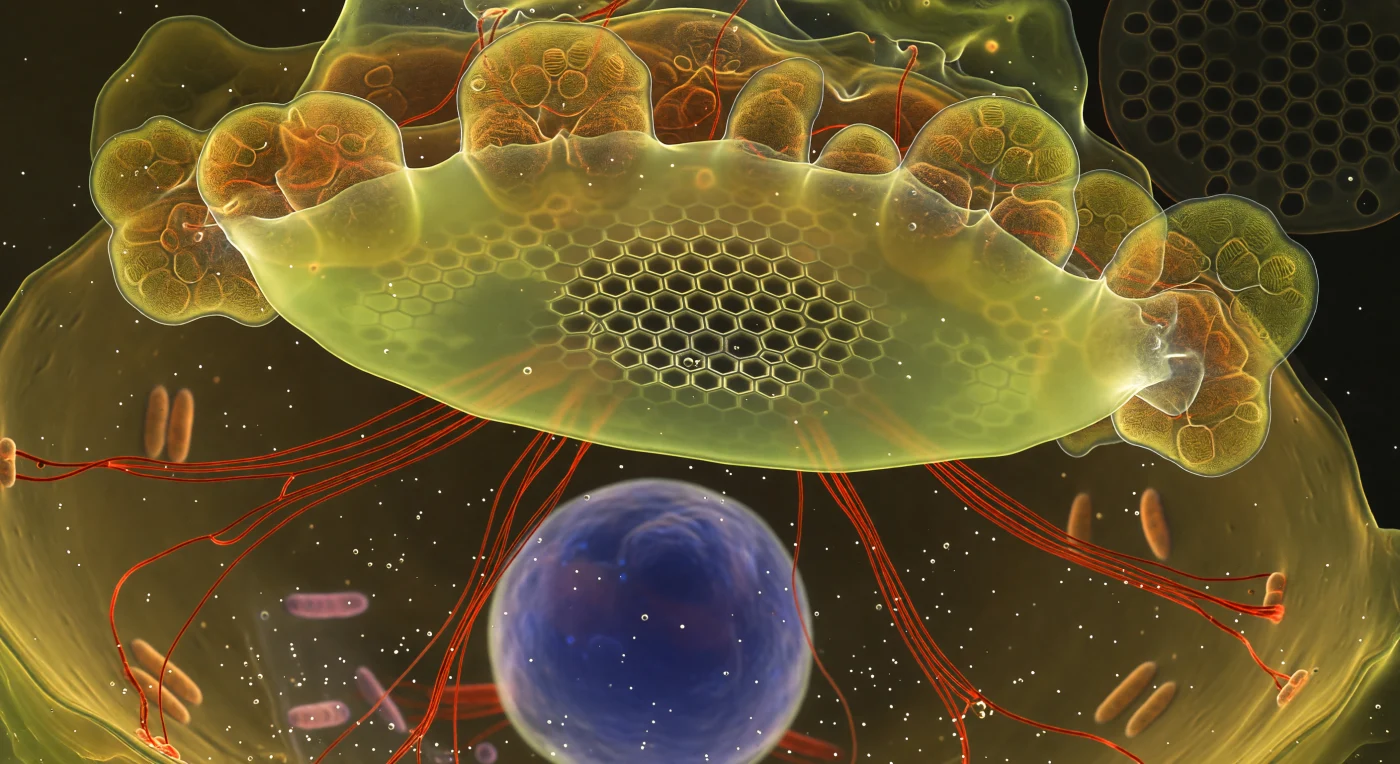
Im Inneren einer sich gerade teilenden Diatomee vollzieht sich ein stiller Gewaltakt der Mineralisierung: Die Silica Deposition Vesicle spannt sich wie eine leuchtende Membran quer durch das gesamte Sichtfeld, ihr gelbgrünes Chartreuse-Leuchten – das Ergebnis des in frisches Silikat eingelagerten PDMPO-Farbstoffs – erhellt den dichten cytoplasmischen Raum wie ein selbstleuchtende Decke aus flüssigem Glas. Im Zentrum dieser Struktur hat sich die fertige Wabenarchitektur bereits zu kristalliner Schärfe verfestigt: hexagonale Areolae, deren hauchdünne Silikatwände in weißgoldenem Licht erglühen, als trügen sie Hitze in sich, während die Geometrie nach außen hin in einen Schimmer noch unentschiedener, halb mineralisierter Geisterformen zerfließt. Strahlförmig verlaufende, intensiv karmesinrote Aktinkabel – durch Phalloidin sichtbar gemacht – schneiden wie glühende Drähte durch das warme Bernsteinlicht des Cytoplasmas und lenken mit molekularer Präzision die Form der entstehenden Valve. Goldbraune Chloroplastenlappen pressen sich an die siliziumgerahmte Decke, während tief im cytoplasmischen Dunkel darunter der Zellkern wie ein mattblauer Mond schwebt – ein fremder, stiller Beobachter in einem Kosmos, der sich in diesem Augenblick von innen heraus neu erschafft.
