Confiança científica: Muito alto

O visitante encontra-se suspenso no interior de uma muralha viva, rodeado em todas as direções por uma malha densa de cadeias de glicano cor de âmbar queimado — longas hastes semicurvas que se entrelaçam numa geometria orgânica e compressiva, como pedra antiga comprimida ao longo de eras, atravessada por pontes peptídicas mais pálidas que fixam o conjunto numa rede tridimensional de densidade extraordinária. Através deste andaime dourado deslizam cortinas sinuosas de ácidos teicóicos em verde-floresta profundo, polímeros aniónicos que pendem entre os montantes de glicano sem se ligarem firmemente, conferindo à parede uma qualidade estratificada e levemente iridescente, como algas escuras suspensas numa corrente lenta. Lá em baixo, parcialmente velada pelas camadas da malha, a membrana interna ondula como uma lâmina fluida de ouro-âmbar aquecido, irradiando calor para cima através de toda a espessura da parede, com complexos proteicos emergindo como ilhas densas e mate na sua superfície luzidia. Acima, em direção à fronteira exterior esfarrapada, os filamentos de glicano tornam-se progressivamente mais finos e desordenados, dissolvendo-se em extremidades franjadas que cedem lugar a uma vastidão aquosa de azul pálido e frio — o meio extracelular — onde a luz difusa e sem origem aparente substitui o calor dourado do interior. Toda a cena transmite o peso imenso de uma estrutura que, nesta escala, funciona como uma fortaleza arquitectónica a suportar a pressão mecânica e osmótica de um mundo inteiro.
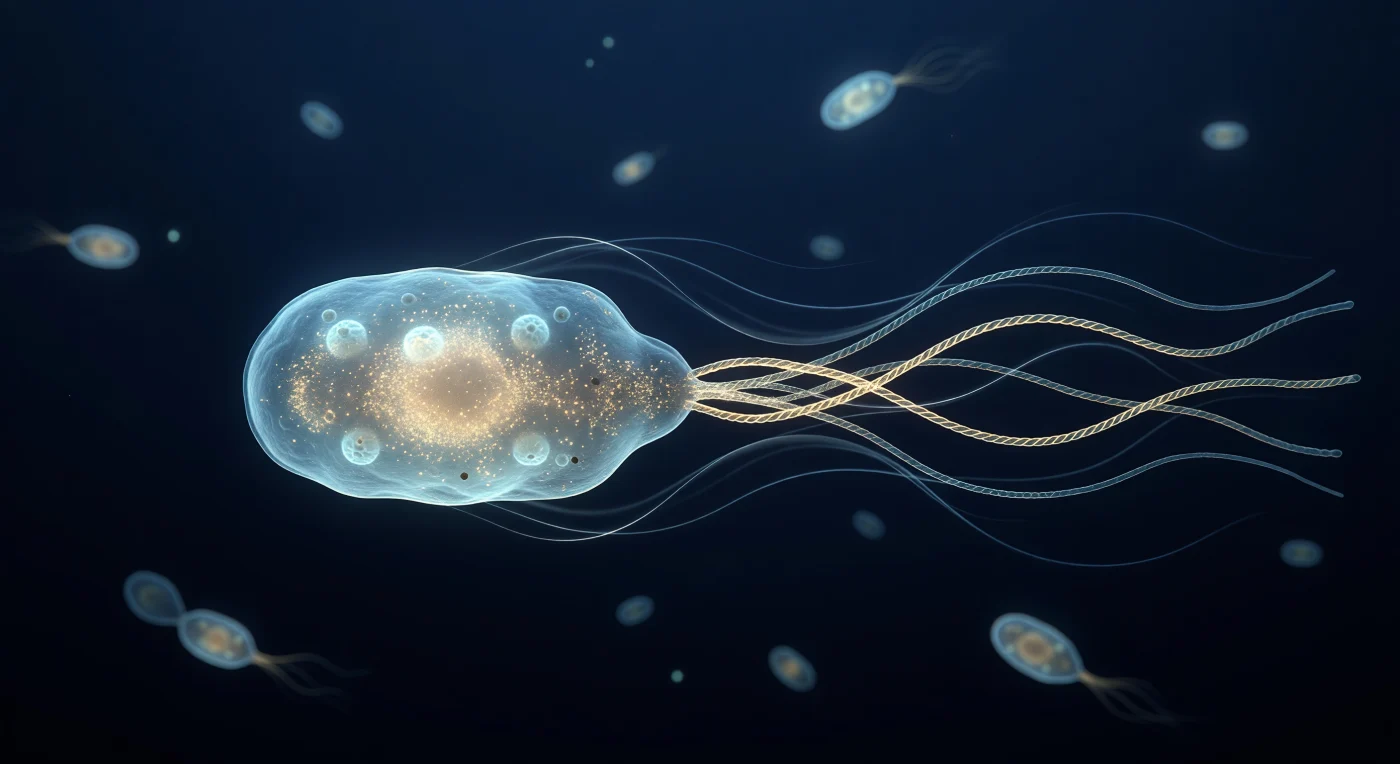
Suspenso imediatamente atrás de uma bactéria *Escherichia coli* em plena propulsão, o observador contempla um corpo celular translúcido de tonalidade ciano-pálida que ocupa o centro do campo visual como o casco de uma embarcação vista de uma chalupa à reboque, com a membrana externa cintilando numa orla prateada e fina que separa o interior âmbar — denso de ribossomas como névoa granulada — do oceano azul-marinho que o envolve. A popa da célula dá origem a quatro filamentos flagelares helicoidais de ouro-pálido que, ao entrançarem-se, formam um cabo superhelicoidal único a rodar no sentido anti-horário a mais de cem rotações por segundo, fundindo-se numa corda iridescente que se afunila para a escuridão com a elegância de um mecanismo molecular antigo. À volta desse feixe, o meio aquoso exibe linhas de fluxo de Stokes laminares — véus prateados e luminosos que se curvam para a frente em arcos perfeitamente simétricos, testemunho de um regime de baixíssimo número de Reynolds onde a viscosidade é soberana absoluta e qualquer impulso cessa no instante em que a rotação para. Ao fundo, uma dúzia de silhuetas bacterianas desfocadas deriva como brasas frias numa imensidão escura, os seus halos capsulares captando a luz ambiente em anéis translúcidos, enquanto o agitar térmico constante da água — cada molécula uma fonte de ruído cinético — confere ao espaço inteiro uma vibração silenciosa e pressurizada que nenhuma escala maior alguma vez conhecerá.

