Wetenschappelijke betrouwbaarheid: Zeer hoog

De blik vanuit het binnenste van de bacteriële celwand onthult een wereld die meer op een gotische kathedraal lijkt dan op een biologische structuur: een ondoordringbaar labyrint van verweven glycaanketens in warme amber- en siënakleuren, onderbroken door kortere, ivoorkleurige peptidebruggen die het geheel vergrendelen tot een driedimensionaal netwerk van verbazingwekkende dichtheid. Dit is de peptidoglycaanmantel van *Bacillus subtilis*, een Gram-positieve bacterie, waarvan de celwand twintig tot tachtig nanometer dik is en als primaire drager van de turgordruk fungeert — een compressieve kracht die het inwendige van de cel bij elkaar houdt tegen een osmotisch verschil dat meerdere atmosferen kan bedragen. Tussen de glycaanstaven hangen donkergroene teichoinezuurketens als slingers van zeewier, hun anionische ruggengraten geladen genoeg om de passage van ionen en moleculen door de wand te sturen en te filteren. Ver onder dit amber gewelf gloeit het binnenste membraan als verwarmde olie, zijn vloeibaar-kristallijne oppervlak bezaaid met eiwitcomplexen die energie omzetten en stoffen transporteren. Naar boven toe rafelt de wand uit in steeds lossere, blekere draden die oplossen in de koele, blauwe stilte van de extracellulaire wereld — een grens die niet scherp is, maar geleidelijk vervaagt, zoals een kustlijn die verdwijnt in de mist.
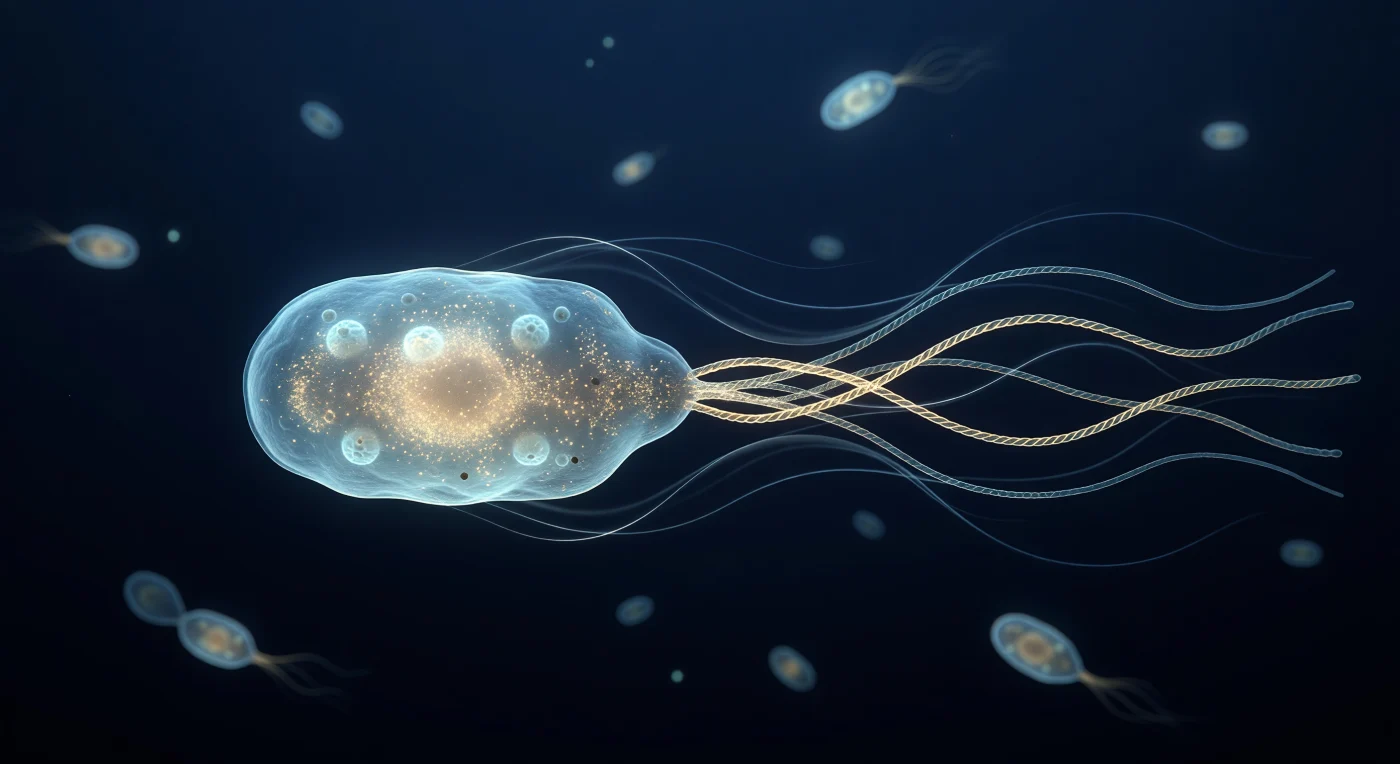
Je zweeft vlak achter een *E. coli*-cel waarvan het blekecyaan doorschijnende lichaam het beeldveld vult als de romp van een traag varend schip — het buitenmembraan glimt met een fijne reliëf van porine-rozetten die het binnenvallende licht opvangen als kleine kuiltjes in een levende mantel, terwijl de cytoplasmische binnenkant oplicht als een amberkleurig nevelwoud van tienduizenden ribosomen. Vier helische flagellaire filamenten hebben zich achter de cel ineengevlochten tot één superhelicaal bundel dat als een zachtgouden tros wegdraait in het donkere, marineblauw aquatische universum — ronddraaiend met honderden toeren per seconde in een wereld waar inertie volkomen betekenisloos is en viscositeit alles beheerst. De omringende vloeistof draagt haar eigen getuigenis van deze laminaire Stokes-stroming: zilveren sluiers krullen in symmetrische bogen vanuit de bundel naar voren, als zijdedraad in glycerol, elk spoor een bewijs dat elke beweging hier onmiddellijk stopt zodra de roterende machinerie stilvalt. In de diepte van het nachtblauwe veld drijven vage bacteriële silhouetten als gloeiende sintels in een donkere lucht, elk omgeven door een nauwelijks zichtbare capsulaehalo — een stille herinnering aan de immense dichtheid van dit microscopische universum, waar elke oppervlakte geladen is en het enige licht het koude zelfgloeien is van moleculaire biologie die haar oeroude werk doet.