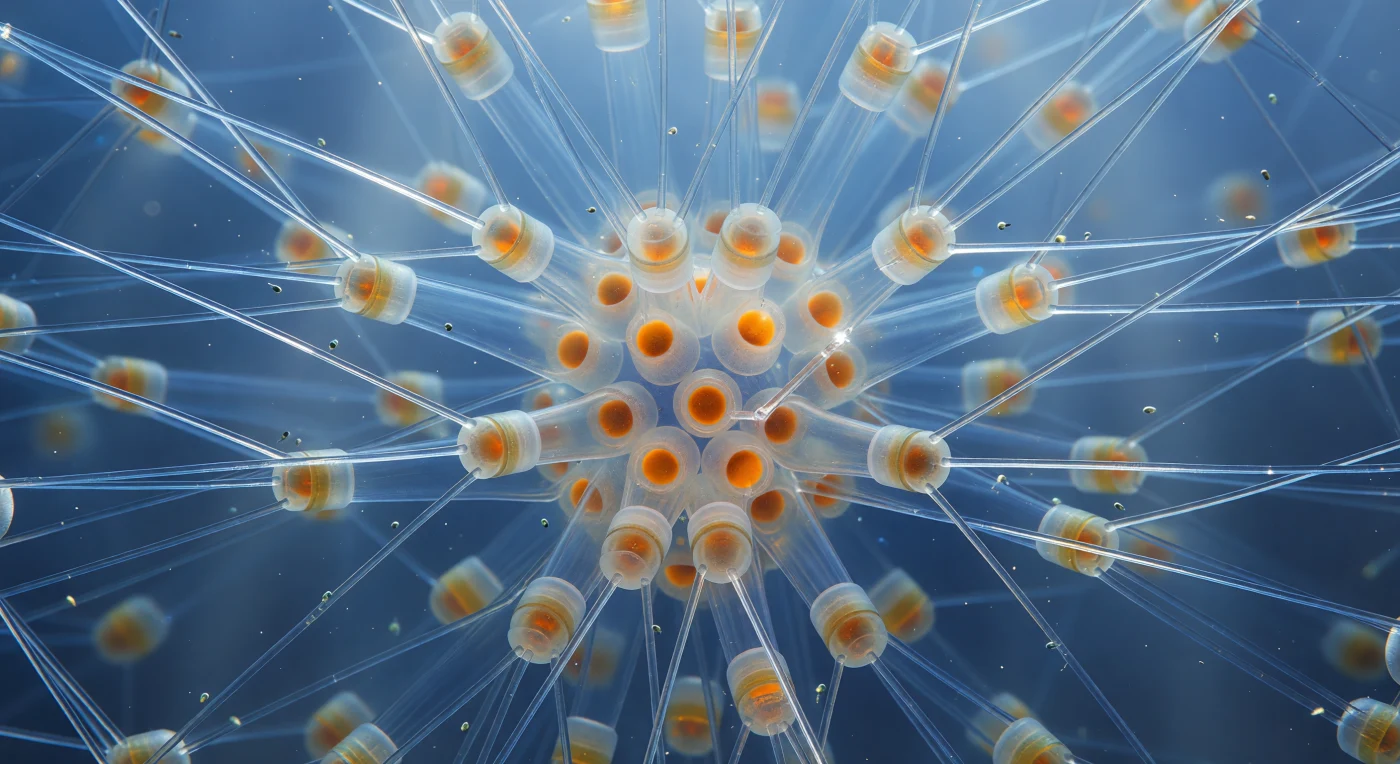
Der Blick führt in das geometrische Herz einer *Chaetoceros*-Kolonie, irgendwo im offenen Ozean, wo das Sonnenlicht noch als lange, kalte blaue Säulen von oben herabfällt — und in jede Richtung strecken sich hohle Kieselsäure-Setae aus, fast durchsichtige Glasnadeln von bis zu 200 Mikrometern Länge, die sich in verschränkten X-Gittern zwischen den gestapelten Zellzylindern kreuzen und an ihren gekrümmten Wandungen das Licht zu weißgoldenen Kaustik-Fäden bündeln, die wie helle Haarlinien gegen das tiefe Ozeanblau aufleuchten. Was von außen betrachtet unsichtbar klein wäre, entfaltet sich hier als architektonisches Innenleben: ein Kathedralenwald aus biogenem Glas, dessen Streben sich in jede Richtung ins Blaugraue verlieren, Schicht um Schicht, bis das Kobalt zu kaltem Indigo versinkt. Die zylindrischen Zellkörper — Frusteln aus amorphem, hydratisiertem Siliziumdioxid — leuchten mit dem warmen Bernsteinton fucoxanthinreicher Chloroplasten, während im dichten Koloniezentrum die schwer verkieselten Ruhesporen wie glühende Kohlen gegen das kühle Glasgerippe abstrahlen. Zwischen den Nadeln treiben nanometergroße Flagellaten auf unsichtbaren Strömungen — träge Staubkörner in einem lebenden Lüster aus Meereseis und Licht.