Suspenso no interior do nucleóide de uma célula de *E. coli*, o observador encontra-se envolto por uma nebulosa comprimida de DNA superenrolado em configuração plectonémica: cabos de dupla hélice de azul-marinho escuro entrelaçam-se em geometrias fractais que se ramificam em todas as direções, preenchendo cada linha de visão com uma densidade labiríntica que não deixa qualquer espaço vazio, enquanto uma luminescência fria análoga ao DAPI irradia das próprias fibras, banhando o ambiente num gradiente de índigo e azul-meia-noite. Ao longo de cada curva abrupta e ponto de cruzamento dos filamentos, pequenas proteínas associadas ao nucleóide — HU e Fis — agarram-se como nódulos amarelo-dourados de poucos nanómetros, introduzindo dobras mecânicas no DNA e devolvendo o brilho azul circundante como constelações de contactos âmbar dispersas pela nuvem. O meio citoplasmático entre as alças superenroladas, um gel viscoso com cerca de algumas dezenas de nanómetros de espessura, captura a luz índigo como um vidro aquamarino fumado, colapsando a perceção de profundidade numa série de planos recuados que se dissolvem em obscuridade azul-negra. Na periferia desta nuvem, onde o nucleóide se dilui e o citoplasma começa a impor a sua identidade, grânulos esféricos de cor castanha escura — ribossomas densos como brasas — agrupam-se em agregações soltas, ligeiramente desfocados pela refração do meio gelatinoso que os separa do interior da massa de DNA. A sensação total é a de flutuar dentro de uma tempestade pressurizada de matéria biológica organizada, onde a arquitetura é intrincada a cada escala de atenção e não existe silêncio, apenas a eloquência densa de uma máquina viva dobrada num volume menor do que o comprimento de onda da luz visível.

O observador paira a uma altitude infinitesimal acima da membrana externa de uma bactéria Gram-negativa viva, contemplando uma planície ondulante de lipopolissacarídeos que se estende até um horizonte suavemente curvado, como a superfície de um planeta minúsculo aquecido por baixo. O terreno oscila em vagas lentas e tectónicas, reflexo das flutuações térmicas da bicamada lipídica, enquanto as cadeias de oligossacarídeos se erguem verticalmente no meio aquoso viscoso como algas num mar sem vento, irradiando uma luminescência fria de interferência azul-esverdeada. De vinte em vinte nanómetros, trimeros de porinas beta-barril emergem como torres de obsidiana com poços interiores que perfuram a membrana em direção ao espaço periplásmico abaixo, as suas superfícies proteicas captando a luz difusa em reflexos violeta e azul sobre os territórios lipídicos vizinhos. Entre essas torres, mosaicos cristalinos de Lípido A cintilam com cores de interferência de fase — cobalto profundo, dourado-esverdeado e aquamarino vivo — numa textura que oscila entre obsidiana polida e a quitina iridescente de um élitro de besouro, suspensa no limiar entre o estado líquido-cristalino e o sólido. À esquerda, o corpo basal flagelar ascende da membrana como uma estrutura industrial de discos proteicos sobrepostos com precisão cirúrgica, o seu gancho nascente curvando-se para o meio iónico acima, enquanto toda a cena é banhada por uma luz criógenica difusa que filtra para cima através da bicamada translúcida, conferindo ao mundo ondulante a quietude luminosa de uma baía glacial ao crepúsculo.

O olhar mergulha num mundo sem horizonte, sem vazio, sem respiro: em todas as direções, ribossomas 70S empilham-se como pedras de uma muralha ciclópica, esferas de cor umber escuro separadas apenas por frestas de cinco a dez nanómetros de meio âmbar viscoso, uma matriz tão saturada de macromoléculas que se aproxima de um gel denso em vez de solução aquosa, com concentrações proteicas da ordem dos trezentos miligramas por mililitro que abolem qualquer noção de espaço livre. Cadeias de polissomas atravessam o campo médio como colares de pérolas irregulares — seis ribossomas enfiados num filamento de mRNA pálido e cremoso que cede e se curva sob o seu próprio peso molecular, desaparecendo na penumbra castanha além de dois ou três comprimentos de ribossoma. À esquerda, o barril duplo de um chaperonino GroEL ergue-se com uma verticalidade quase arquitetónica, os seus anéis empilhados em ardósia escura com cristas superficiais subtis e uma cavidade central ligeiramente mais fria onde proteínas mal dobradas encontram refúgio e reconfiguram a sua estrutura — uma fábrica de resgate conformacional que funciona num ciclo ATP-dependente de aproximadamente dez segundos. Por todo o campo, lampejos esmeraldas de proteínas marcadas com GFP irrompem brevemente como brasas verdes antes de serem obliterados pelo próximo ribossoma que passa, lembrando que este universo denso e monócromo está em movimento constante, governado não pela gravidade mas pelo ruído térmico que tudo agita, separa e volta a colidir sem cessar.

