht der Beute, bevor die Tentakelmuskulatur mit langsamer, unaufhaltsamer Kontraktion zurückzieht — in der Zeitskala von einer bis zehn Sekunden wird der Copepode eingeholt sein.