Der Blick fällt hinab auf eine kreisrunde Ebene aus lebendigem Glas, die sich in alle Richtungen bis an den Rand des Sichtfelds erstreckt – eine Coscinodiscus-Valve von 150 Mikrometern Durchmesser, die im Durchlicht wie ein liegendes Kirchenfenster leuchtet. Zehntausende hexagonaler Areolen aus amorphem, hydratisiertem Silikat – SiO₂·nH₂O, also biologisch abgeschiedenes Opalgas – fügen sich lückenlos zu konzentrischen Ringen zusammen und wirken dabei wie Linsen, die das von unten einfallende Licht zu warmen Bernstein-Punkten bündeln, während die Wandstärke zum Rand hin abnimmt und dort durch Dünnfilm-Interferenz in ein tiefes Blauviolett umschlägt. Aus dieser Glasebene erheben sich in gleichmäßigen Abständen die Fultoportulae – hohle Zylinder aus Silikat, deren offene Kanäle senkrecht nach oben zeigen wie Fernrohre, die mal goldenes Innenleuchten zeigen, mal kaltes blaues Streiflicht, und deren Schatten weich über je zwei oder drei benachbarte Areolen fallen und dem Ganzen eine stille topographische Tiefe verleihen. Das umgebende Meerwasser besitzt jene feine Cyanfärbung gelöster organischer Stoffe, und der abgeschrägte Rand des Diskus sendet beim Abfallen ins offene Wasser einen letzten Aufblitz prismatischer Irisierung – als löse sich ein Kontinent aus Glas langsam in blaues Dämmerlicht auf.

Vor dir erstreckt sich ein schier endloser Boden aus poliertem Glas, so flach und klar, dass er das diffuse Bernstein-Blaue Licht von oben wie ein stilles Wasser zurückwirft – überzogen von einem kaum wahrnehmbaren Schleier aus Meerwasser, einem molekülsdünnen Glykoproteinfilm, der dem Glas einen schwach irisierenden Glanz verleiht. Über diese Fläche gleitet eine *Navicula*-Zelle mit der ruhigen Unausweichlichkeit eines Binnenschiffs: dreißig Mikrometer lang, bikonvex im Querschnitt, ihre Kieselsäurehülle – ein amorphes, opalartiges Silikatglas aus zwei ineinandergreifenden Schalenhälften, Epi- und Hypotheka – fängt das schiefe Phasenkontrastlicht entlang ihres Dorsalkamms in einem dünnen Silbersaum ein, der sanft in warmes Gold und schließlich in Bernsteinschatten übergeht. Durch die transluzenten Siliziumdioxidwände leuchten zwei große Chloroplasten wie bernsteinfarbene Laternen hinter Mattglas, ihr Fucoxanthin-Pigment taucht den Glassockel darunter in zwei langgestreckte Lichtlachen warmen Goldes; das Cytoplasma strömt in kaum merklicher Zirkulation um sie herum, sodass man die Bewegung eher als ein Atmen des Lichts wahrnimmt denn als sichtbares Fließen. Hinter der Zelle liegt ein blassgelbes, schwach lichtbrechendes Schleimband auf dem Glas – der Mucilage-Faden, der durch den Raphespalt extrudiert wird und den Antrieb dieser rätselhaften Gleitbewegung ermöglicht –, und darin haftet, unbewegt wie ein dunkles Komma auf heller Seide, ein einzelner Bakterienrod: zwei Mikrometer klein, gehalten von derselben klebrigen Chemie, die auch den Kurs der Diatome in diese gläserne Welt einschreibt.