O olhar paira a cerca de doze micrômetros acima do substrato, inclinado sobre uma metrópole viva que nunca foi construída — apenas crescida: torres em forma de cogumelo erguem-se vinte a quarenta micrômetros desde a base, suas superfícies cobertas por células bacilares individuais de *Pseudomonas* dispostas em rosetas densas, cada bastão com pouco mais de um micrômetro de comprimento, luminescente em verde-jade pela expressão de GFP, a fluorescência emanando de dentro para fora como brasa biológica. Entre as torres, canais de água escuros e anecóicos cortam a paisagem com precisão hidráulica, resultado do fluxo contínuo que remove produtos residuais e distribui nutrientes — arquitetura funcional esculpida pela seleção, não pelo design. O espaço intersticial é preenchido por uma matriz de polissacarídeos extracelulares âmbar e translúcida, levemente autofluorescente em laranja, um gel viscoso secretado coletivamente pelas células e que lhes confere resistência mecânica, proteção contra antibióticos e coesão social — a própria substância do biofilm como organismo distribuído. Nas camadas basais das torres mais maduras, onde o oxigênio já não penetra através da matriz densa, manchas de vermelho iodeto de propídio revelam células com membranas rompidas, mortas pela hipóxia criada pelo metabolismo de suas vizinhas acima — o custo invisível da vida em colônia, legível apenas na profundidade confocal. Todo o mundo tem a espessura de um fio de seda e a complexidade de uma cidade inteira.
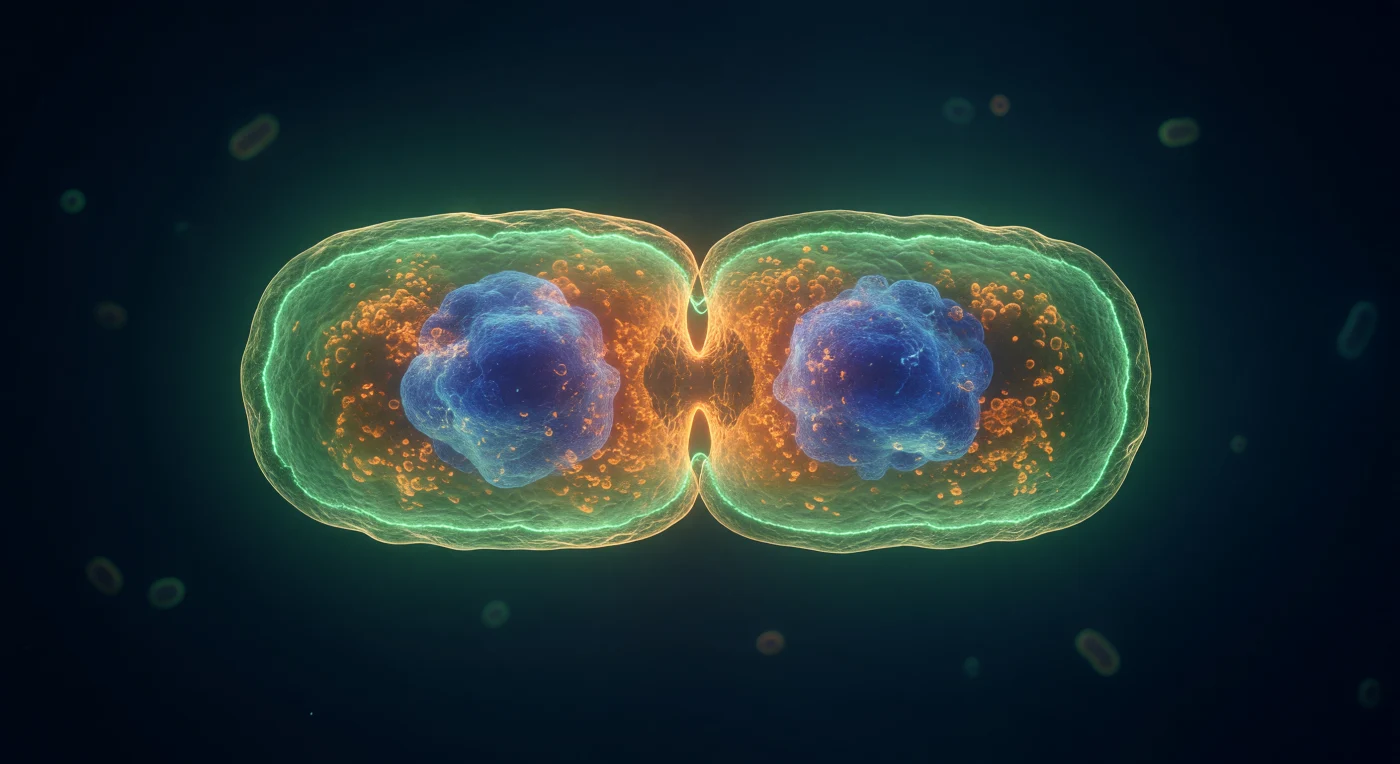
Suspendemo-nos na cintura equatorial de uma célula de *E. coli* no auge da sua própria divisão, orbitando o estreitamento em ampulheta onde o anel FtsZ — uma banda contínua de filamentos de proteína GTPase polimerizados com apenas 200 nm de largura — arde em verde elétrico de GFP, circundando toda a circunferência como uma coroa de fogo molecular que coordena o aparelho contrátil da divisão bacteriana. A membrana interna corada com FM4-64 forma uma dupla curva laranja-âmbar que segue cada milímetro da constrição, enquanto a parede celular de peptidoglicano — uma fina carapaça laminada de ouro pálido — capta a luz oblíqua e revela a textura hexagonal das suas ligações cruzadas à escala de nanómetros. Olhando através do corpo celular translúcido, as duas massas nucleóides de DAPI-azul-cobalto já se segregaram completamente para cada futura célula-filha, brilhando como nebulosas compactas envoltas em citoplasma âmbar e denso — um gel a 300 mg/mL de proteínas, ribossomas e RNA onde a difusão por movimento browniano supera qualquer força gravitacional. O meio aquoso que nos rodeia é um azul-marinho quase negro, onde formas bacterianas distantes e desfocadas captam o sangramento do brilho verde do anel Z como silhuetas fantasmagóricas, lembrando-nos que este drama teatral e iluminado acontece num mundo microscópico de uma densidade e atividade inimagináveis.