Je zweeft in het absolute centrum van een levende *E. coli*-cel, volledig omgeven door een verstikkend dicht woud van donkerblauwe DNA-strengen die zich plectonemisch opwinden in ingewikkelde, vertakkende lussen — samen zo'n vier tot vijf miljoen basenparen DNA samengeperst in een volume kleiner dan een kubieke micrometer. De strengen zijn bezaaid met kleine geelwitte knooppunten van nucleoïd-geassocieerde eiwitten zoals HU en Fis, die scherpe knikken in het DNA-touw klemmen en het koude blauwe schijnsel van de omringende vezels terugkaatsen als warme amber-gouden sterretjes verspreid door een diepe nevel. Geen enkele kijklijn is vrij: elke richting eindigt op korte afstand tegen een nieuwe laag vloeiende lussen, en de diepte lost op in reeksen van zachter wordende indigoblauwe vlakken die vervagen tot blauwzwarte duisternis, terwijl de viskeuze, gelachtige cytoplasmavloeistof tussen de lussen schemert als bewogen aquamarijn glas. Aan de rand van deze nucleoïdwolk, waar de DNA-dichtheid afneemt, drijven donkerbruine ribosoomkorrels als koolstofbollen in de iets opener cytoplasmatische zone nabij het binnenste membraan, licht wazig door de brekende werking van het dichte biologische medium. Het geheel voelt als zweven binnen een samengeperste, gloeiende storm van georganiseerd leven — een wereld zonder lege ruimte, zonder stilte, enkel de ondoorgrondelijke dichtheid van een functionerende biologische machine.

Je zweeft amper anderhalve handbreed boven een wereld die volledig van lipiden en eiwit is gemaakt — de buitenmembraan van een levende *Escherichia coli*, een golvend landschap van lipopolysacchariden dat zich uitstrekt tot aan een licht gebogen horizon, als het oppervlak van een kleine, warme planeet. Overal om je heen rijzen bèta-barrel porine-trimeren op als obsidianen torens, met geometrische precisie uit de membraan geboord, hun holle interieurs donkere schachten die recht door de dubbellaag naar de periplasmatische ruimte eronder leiden. De kristallijne Lipid A-vlakken tussen de torens flikkeren met een interferentieschittering die ergens zweeft tussen gepolijst obsidiaan en de iriserend chitine van een kever, noch volledig vloeibaar noch volledig vast, gevangen in een liquid-crystallijne toestand die slechts bestaat door de moleculaire geometrie zelf. Aan de verre horizon domineert het flagellaire basaallichaam het tafereel als een industrieel monument — gelaagde eiwitringen die met chirurgische regelmaat zijn gestapeld, de bovenste ring vlak ingebed in de membraan, de haak erboven verdwijnend in de ionenwolk die als een diffuse nevel boven het oppervlak hangt. Het gehele landschap baadt in een koud, blauwwit schijnsel dat van onderen door de half-transparante membraan sijpelt en elke schaduw verzacht tot de stille gloed van een gletsjerfjord in het eerste ochtendlicht.

De beschouwer bevindt zich volledig ondergedompeld in het cytoplasma van een bacterie, net voorbij de vage, vezelige rand van het nucleoïd, en wat zich in alle richtingen ontvouwt lijkt minder op een vloeistof dan op een levende steen — een amberkleurig, harsachtig medium zo verzadigd met macromoleculen dat geen enkel leeg volume zichtbaar is. Ribosomen vullen het volledige gezichtsveld als donkere, nauw opeengepakte bollen van diepgrijs tot warm oker, van elkaar gescheiden door nauwe spleten van slechts vijf tot tien nanometer, zodat het geheel eerder doet denken aan het metselwerk van een oeroude muur dan aan de binnenzijde van een cel. Door deze granulaire dichtheid weven polysome-ketens als onregelmatige parelnoeren — zes ribosomen geregen langs een nauwelijks zichtbare mRNA-draad, een bleekcrèmekleurig, lichtgevend filament dat doorbuigt onder zijn eigen moleculaire gewicht en in het warme bruine waas verdwijnt. Links domineert een GroEL-chaperonine de scene als een massief, gesegmenteerd vat van leisteengrijs, zijn dubbelring-architectuur zo groot ten opzichte van de omringende ribosomen als een stenen zuil ten opzichte van keitjes, zijn centrale holte openend naar een iets koeler, zwak oplichtend interieur. Hier en daar flikkert een GFP-gemerkt eiwit als een smaragdgroene vlam op — de enige kleurbreuk in een wereld die verder volledig in amber en duisternis is verzegeld.

De blik zweeft boven een zwijgende stad die nooit is gebouwd maar is gegroeid: van hieruit strekken zich koepelvormige microkolonie-torens uit, elk opgetrokken uit duizenden stafvormige *Pseudomonas*-cellen die zo dicht opeengepakt zijn dat hun gebogen oppervlakken glinsteren als natte keramische tegels, badend in een koel jadegroen GFP-schijnsel dat van binnenuit lijkt te komen. Tussen de torens snijden donkere, inktzwarte waterkanalen met hydraulische precisie door het landschap — microfluidische stroomgeulen die de kolonie van zuurstof en voedingsstoffen voorzien en afvalproducten afvoeren, een circulatiesysteem dat niet is ontworpen maar is ontstaan door collectief bacterieel gedrag. De ruimte tussen de torens is gevuld met een amberkleurig, halfdonorzichtig extracellulaire polysaccharidematrix — een taai, viskeus gel dat de gemeenschap bijeenhoudt, haar beschermt tegen antibiotica en afweerreacties, en hier en daar oplicht in een warme oranje autofluorescentie alsof er smeulende kolen diep in het materiaal verborgen liggen. Diep in de basale lagen, verborgen onder het groene bladerdak van de rijpste torens, gloeien propidiumjodide-rode vlekken: cellen waarvan de membranen het hebben begeven in de hypoxische schaduw van hun eigen kolonie, stervend in stilte terwijl de wereld boven hen bloeit. Dit is geen metafoor voor een stad — dit ís een stad, gemeten in tientallen micrometers, volledig bewoond en aardedonker van leven.