Vor dir hängt eine Kette aus *Thalassiosira weissflogii*-Zellen wie eine Perlenschnur in das blaugrüne Wasser des sonnendurchfluteten Oberflächenozeans, jede kreisrunde Silicascheibe etwa hundert Mikrometer breit, ihre Ventilfläche ein präzises Mosaik aus hexagonalen Areolen, das das von oben einfallende Licht in schimmernde Interferenzfarben – blasses Grün, warmes Gold – bricht. Ein einziger Chitinfibril verbindet die Zellen von Fultoportula zu Fultoportula: fast unsichtbar, bis ein schräger Lichtstrahl ihn als silbrig-weißen Faden aufleuchten lässt und damit die gesamte mechanische Last dieser lebenden Kolonialarchitektur offenbart. Durch die bernsteinfarbene Transparenz jeder Valve zeichnen sich innen goldene Chloroplastenlappen wie die Blütenblätter einer Rosette ab, durchsetzt von brechenden Lipidtröpfchen, während Bakterien als winzige dunkle Flecken an den Silicaoberflächen haften. Unterhalb der letzten sichtbaren Zelle verliert sich die Kette im volumetrischen Blau des offenen Wassers, und ein vorbeitreibendes Marine-Snow-Aggregat – ein zartes Gespinst aus Schleim, Detritus und Frustulen-Bruchstücken – erinnert daran, dass diese filigrane Konstruktion aus biogenem Glas zugleich Fundament der ozeanischen Nahrungskette und künftiges Sediment in den Tiefen des Meeresbodens ist.

In absoluter Schwärze schwebt man im Zentrum eines riesigen Arrangements aus Kieselsäure-Architekturen, die sich wie ein erstarrtes Planetarium bis an die Grenzen der Wahrnehmung ausdehnen – jede Frustule ein leuchtendes Reliquiar aus biogenem Opal-A-Glas, das aus dem Nichts heraus kalt aufzustrahlen scheint. Unmittelbar vor einem dominiert ein *Triceratium* das Sichtfeld: ein gleichseitiges Dreieck aus gestuften Kieselsäurewällen, dessen hexagonale Areolen – jeweils kaum 200 bis 500 Nanometer im Durchmesser – als Beugungsgitter wirken und elektrisches Kobaltblau, geschmolzenes Gold und violette Interferenzfransen in die umgebende Leere werfen, während die kaum nanometerdicken Glaswände selbst im reinsten Kontrast zwischen Licht und Nichts oszillieren. Weiter außen rotiert ein *Coscinodiscus* wie eine Domkuppel aus konzentrisch angeordneten Fultoportulae-Schornsteinen in tiefem Zitronengelb, daneben strahlt ein *Arachnoidiscus* speichenartig weißes und zartrosa Licht in gleichmäßigen Fächerstrahlen aus, während ein *Pleurosigma* in Kurven wie ein Kalligraphenstriches in grünem Turmalin und Amethyst durch das Bildfeld gleitet – sein obliques Areolengitter macht jede Perspektivverschiebung zu einem Schauer aus Blaugrün und Violett. Diese säurebereinigten, organisch entkleideten Mineralskelette – fossile Überreste photosynthetischer Einzeller, die seit Millionen Jahren in Sedimenten überdauern – versammeln sich hier zu einer Konstellation aus gefrorenem Meereslicht, einem viktorianischen Kabinett kosmischer Edelsteine auf einem einzigen Fokalebene.