Diante de você, a cadeia de magnetossomas estende-se como uma obra de arquitetura mineral impossível, quinze cristais cúbicos de magnetita negra suspensa em fila perfeita, cada um erguendo-se a cerca de quatro vezes a sua altura, suas arestas facetadas lançando reflexos azul-violeta pelo citoplasma denso e âmbar que os envolve. Cada cristal é abraçado por uma vesícula lipídica translúcida, uma membrana delicada que pulsa levemente com a agitação térmica, conferindo a cada corpo mineral escuro um halo suave de pérola luminescente — a fronteira entre o mineral e o biológico tornada visível. Abaixo da cadeia inteira, o filamento citoesquelético de MamK corre como um trilho cobalt-elétrico, suas fibras proteicas entrelaçadas mantendo toda a série de cristais alinhada com uma precisão que serve de bússola interna para a célula, orientando-a ao longo das linhas invisíveis do campo magnético terrestre. O meio circundante é um gel opaco e quente, salpicado por ribossomas como pedregulhos escuros numa névoa âmbar-ocre, a visibilidade reduzida à densidade de uma matéria viva em constante agitação browniana. Ao longe, a membrana interna curva-se como o casco de uma caverna abobadada, além da qual apenas escuridão aquosa aguarda — o exterior indiferente de um mundo dominado pela viscosidade, onde nadar é empurrar o xarope e parar é cessar de existir.
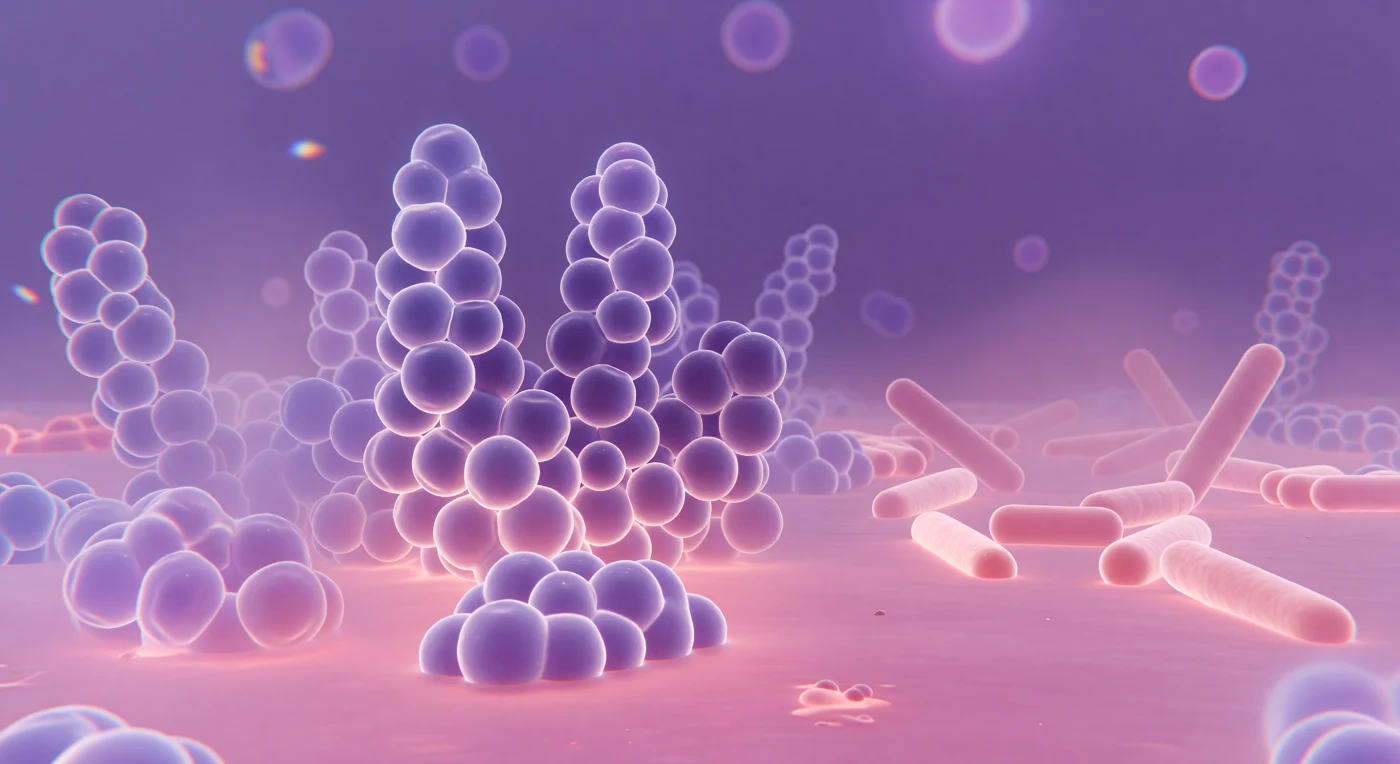
Diante de nós ergue-se um mundo saturado de violeta profundo e rosa quente, onde esferas colossais de *Staphylococcus aureus* — cada uma com o dobro da nossa altura — se agrupam em cachos compactos, suas paredes densamente impregnadas de violeta cristal absorvido pelas espessas camadas de peptidoglicano da parede celular Gram-positiva. Em torno do equador de cada esfera treme uma coroa de difração pálida, um halo iridescente que revela o momento em que a luz visível, cujo comprimento de onda se aproxima das próprias dimensões bacterianas, se dobra e se dispersa ao contornar formas que mal a ultrapassam em tamanho. O chão translúcido banha-se em safranina cor-de-rosa, um lago raso de contrastante que se acumula entre os clusters como vidro rosado, enquanto à direita bastone alongados de *E. coli* jazem espalhados numa planície aberta, seus cilindros pálidos absorvendo muito menos corante e irradiando um rosa suave em contraste com a fortaleza violeta que os domina. A profundidade dissolve-se rapidamente neste universo de foco raso: células ligeiramente acima ou abaixo do plano luminoso desvanecem em fantasmas lavanda, e franjas cromáticas de azul e âmbar coroam as formas periféricas, a imperfeição honesta da óptica de vidro que lembra que este mundo se estende em camadas infinitas de púrpura e rosa além desta única janela luminosa.