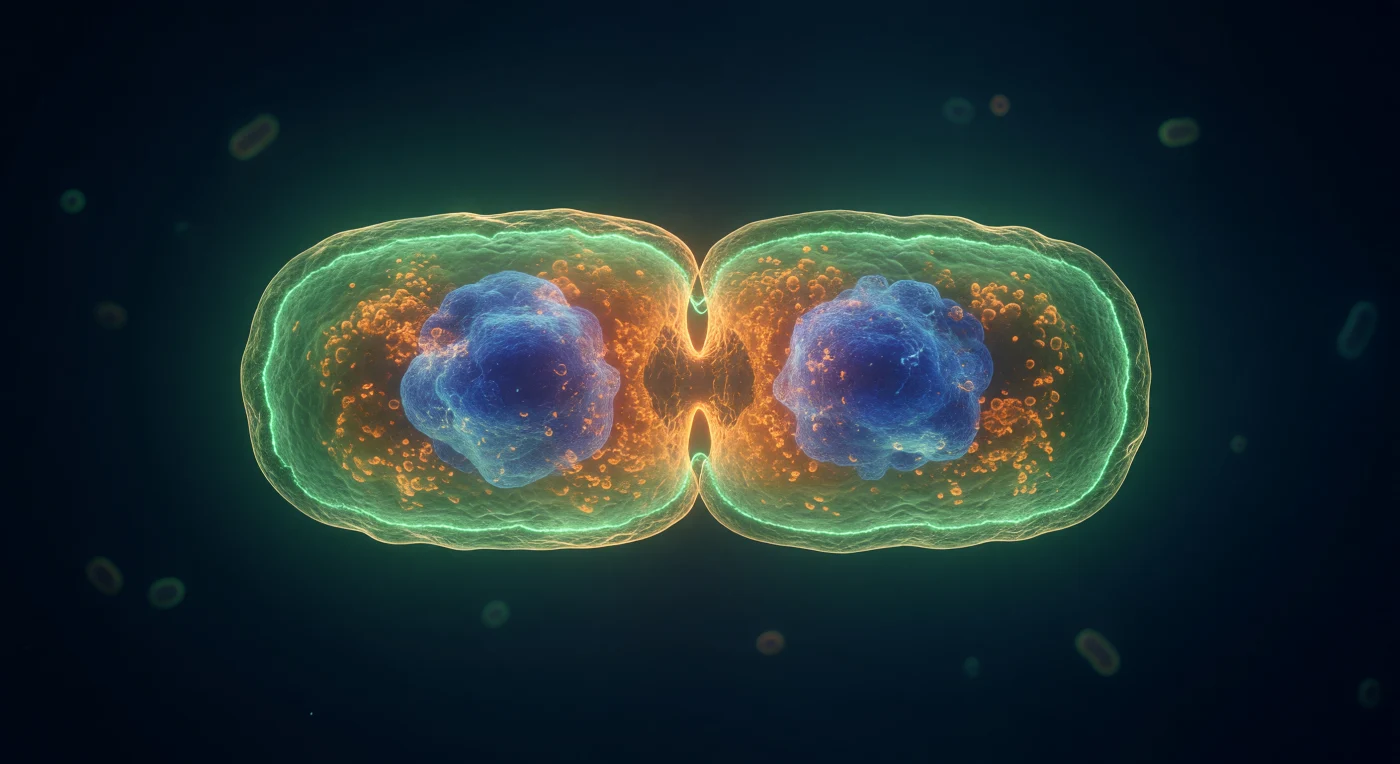
Op de equatoriale gordel van deze reusachtige, langzaam samenknijpende bacteriewereld cirkelen we als getuigen van één van de meest fundamentele gebeurtenissen in de biologie: de celdeling van *Escherichia coli*. Centraal in ons blikveld brandt de FtsZ Z-ring als een kroon van elektrisch groen vuur rondom de volledige omtrek van de versnelling — een continue band van gepolymeriseerde FtsZ-eiwitten, slechts enkele tientallen nanometers breed, die als een moleculaire trekkoord de cel naar binnen trekt via GTP-hydrolyse en dynamische filamentreorganisatie. Door de doorschijnende celwand heen — een goudkleurige lamel van peptidoglycaannetwerk en dubbele membraanlagen — onthult het celinterieur zijn twee gesegregeerde bestemmingen: dichte kobaltblauwe nucleoïdmassa's, volledig uit elkaar gedreven door ParABS-segregatiemachines en chromosomale condensatie, gloeien als compacte nevels in elk ontluikend dochterdeel. De warme, amberkleurige nevel van ribosomen vult elke tussenliggende ruimte met de visuele dichtheid van doorschijnend hars, terwijl de FM4-64-gestempelde binnenmembraan zijn oranje dubbele curve vormt langs de insnoering — een thermodynamisch onvermijdelijk moment, bevroren in fluorescerend licht, waarop één leven in tweeën splitst.