Direkt unter dir erstreckt sich eine weitläufige, leicht gewölbte Ebene aus biogenem Siliziumdioxid – eine einzige Zellvalva von *Pinnularia nobilis*, die unter dem schräg einfallenden Licht der Differentialinterferenzkontrast-Optik wie gehämmertes Elektrum aus Gold und Zinn schimmert. Parallele Transversalcostae – die Rippen des Frustulums – ziehen in nahezu militärischer Regelmäßigkeit quer über das gesamte Sichtfeld: niedrige Rücken aus poliertem Opalglas, deren Luvseiten in Knochenweiß aufleuchten, während ihre Leeseiten in kühles Blaugrau absinken, denn die amorphe Kieselsäure ist kein kristallines Mineral, sondern ein hydratisiertes, glasartig erstarrtes Biopolymer, das Licht durchlässt wie bernsteinfarbenes Opalglas. Längs der gesamten Hauptachse zieht das Sternum als helle, axiale Hochstraße – ein struktureller Kiel, der die Valve bilateral teilt –, und wo der Rapheschlitz das Sternum unterbricht, öffnet sich eine in molekularer Präzision gefräste Rinne von weniger als 200 Nanometern Breite, deren Schatten eine Tiefe suggeriert, die in keinem Verhältnis zur sichtbaren Oberfläche steht. Durch die transparente Silicawand hindurch leuchten zwei H-förmige Chloroplastenlappen in warmem Bernsteinorange – das Fucoxanthin-Pigment verleiht dem Licht eine glühende Tiefe –, während zwischen ihnen der Zellkern als milchig-opaleszendes Ellipsoid liegt, von innen beleuchtet wie eine mattierte Glaskugel, die ihr eigenes diffuses Licht zu erzeugen scheint.

Einen Zentimeter über der Wattoberfläche schwebt man über einem lebenden Teppich aus Bronze und Bernstein, der sich bis zum Horizont erstreckt: Millionen pennater Diatomeen haben sich in einem dichten Biofilm zusammengedrängt, ihre polierten Silikatfrusteln liegen Flanke an Flanke wie Blattgoldstreifen unter dem schräg einfallenden Morgenlicht, das die gesamte Fläche mit einer warmen, metallischen Luminosität erfüllt. Die Oberfläche ist keine tote Schicht, sondern lebt — ganze Populationen wandern koordiniert nach oben, verdichten das Mat zu sanften Rücken aus konzentriertem Glas und Pigment, die in sattem Amber glühen, während die Ränder in transparentere Zonen ausblenden, wo die Zellen dünner werden. Zwischen diesen Rücken zittern Tröpfchen zurückweichenden Meerwassers wie kleine Linsen, die den goldenen Teppich darunter leicht verzerrt vergrößern, bevor sie verdunsten oder im Seewind davongleiten. Tiefer im Blickfeld, dort wo das Sediment noch feucht ist und dünne Wasserfilme abfließen, wechselt die Farbe von hellem Gold über tiefes Olivgrün und Siena — der Biofilm, durch trübes Salzwasser gesehen, das organische Partikel trägt und das Licht in einen milchigen Schimmer streut. In der unmittelbaren Nähe kurven einzelne Gyrosigma-Zellen wie polierte Mahagonikanus durch das transparente extrazelluläre Polymergel, während Sandkörner wie Felsbrocken aus der Matrixragen, ihre Flanken dicht mit flach aufliegenden Cocconeis-Valven gekachelt, deren feine Striae das Licht in zarte Spektralfächer brechen — ein stilles Zeugnis dafür, dass die regelmäßige Nanostruktur der Silikatschalen als Beugungsgitter wirkt.

In einem schwerelosen Zwischenraum über der beleuchteten Epilimnion-Zone eines Süßwassersees schwebt vor dir eine *Asterionella formosa*-Kolonie: ein vollkommener achtarmiger Stern aus biogenem Silikat, der das Sichtfeld mit einer Geometrie füllt, die zugleich architektonisch und lebendig wirkt. Jeder der acht Arme ist ein schlanker Frustulstab, dessen transluzente Silika-Wandung das diffuse Tageslicht von oben in kalte Interferenzfarben bricht – dünne Goldsäume und kühles Silber entlang der Striae-Riefen auf jeder Ventilscheibe. Im Zentrum bindet ein dichtes Mukopolysaccharid-Haftpad alle Arme zu einer Rosette zusammen, harzbernsteinartig und dicht, während sich entlang jedes Stabs zwei goldbraune Chloroplasten erstrecken, deren Fucoxanthin-Pigmentierung durch das Glas wie von innen beleuchtet scheint. Das Medium ringsum ist keine abstrakte Flüssigkeit, sondern eine viskose, widerstrebende Präsenz – das tannin- und organikgetönte Wasser drückt gleichmäßig gegen jede Silika-Fläche, und die gesamte Kolonie dreht sich in einer so langsamen Rotation, dass Bewegung erst durch das Verschieben der Interferenzreflexe erkennbar wird. Im unscharfen Hintergrund treiben dreiarmige *Staurastrum*-Desmidiaceen wie jadegrüne Laternen, und cyanobakterielle Fäden ziehen sich als blaugrüner Rauch durch die Wassersäule – ein stilles Ökosystem aus Photosynthese, Silikat-Mineralisierung und biologischer Zeit.