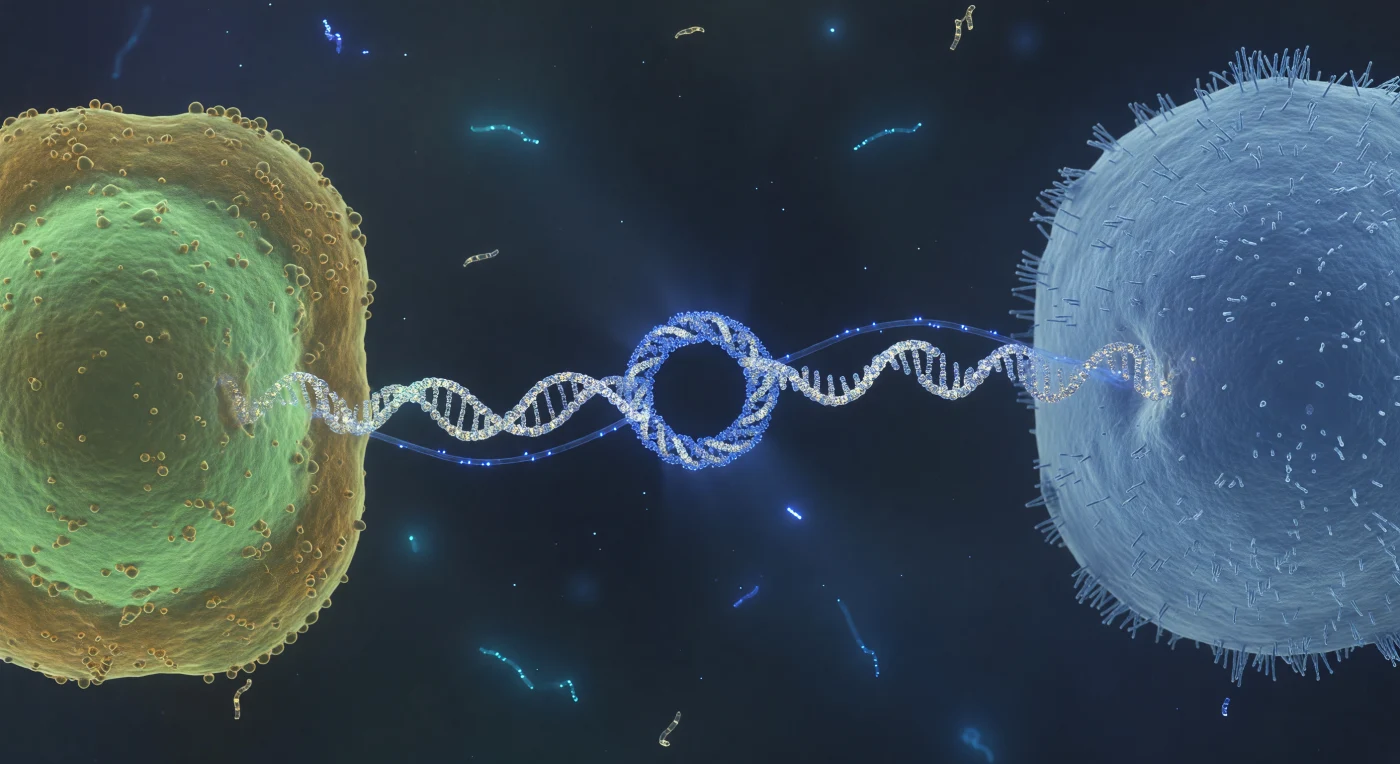
Pairando no ponto médio desta ponte molecular, o observador encontra-se suspenso entre dois mundos celulares separados por uma distância que, nesta escala, equivale a oceanos: um cabo proteico de oito nanómetros de largura — o pilus conjugativo — estende-se em arquitectura helicoidal entre a célula dadora, banhada num suave fulgor celadon-verde de fluorescência GFP, e a célula receptora, um monólito matte azul-pálido eriçado de fímbrias proteicas como uma geada densa. No ponto de contacto directo entre as duas membranas, o poro conjugativo resolve-se como um anel de ancoragem de vinte nanómetros, uma escuridão circular perfeita orlada por um halo de densidade proteica que capta a luz ambiente em aço azulado — a maquinaria molecular que autoriza a passagem de informação genética de uma linhagem para outra. Através do canal do pilus, um filamento de DNA de cadeia simples progride como uma fibra óptica viva, azul eléctrico e luminescente, difundindo halos azuis transitórios para o meio aquoso circundante numa pulsação que se sente mais como onda óptica do que como objecto discreto. O meio envolvente não é vazio mas sim um gel aquoso subtilmente leitoso, saturado de iões e detritos macromoleculares, onde o tremor térmico de tudo regista como uma imperceptível oscilação nas margens de cada estrutura, e onde, no fundo azul-negro do fluido em volume, fragmentos fantasma de DNA livre derivam como névoas ciano sob o bombardeamento browniano implacável.