Terwijl je oog vangt wat zich voor je uitstrekt, rijst een rij van vijftien pikzwarte kuboctaëdrische magnetietkristallen op als een monument van geologische precisie, elk ruwweg vier keer jouw eigen hoogte, hun gefacetteerde mineraaloppervlakken een diepblauw-violet metaalglans werpend door het troebele, amberkleurige cytoplasma. Om elk kristal omsluit een doorschijnende lipidenvesikel het donkere mineraallichaam als een gesluierde glazen mantel, zijn dubbele membraanlaag zachtjes rimpelend door thermische agitatie — een nauwelijks zichtbare echo van de Brownse onrust die de hele cel doordringt. Onder de ketting loopt het MamK-cytoskeletfilament als een elektrisch kobaltblauwe rail over de volle lengte van de kristallenreeks, een gevlochten eiwitstructuur die de magnetosomen in hun perfecte lineaire ordening verankert en het geheel de suggestie geeft van een door de cel zelf gebouwde kompasnaald. De achtergrond is geen leegte maar een dichte, half-ondoorzichtige suspensie van ribosomen — matte, onregelmatige bolletjes die als donkere keien in de ambergouden vloeistof drijven en de zichtlijn tot slechts enkele celdiameters beperken. Verder weg welft het binnenste membraan als een glanzende goudbruine grens weg naar het duister van de buitenwereld, waar de cel ophoudt te bestaan en het koude, magnetisch georiënteerde water buiten elke illuminatie onttrekt aan het oog.
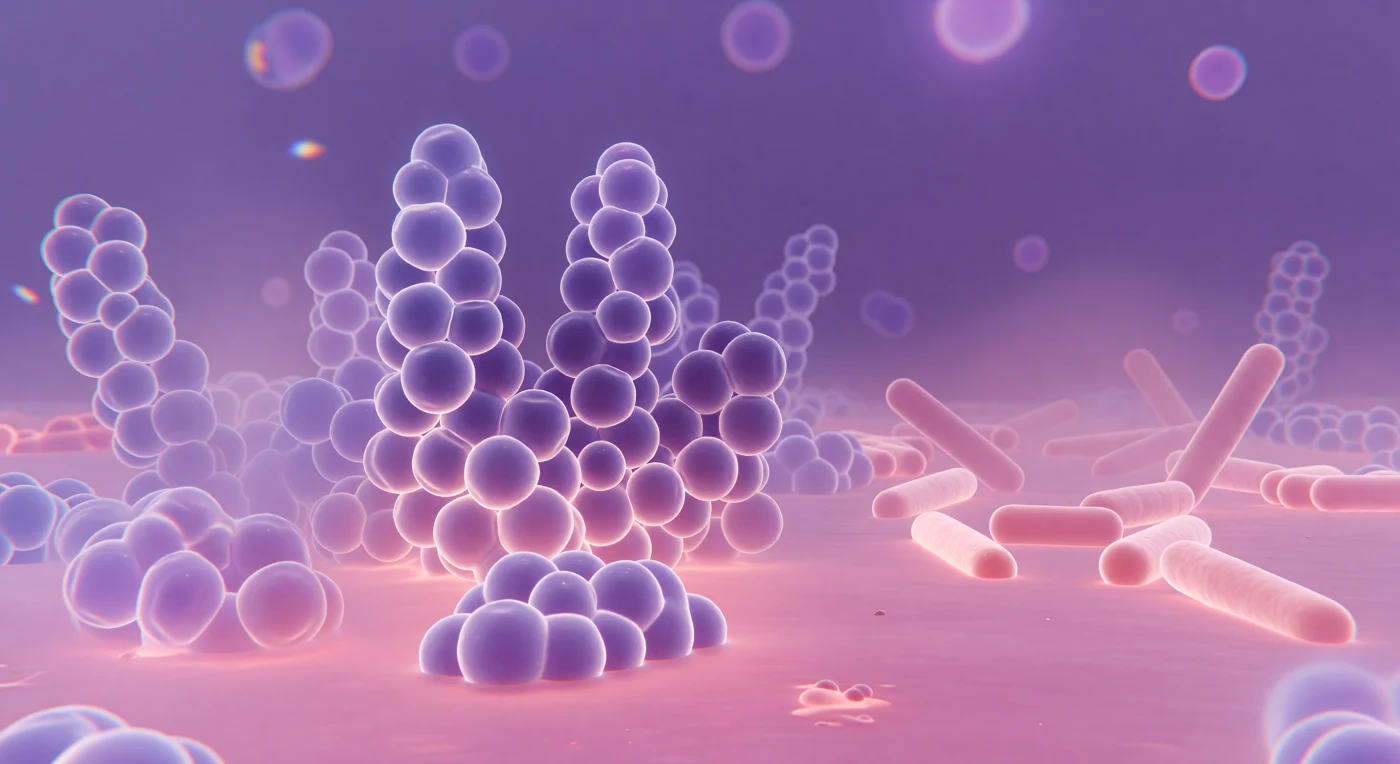
Op de grens van wat licht nog kan onthullen, strekt zich een immens landschap uit in diepe violet-paarse tinten — reusachtige bolvormige structuren rijzen op in druiventros-achtige clusters, elk individu opgebouwd uit een dikke peptidoglycaanwand die het kristalviolet van de Gram-kleuring vasthoudt als een spons die kleur absorbeert. Dit zijn *Staphylococcus aureus*-cellen, Gram-positieve kokken van ongeveer één micrometer doorsnede, waarvan de celwanden tot tachtig nanometer dik zijn en zo rijk aan mureine netwerken dat zij de violette kleurstof behouden zelfs na het ontkleuren met alcohol. Aan de horizon, naar rechts, verspreiden zich lichtroze, langgerekte vormen over een safranine-roze vlakte — *Escherichia coli*-staven die, als Gram-negatieve bacteriën met hun dunnere peptidoglycaanlaag, het violet niet vasthouden en slechts de tegenkleuring opnemen, hun contouren licht vervaagd omdat de golflengte van zichtbaar licht hier nauwelijks kleiner is dan de cellen zelf. De diepte lost snel op: cellen buiten het haarscherpe focusvlak vervagen tot spookachtige lavender-aureolen, en aan de rand van het gezichtsveld flikkeren zwakke chromatische ringen van blauw en amber — de eerlijke optische imperfecties van glas die herinneren aan de absolute fysische grens waarop dit microbiële universum voor menselijke ogen toegankelijk wordt.