Zwischen zwei aufgesplitterten Fragilariopsis-Schalen, die wie abgelegte Rüstungen seitlich davontreiben, beherrscht die Auxospore das Blickfeld – eine perlmuttschimmernde Kugel, deren Durchmesser die leeren Kieselgehäuse bereits um ein Vielfaches übertrifft und die sich mit stiller, unaufhaltsamer Kraft weiter ausdehnt. Die verlassenen Frusteln flankieren die Szene wie zerbrochenes Treibgut: aschgrau gerippt, ihre Gürtelbänder in konchoidalen Bruchkanten geöffnet, ihre Siliziumwände in stahlblauem und geisterhaft grünem Interferenzschimmer leuchtend – physikalische Farbigkeit ohne Pigment, entstanden allein durch die Dicke von Glas, das dünner ist als Licht. Die Perizoniumhülle der Auxospore trägt kaum aufgelöste Querrippen aus frisch abgelagertem Silizium, die im schrägen Durchlicht zu goldenen Bögen aufflackern und wieder verblassen, während innen ein dichtes Goldbraun aus gestapelten Chloroplasten glüht, durchsetzt von hochbrechenden Lipidtröpfchen, die wie kleine Sonnen im Bernstein stehen. Im geometrischen Zentrum öffnet sich eine große klare Vakuole wie eine Wasserlinse, die den gesamten lebenden Inhalt nach außen gegen die spannungsgeladene Membran presst – eine stille Expansion, angetrieben einzig von osmotischem Druck und dem mechanischen Gesetz wachsender Oberflächen.

Der Blick öffnet sich in den Innenraum eines schmalen Kanals, der durch das antarktische Meereis schneidet – die Wände steigen wie facettiertes Polarglas um den Betrachter auf, tiefblau leuchtend im Kern, aquamarinfarbig ausblassend, wo winzige Eiskristalleinschlüsse das von oben herabsickernde, stark gedämpfte Licht brechen und in Keile aus Cyan, Silber und blassem Violett zerspalten. Die hyperhaline Sole, die diesen Kanal füllt, ist leicht viskos und trägt einen bernsteinfarbenen Schimmer aus gelöstem organischem Material, der jedem Lichtstrahl eine schwere, prismatische Qualität verleiht. Durch diese Flüssigkeit driften Ketten von *Fragilariopsis cylindrus* – acht Mikrometer lange Siliziumdioxid-Zylinder, aneinandergereiht wie transparente Perlenschnüre, deren fucoxanthinreiche Chloroplasten wie zusammengepresste Glut in Bernsteingold hinter den gläsernen Frustulenwänden brennen, umhüllt von kaum sichtbaren antigefrierenden EPS-Gelscheiden, die sich wie Atemhauch auf kaltem Glas um jede Kette legen. Den Boden des Kanals bedeckt eine dichte, dunkelgoldbraune Lage abgesunkener Diatomeenbiomasse – eine lebende Matte aus abgelagerten Zellen in der Farbe alten Bernsteinharzes, durchzogen von einzelnen, noch photosynthetisch aktiven Chloroplasten, die wie Lichtfunken in der Tiefe des polaren Winters glühen.

Wer hier schwebt, sieht kein biologisches Objekt mehr, sondern eine Welt aus geschliffenem Glas — eine gewölbte, grau-weiße Ebene, die sich in alle Richtungen bis zum Horizont erstreckt und deren Oberfläche in Tausende sechseckiger Schächte aufgeteilt ist, jeder einzelne ein präzise geformter Brunnen aus amorphem Opalsilizium, zweihundert Nanometer tief, an seinem Grund überspannt von einem Velum-Maschennetz, das nur durch die schräg eintreffende Elektronenstrahlung überhaupt sichtbar wird. Diese Areolae sind keine Zufallsprodukte, sondern das Ergebnis einer streng genetisch kontrollierten Biomineralisation: Die Kieselsäure wird innerhalb spezialisierter Membranvesikel — den Silica Deposition Vesicles — innerhalb weniger Stunden aus gelöster Monosiliziumsäure polymerisiert und in hexagonal gepackte Kammermuster gezwungen, deren Präzision kein menschliches Fertigungsverfahren erreicht. Aus der Fläche erheben sich in unregelmäßigen Abständen die Fultoportulae, hohle Siliziumsäulen mit drei Satellitenporen an ihrer Basis, durch deren offene Schächte die Zelle Chitinfibrillen nach außen sekretiert — stumme Kamine über einem mineralischen Plateau. Am Rand der Valve, wo die flache Oberfläche scharf in den Mantel abknickt, öffnen sich die Rimoportulae als dunkle, schlitzförmige Durchbrüche im Siliziumglas, und darunter beginnen die glatten, ornamentlosen Cingulumbänder, die — wie die Ringe eines steinernen Mauerwerks — die beiden Schalenhälften der Frustule zusammenhalten und in die Tiefe des Blickfelds verschwinden.