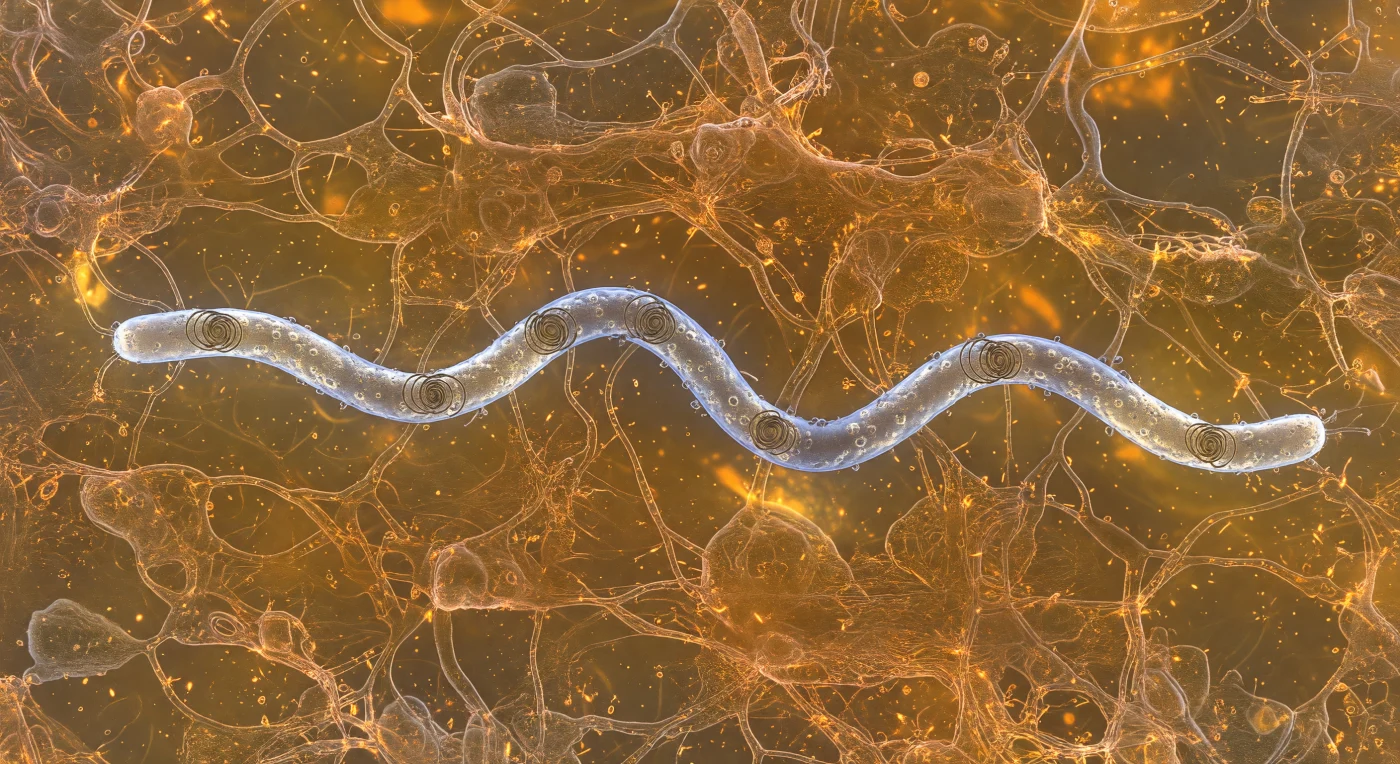
Suspenso na espessura âmbar-alaranjada do gel de mucina, o observador partilha o espaço imediato com uma *Borrelia burgdorferi* que avança em espiral a apenas cinco micrómetros de distância — um filamento helicoidal de prata fria, com dezoito micrómetros de comprimento, cujo movimento ondulatório de onda plana fende a malha de glicoproteínas como uma proa cortando seda molhada. A propulsão não provém de flagelos externos, mas de cabos helicoidais periplásmicos confinados sob a membrana exterior, que dobram toda a célula numa onda viajante contínua — uma solução evolutiva rara para navegar meios viscoelásticos de elevada resistência, como o muco intestinal ou sinovial que este organismo patogénico coloniza no decurso da doença de Lyme. A senda aberta pela bactéria reorganiza temporariamente os polímeros de mucina num rastro em V, cujos filamentos se curvam para fora e regressam à sua posição como um tecido de cobre em câmara lenta, enquanto vesículas da membrana exterior libertam halos refrátivos de material difundido na matriz circundante. Neste regime de número de Reynolds ultrabaixo — da ordem de 10⁻⁴ —, a inércia é inexistente e o fluido viscoso travar-se-ia instantaneamente se o ondulação cessasse, tornando cada milissegundo de movimento uma conquista contínua contra a resistência do meio. A nitidez cristalina dos filamentos em primeiro plano dissolve-se gradualmente numa névoa dourada e densa ao fundo, revelando a profundidade biológica de um ambiente em que cada volume cúbico de micrómetro encerra milhões de interações moleculares simultâneas.

O observador encontra-se suspenso no interior de uma *Synechocystis* viva, rodeado por camadas concêntricas de membranas tilacoides que se estendem em todas as direções como naves de uma catedral submersa, cada superfície irradiando uma luminescência carmesim profunda proveniente das centenas de complexos de clorofila embutidos na bicamada lipídica. Sobre essas membranas, os ficobilissomas hemisféricos — de um coral alaranjado e amarelo-açafrão intensos — cobrem cada face disponível como ornamentos barrocos, dispersando a luz vermelha em halos âmbar que preenchem os corredores luminais com uma névoa dourada e translúcida, quente e viscosa como resina em suspensão. Flutuando no citoplasma além da pilha de membranas, carboxissomos icosaédricos de cerca de 150 nanómetros de diâmetro derivam como lanternas de marfim fosco, as suas faces poliedrais captando o brilho vermelho das membranas num lado e projetando sombras azul-acinzentadas no outro, enquanto o interior leitoso e difuso denuncia os aglomerados de RuBisCO concentrados no seu interior. Todo o meio citoplasmático é um gel âmbar granuloso e densamente compactado — ribossomas demais numerosos para serem resolvidos individualmente constituem uma textura de ouro fosco que impregna cada interstício entre as estruturas. À distância, a fronteira curva da célula deixa transpirar uma neblina azul-esverdeada de exterior aquático, lembrando que este universo auto-iluminado e estratificado cabe inteiramente numa esfera com menos de três micrómetros de diâmetro.