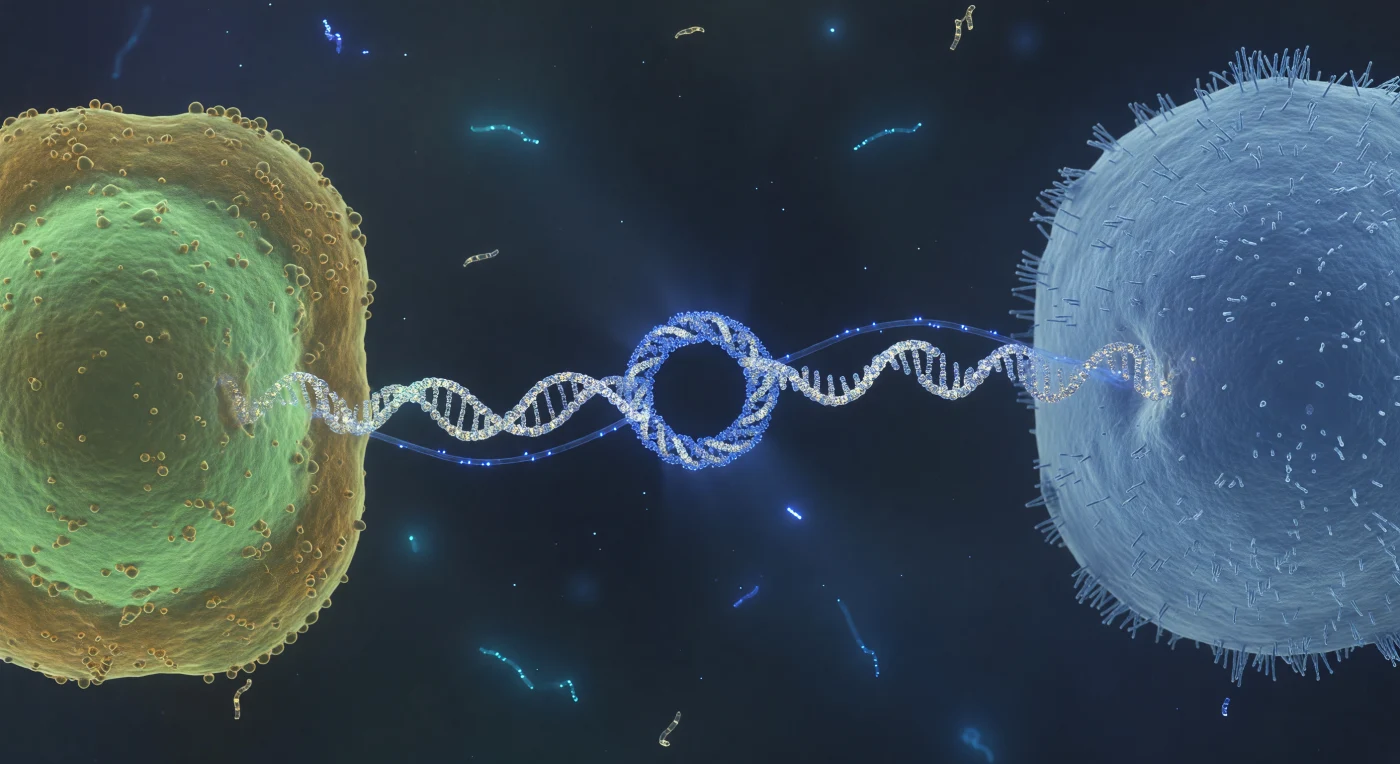
De blik zweeft op ooghoogte met de conjugatiebrug zelf — een witte-gouden eiwitkabel van nauwelijks acht nanometer breed, gespannen als een ankertouw tussen twee reusachtige cellulaire werelden die zich links en rechts uitstrekken tot aan de rand van het zichtbare. Links gloeit de donorstaf in koele celadoongroen, zijn buitenmembraan gegolfd als amber-goud folie bezaaid met porinekronen en de ruwe reliëfs van lipopolysaccharidekoppen; rechts tornt de recipiëntcel op als een matblauw monoliet, zijn oppervlak dicht bezet met fimbriae als een bevroren veld van eiwitstekels. Door het nauwe kanaal van de pilus pulseert een elektrisch-blauwe draad van enkelstrengs DNA — geen scherp object maar een levend licht dat zijn randen uitwaseert in het oplosmiddel, zoals een glasvezelkabel blauw bloeit in het donker — een moleculaire overdracht waarbij genetische informatie van de ene cel in de andere wordt geschreven via een eiwitbrug dunner dan een ribosoommiddellijn. In de verte drijven cyaankleurige slierten vrije DNA-fragmenten geluidloos door het bijna-zwarte medium, meegevoerd door de thermische onrust van een wereld waar de vloeistof aanvoelt als siroop en afstand wordt gemeten in eiwitdiameters.