Der Blick geht über eine Landschaft, die sich wie ein sanft gewölbter Kontinent aus Olivgrün und bernsteinfarbenem Amber erstreckt – die Oberfläche eines Kelpblattes, dessen pflanzliche Zellen wie ein Mosaik aus matten Glasscheiben unter dem gefilterten Meerwasser schimmern, während kaustische Lichtvorhänge im Sekundentakt darüber hinwegtreiben. Aus diesem Untergrund erheben sich Dutzende *Licmophora*-Kolonien auf schlanken Muzilago-Stielen: jede ein goldbraunes Fächer-Gespann aus zwanzig bis vierzig länglichen Zellen, deren fein gestreifte Silikatfrusteln warmes Bernsteinlicht durchlassen und bei flachem Einfall zarte Interferenzfarben – Türkis, Kupfer – über ihre Ventilflächen gleiten lassen, als wären es Pfauenfedern aus gefrorenem Licht. Zwischen ihnen liegen *Cocconeis*-Valven nahezu unsichtbar dem Kelp angeschmiegt, elliptische Silikatschilde, die sich kaum von der EPS-Matrix abheben, während *Rhabdonema*-Bänder in Zickzack-Spiralen durch das Mittelfeld fädeln, ihre Zellkanten als helle Silberlinien im blauen Wasser aufblitzen. Im Hintergrund zieht sich eine helle Schabnarbe durch den Biofilm – der frische Weg eines grasenden Amphipoden, dessen zerstörte Muzilago-Ränder noch leicht aufgewölbt sind und das Licht in einem weichen irisierenden Dunst brechen, während das Tier selbst wie ein Berg aus durchscheinendem Gelenkpanzer am Narbenrand hockt und mit seinen Mundwerkzeugen die letzte Reihe goldener Fächer in gleichmäßige Schwingungen versetzt.

Im Inneren dieser gewölbten Kalzitkammer schwebt der Blick durch einen Raum, der an das Mittelschiff einer romanischen Kathedrale erinnert – cremeweiße Wände aus feinkristallinem Kalzit, halbdurchscheinend und von gedämpftem Licht durchdrungen, das aus der Korallensandwelt jenseits der Kammerwand hereinfiltriert, ohne eine einzige klare Quelle zu besitzen. Die Wände sind nicht glatt, sondern mosaikartig aus winzigen Kalzitkristallen gefügt, und die runden Porenöffnungen im Gehäuse erscheinen als tiefschwarze Bögen, durch die feine, fadenhafte Pseudopodien in das umgebende Meerwasser greifen. Den Boden und die Mitte des Raums füllt ein blasses, gallertartiges Zytoplasma der Foraminifere Amphistegina, ein lebendiges Medium, das seine Nitzschia-Diatomeen wie Passagiere in einem geschützten Hafen trägt – jede Zelle ein schlankes, bootförmiges Kieselglasfragment von zwanzig bis vierzig Mikrometern Länge, an den Rändern mit einem kaum wahrnehmbaren Irisieren aus Interferenzlicht. Tief in ihrem Inneren leuchten die Chloroplasten aus bernsteinfarbenem Fucoxanthin wie altes Harz im Kerzenschein, reich und warm gegen das blassgraue Zytoplasma gesetzt, während winzige Lipidtröpfchen als helle, stark lichtbrechende Punkte zwischen den Farbstoffloben funkeln. Diese Kammer ist ein Ort produktiver Stille, in dem die Diatomeensymbionten – ruhiggestellt, aber photosynthetisch aktiv – in der Wärme ihres Wirtes gehalten werden, während gefilterte Helligkeit durch Kalzitwände sickert und alles in einem gleichmäßigen, perlmuttartigen Schimmer badet.