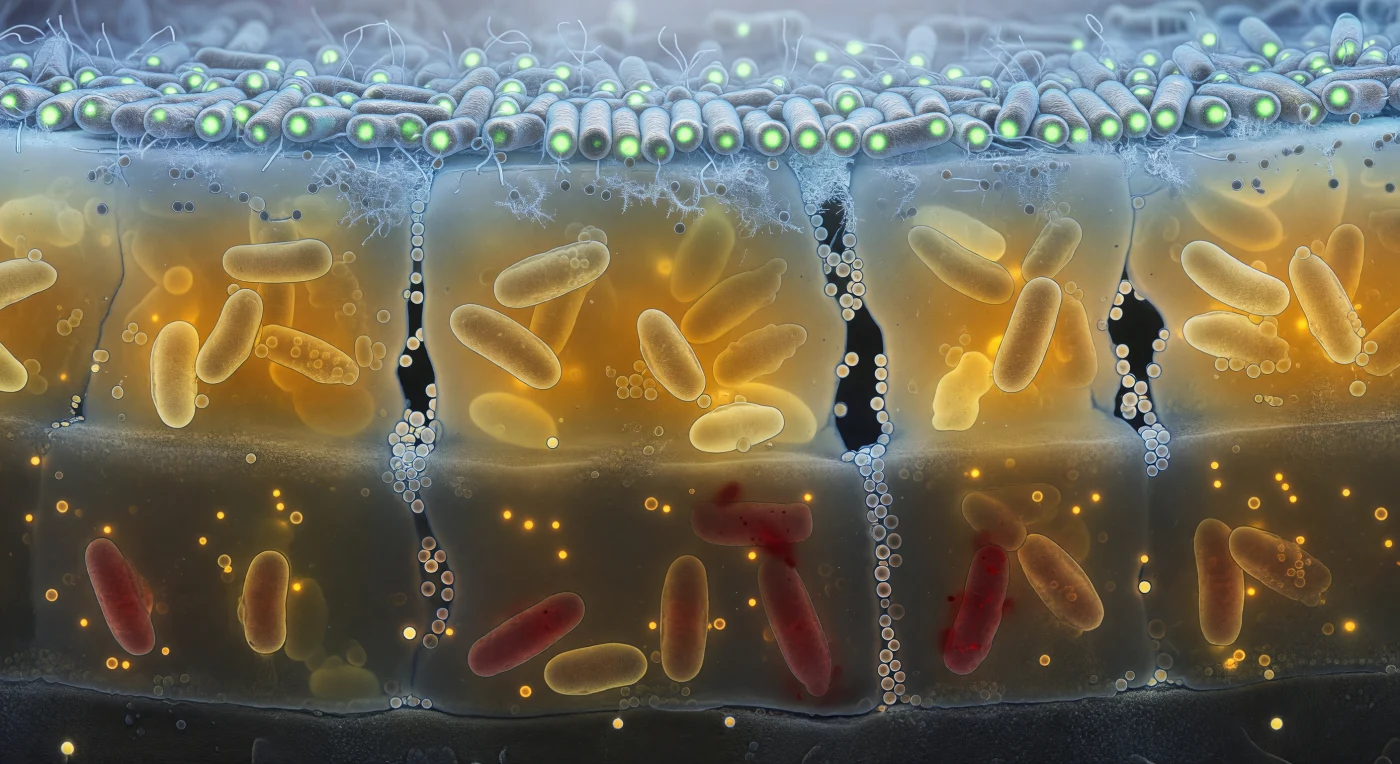
O observador encontra-se imerso na face vertical de um biofilme vivo, revelado como uma sequência de estratos geológicos empilhados em cinquenta micrómetros de espessura: a camada superficial resplandece em verde-frio de fluorescência GFP, com bacilos compactos e ordenados banhados por uma névoa azul-luminosa que marca a frente de oxigenação, onde as membranas lipídicas captam a luz como cilindros de vidro molhado. Descendo para a zona intermédia, a substância polimérica extracelular instala-se como uma resina âmbar translúcida que envolve células metabólicamente stressadas e canais de água escuros — vazios irregulares cujas paredes cintilam com vesículas membranares suspensas como bolhas de sabão em suspensão — enquanto o gradiente de oxigénio se apaga progressivamente numa penumbra castanha. No fundo hipóxico, apenas o vermelho baço do iodeto de propídio delimita células mortas e deformadas, as suas membranas permeabilizadas difundindo os seus conteúdos na matriz circundante num halo alaranjado, enquanto vesículas esféricas perfeitamente formadas flutuam imóveis no meio viscoso como bioluminescência de alto mar. A laje de substrato anceia tudo na base: uma superfície mineral cinzenta e quase sem traço, com as primeiras células bacterianas fundidas a ela por adesão irreversível, lembrando que nesta escala a gravidade é irrelevante e são as forças de superfície — de Van der Waals, electrostáticas, hidrofóbicas — que governam toda a arquitetura desta civilização estratificada.

No interior absoluto de uma reconstrução STORM, o observador flutua dentro de uma constelação tridimensional de pontos luminosos suspensos sobre um vazio negro total — cada ponto uma localização individual de fluoróforo, um evento fotónico recuperado do ruído óptico com precisão inferior a 20 nanómetros. Um arco incompleto de manchas amarelo-douradas traça o anel de FtsZ na cintura equatorial da célula, revelando como esta proteína de divisão não forma um círculo contínuo mas antes uma sequência de oligómeros discretos separados por lacunas de escuridão pura — a geometria da decisão de se dividir, escrita em luz esparsa. Ladeando o campo visual, dois trilhos helicoidais de pontos ciano-frio delineiam o citoesqueleto de MreB que percorre o comprimento celular, enquanto milhares de pontos vermelho-laranja definem a membrana com uma densidade tão elevada que quase formam arcos contínuos de brasa. A codificação axial de profundidade transforma o espaço num gradiente térmico — as moléculas mais próximas ardem em âmbar quente e as mais distantes esfriam progressivamente até um violeta gelado — fazendo com que o observador habite o interior de um planeta molecular vivo cujos polos estão perpetuamente congelados.