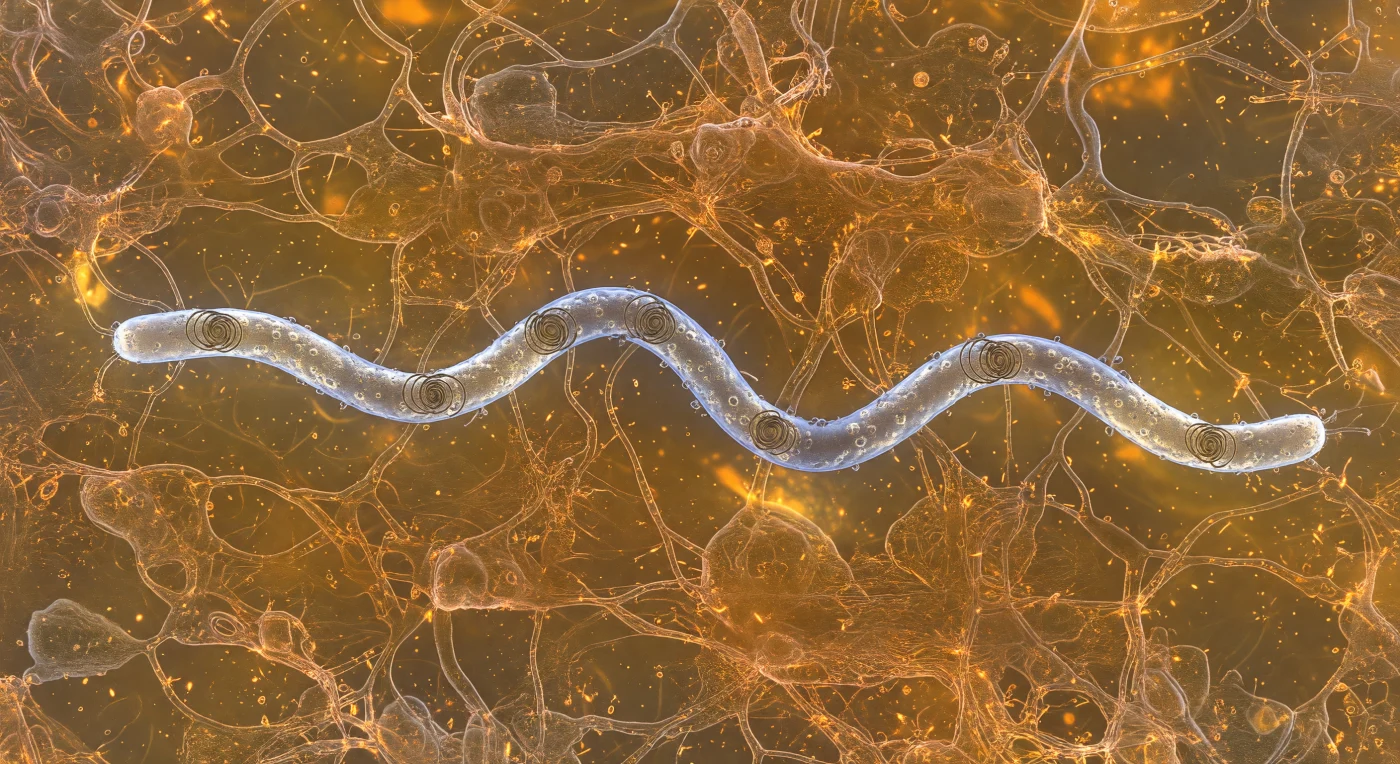
Je beweegt je niet — je hangt roerloos in een amber-oranje wereld van viscoelastisch slijm, omringd door een driedimensionaal netwerk van glycoproteïne-vezels die glinsteren als koperen draden in diffuus bioluminescerend licht, elk strengetje strak en kristalhelder op de voorgrond terwijl de diepte vervaagt in een warme bruingouden nevel. Vijf micrometer links van jou snijdt een *Borrelia burgdorferi*-spirocheet door dit gel-medium: een bleekzilverblauwe kurketrekker van achttien micrometer lang, waarvan het buitenmembraan een koude irriserende glans terugwerpt terwijl de periplasmatische flagellen — nooit blootgesteld aan het medium — als donkere helische kabels zichtbaar zijn onder de buitenste schede en de cel als geheel in zijn karakteristieke vlakke golfbeweging buigen. De bacterie baant zich voort door de mucine-matrix via een mechanisme waarbij de buitenmembraaneiwitten, in samenspel met de intern aangedreven gecoördineerde flagellaire rotatie, het viscoelastische netwerk uiteen doen wijken in een zachte V-vormige kielzog: de glycoproteïnestrengen buigen opzij en sluiten zich daarna langzaam — als stroop — achter de cel, oplichtend in nieuwe goud-koperen reflecties. Dit is het regime van extreem lage Reynoldsgetallen, waar traagheid niet bestaat en beweging op stel en sprong stopt zodra de aandrijving wegvalt — een biologische wereld volledig beheerst door viscositeit, moleculaire dichtheid en de voortdurende, onzichtbare ruis van thermische beweging.

Vanuit het binnenste van *Synechocystis* ontvouwt zich een wereld die aanvoelt als een levende kathedraal: concentrische membraanlagen strekken zich in alle richtingen uit, gloeiend van dieprood chlorofylfluorescentie die het hele interieur doordrenkt met een wijndonkere warmte, als kolen die smeulen onder doorschijnende huid. De thylakoïdmembranen — gestapeld in parallelle golven van slechts enkele nanometers van elkaar — zijn bezet met koraloranje fycobilisome-antennestructuren die als barokke koepels uit het lipidedubbellaag oprijzen en het rode licht zijwaarts verstrooid terugkaatsen in amber aureolen; deze complexen fungeren als moleculaire lichtvangers die fotonen doorgeven aan de fotosystemen diep in het membraan. Zwevend in de cytoplasmatische ruimte daarbuiten hangen icosaëdrische carboxysome-omhulsels — bleke, ivoorkleurige geometrische schalen van 150 nanometer breed, gevuld met RuBisCO-enzymen die kooldioxide concentreren voor de Calvincyclus, elk verlicht als een bevroren glazen lantaarn in het amberkleurige medium. Het cytoplasma zelf is geen lege vloeistof maar een dicht, gelachtig milieu, ribosomaal korrelig en doortrokken van warmte-uitwisseling op moleculaire schaal, terwijl aan de verre celgrens een zachte blauwgroene gloed de buitenwereld verraadt — een aquatische planeet, gezien van binnenuit.
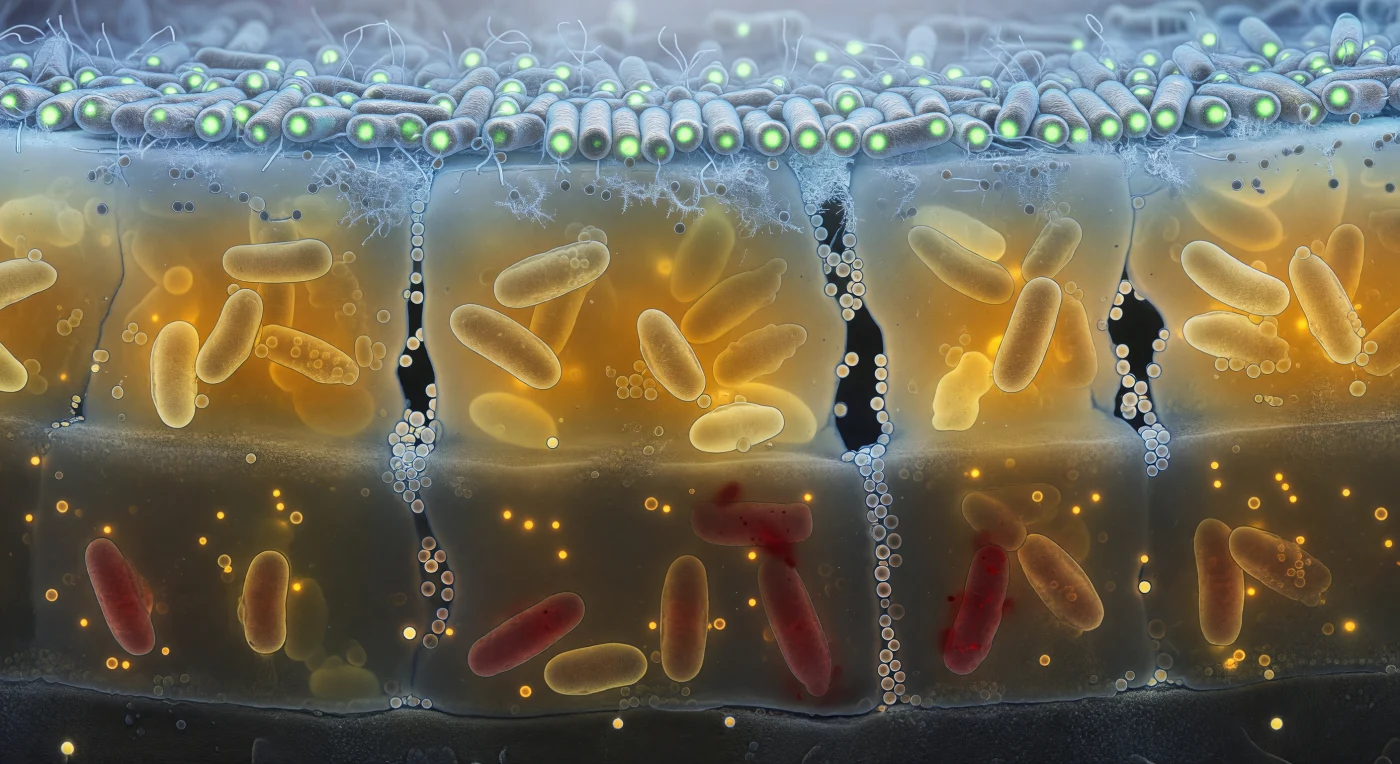
Je bevindt je in het binnenste van een levende biofilm, alsof je voor een levende rotswand staat die van binnenuit gloeit: boven je strekt zich een dicht baldakijn van GFP-groene staafcellen uit, strak opeengepakt als kolommen in een kathedraal, badend in het blauw-witte zuurstoflicht dat vanuit de waterige buitenwereld naar beneden sijpelt door de dunne lipidemembranen heen. Halverwege verdicht de extracellulaire polymere matrix zich tot een amberkleurige hars die alles omhult — metabolisch gestresste, geelgroene cellen half ingebed in het gel, donkere waterkanalen die als verticale schachten door de laag snijden en bezaaid zijn met membraanvesikels die zweven als zeepbellen in stilstaande lucht. Dieper nog, waar de zuurstof is uitgeput, onthult het roodpaarse schijnsel van propidiumjodide een begraafplaats van aangetaste, vervaagde cellen die hun inhoud lekken in de omringende matrix, terwijl volmaakte nanobellen van afgeworpen bilayer-materiaal geestig oplichten in het halfduister. Heel deze gelaagde wereld rust op een donkergrijze minerale basis waaraan de eerste cellen zich onomkeerbaar hebben vastgehecht — een geologie niet van steen, maar van leven, neergelegd in lagen van amper vijftig micrometer dik.

Je bevindt je in het absolute duister van een STORM-reconstructie, omringd door een spaarzame maar nauwkeurige constellatie van lichtpunten die samen de binnenste architectuur van één delende bacteriecel vormen. Vóór je beschrijft een gebroken boog van warm goudgele Gaussische vlekken de FtsZ-ring — het eiwit dat als moleculaire scheidslijn de cel in tweeën begint te trekken — waarbij elke vlek een discrete oligomeerpatch vertegenwoordigt van slechts twintig nanometer breed, gescheiden door holtes van puur niets, zoals een zin waaruit letters zijn weggevallen maar waarvan de curve een volmaakte cirkel suggereert. Aan weerszijden spiraleren koele cyaanwitte punten in een gebroken helix langs de cellengte: het MreB-cytoskelet, de interne steigerconstructie die de staafvorm van de cel handhaaft en begeleidt bij celwandsynthese. Duizenden rood-oranje punten vormen het membraanoppervlak boven en onder je — zo dicht opeengepakt dat ze bijna samensmelten tot gloeiende bogen, en toch blijft elk punt een afzonderlijke, nauwkeurig gelokaliseerde fluofoorevent, een enkel foton teruggewonnen uit optisch ruis. De dieptekleurcodering verandert de ruimte in een thermisch verloop: moleculen vlakbij je gloeien warm barnsteen, terwijl die verder weg geleidelijk verschuiven naar koper, roestbruin en ten slotte een koud blauwviolet aan de poolgebieden van de cel, alsof je op de equator staat van een moleculaire planeet waarvan de sfeer besluit te splitsen.