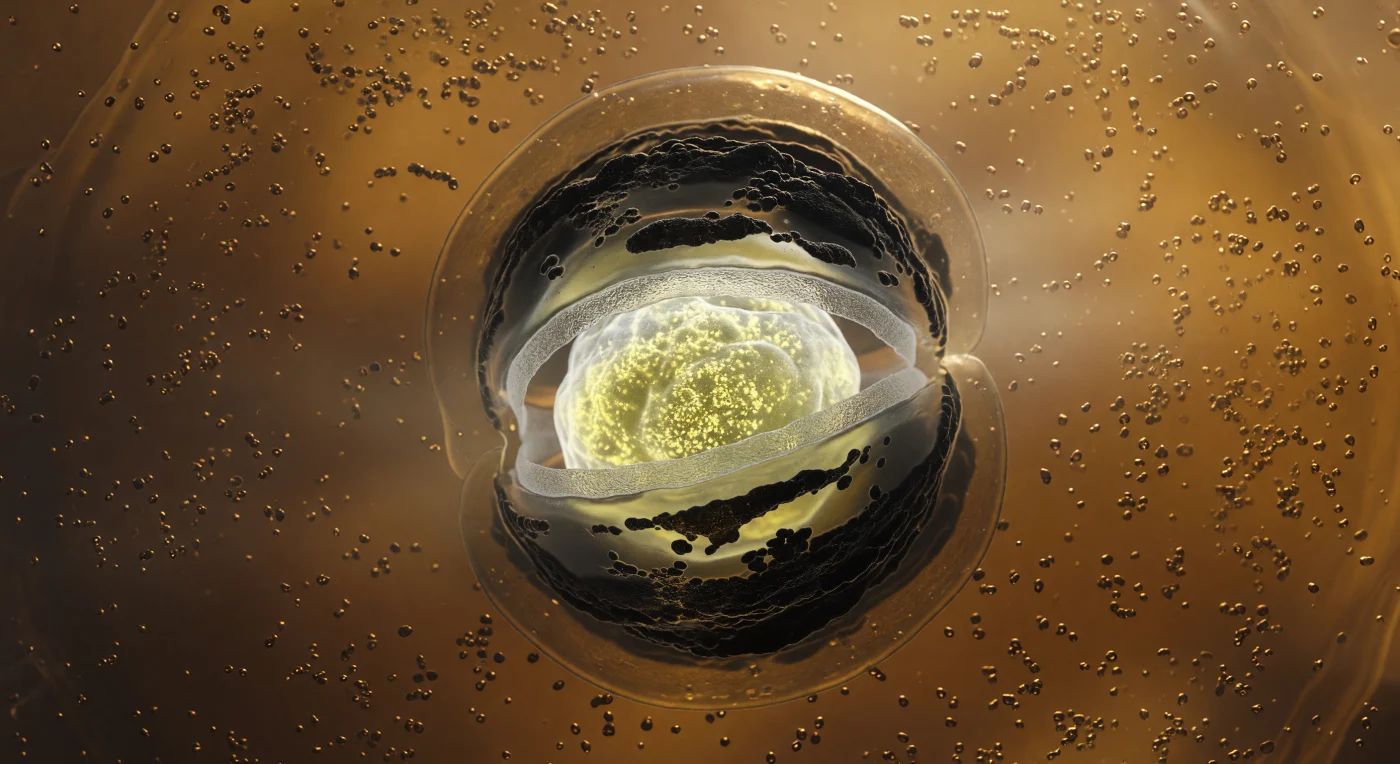
O observador encontra-se suspenso no interior de uma célula-mãe de *Bacillus subtilis* em esporulação, envolto numa caverna âmbar de citoplasma viscoso densamente repleto de ribossomas que captam a luz tênue como grãos de areia molhada — neste regime de número de Reynolds ultrabaixo, nada se move com leveza, e o meio resiste como mel morno a qualquer deslocamento. Ao centro desta caverna biológica, o forosporo domina o campo visual como um mundo dentro de um mundo: uma massa ovoide iluminada de dentro por uma luminescência amarelo-branca fria e mineral, emanada dos depósitos ultradensos de cálcio-DPA e do DNA condensado compactado para além de qualquer densidade biológica normal. Em torno desse núcleo resplandecente, um anel de cortex de peptidoglicano dispersa a luz como vidro fosco, enquanto camadas concêntricas de proteínas de coat — SpoIVA e CotC — polimerizam em arcos quase negros e opacos, absorvendo a luz como rocha vulcânica em arrefecimento. O que verdadeiramente prende o olhar é a membrana de engolfamento: uma dupla folha lipídica que se curva com extrema dramatismo em redor do equador do forosporo, formando um abraço fagocítico cujas superfícies iridescentes separa apenas um estreito espaço periplásmico, iluminadas do lado interior em âmbar dourado e mergulhadas em sombra cobreada do lado da célula-mãe. O citoplasma remanescente — granuloso, metabolicamente rico — vai sendo progressivamente excluído à medida que o engolfamento se aproxima da conclusão, e uma névoa proteica semiopaca suaviza cada contorno para além de algumas centenas de nanómetros numa dissolução suave e irremediavelmente biológica.

O observador paira a apenas oitenta nanómetros acima da membrana externa de uma célula de *Escherichia coli*, uma planície interminável de lipopolissacarídeos que se estende até todos os horizontes em tonalidades de azul-petróleo e ouro oxidado, pontuada por complexos proteicos que emergem como torres escuras e ventiladores vulcânicos — porinas rodeadas por um tênue brilho eletrostático âmbar onde a dupla camada iónica oscila em equilíbrio instável. Dominando o enquadramento superior, o capsídeo icosaédrico do fago T4 desce em silêncio como um corpo celeste facetado de cem nanómetros de diâmetro, as suas faces cinzento-escuro delimitadas por finas linhas douradas onde os capsómeros se encaixam em geometria precisa, seis longas fibras caudais espalhadas radialmente até à superfície lipídica como patas de um inseto ancestral gigante, cada almofada de ligação ao recetor imprimindo uma suave deformação local na folha teal-dourada. Ao centro da composição, o tubo de cauda já perfurou a membrana, criando um poro de escassos nanómetros cujo anel lipídico desordenado irradia um padrão de tensão mecânica, e através dessa agulha flui um filamento luminescente de DNA de cadeia simples — azul-branco translúcido, opulescente como fibra ótica na escuridão — em cascata contínua para o espaço periplásmico abaixo, impulsionado pela pressão osmótica interna do virião. Ao fundo, dois fagos adicionais pairam no meio aquoso azul-acinzentado com as suas bainhas caudais ainda não contraídas, reluzindo em prata fria, enquanto o próprio meio hídrico se manifesta como uma atmosfera volumétrica viva de flutuação térmica que suaviza contornos distantes e preenche o espaço negativo com a presença química saturada e intimidade biológica extrema deste mundo invisível.