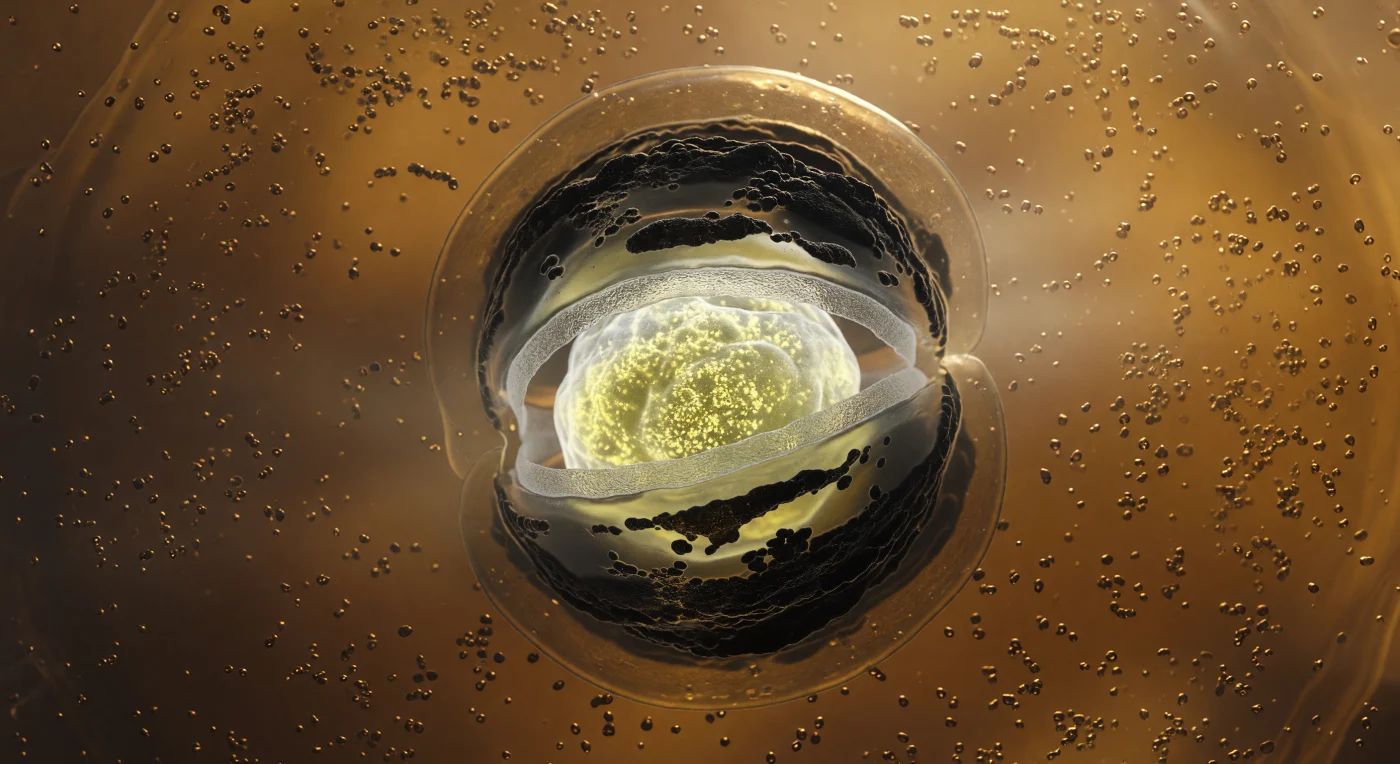
In het diepst van de moedercel kijk je naar buiten in een gouden, trilbare ruimte — een wereld van amberkleurig cytoplasma zo dicht verpakt met ribosomen dat het licht er doorheen filtert als door nat zand, alles trillend in de eeuwige Brownse beweging die op deze schaal zwaartekracht vervangt. Centraal in dit binnenste domineert de voorspore als een ingesloten wereld op zich: een ovale massa die van binnenuit gloeit met een koud, mineraalwit licht afkomstig van ultrageconcentreerde calcium-DPA-kristallen en SASP-gebonden DNA dat op bijna onmogelijke dichtheid is samengeperst. Om die kern heen liggen concentrische schillen van toenemende duisternis — eerst een doorschijnende ring van peptiglycaan-cortex die het innerlijke licht diffuus verspreidt als matglas, dan donkere, bijna matte lagen van SpoIVA- en CotC-manteleiwitten die licht absorberen in ruwe, onregelmatige bogen die doen denken aan afkoelend vulkanisch gesteente. Wat de blik definitief gevangeneemt is het omsluitende membraan zelf: een dubbelgelipide vel dat zich met extreme dramatiek rondom de voorspore kromt in een fagocytische omhelzing, verlicht vanuit de gloeiende kern in warm ambergoud en verduisterd aan de moedercelzijde tot koperbruin, terwijl het omringende cytoplasma geleidelijk wordt uitgesloten naarmate de insluiting zijn voltooiing nadert — het resterende volume vult zich met een vochtige, half-doorschijnende nevel van eiwitdamp die elke grens op enige afstand in zachte, biologische onscherpte doet oplossen.

Vanuit tachtig nanometer boven het oppervlak strekt de buitenmembraan van *E. coli* zich uit als een eindeloos, golvend landschap van lipopolysacchariden — een taaie, teel-gouden vlakte bezaaid met porine-trimeren die als vulkanische schoorstenen uit de biloog rijzen, voortdurend bevend in de thermische ruis van het omringende water. Vanuit het donkere aquatische medium daalt een T4-faag neer: een ijskoude, honderd nanometer brede icosaëdrische capsidekorrel wiens facetten in goud en donkergrijs de precieze geometrie van onderling vergrendelde capsomeereiwitten verraden, zes lange staartfibres wijd uitgespreid als de gelede poten van een kolossaal insect dat landt op onbekend terrein, elk contactpunt een zachte deuk drukkend in het LPS-bladsel. Het staartbuisje heeft de membraan al doorboord met een porie van nauwelijks drie nanometer, de omringende lipiden licht in wanorde gebracht, het periplasmatische compartiment dieper in donker teel te raden, terwijl een oplichtende, bleekblauwe draad van enkelvoudig-strengig DNA — onder osmotische druk vrijgelaten uit de capsideruimte — als een optische glasvezel door die naald stroomt en verdwijnt in de cel. Op de achtergrond zweven twee nog niet gecontracteerde fagenmoleculen als kille zilveren silhouetten in de blauwgrijze ternevel van het medium, hun staartscheden nog uitgestrekt, wachtend op het moment van eigen landing — een oeronbewuste maar onstuitbare kolonisatie, molecuul voor molecuul, in een wereld waar afstand wordt gemeten in bindingslengthen en tijd in microseconden.