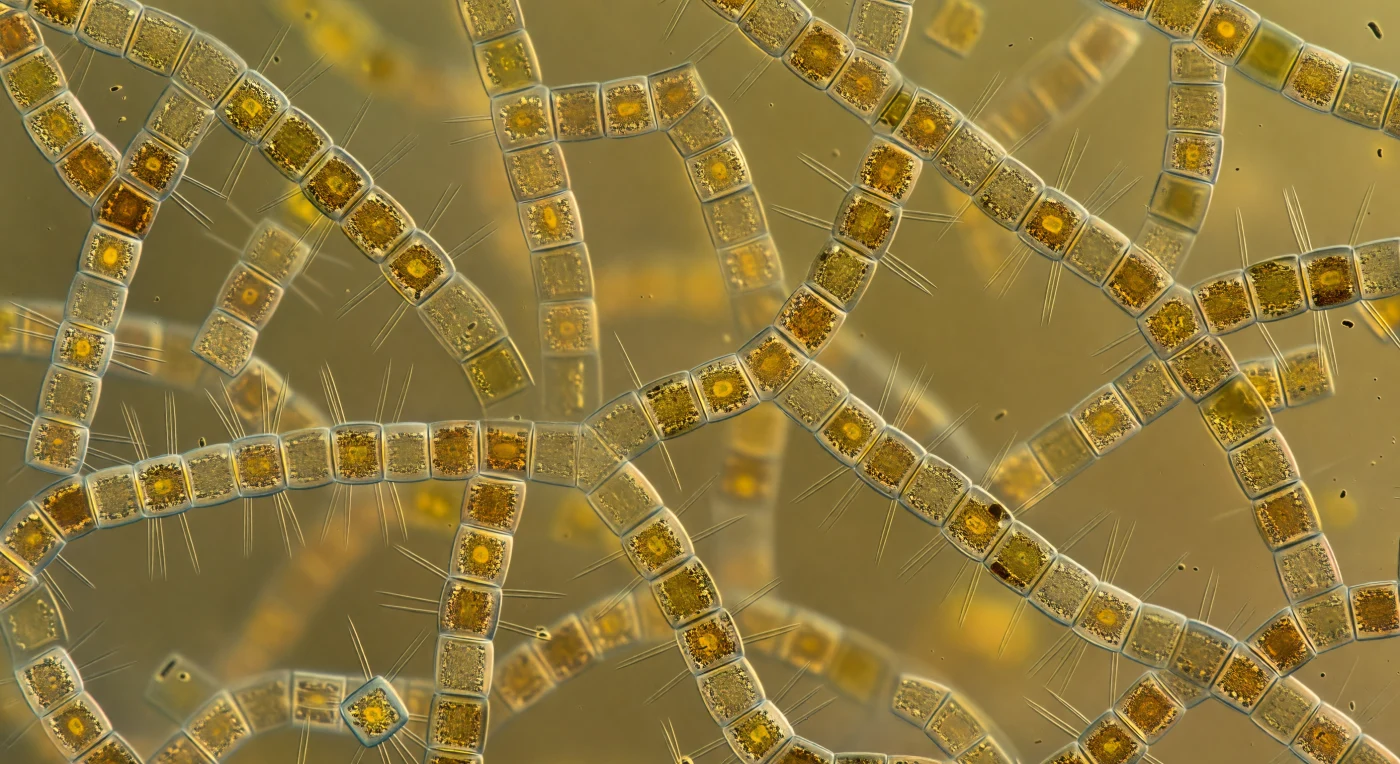
Der Blick löst sich in alle Richtungen gleichzeitig auf: Zylinder aus Kieselglas, fünf bis acht Mikrometer breit, reihen sich zu gewundenen Ketten, die sich verflechten, gabeln und in einen gleichmäßig leuchtenden Bernsteinnebel auflösen, ohne dass irgendwo ein freier Hintergrund sichtbar wäre. Jede Verbindungsstelle zwischen zwei Zellen ist ein aufgefächertes Bündel haarfeiner Fultoportula-Streben aus Siliziumdioxid, die wie die Rippen einer zersprengten Laterne in den Zwischenraum greifen und mit den Streben der Nachbarkette ineinandergreifen – ein dreidimensionales Gerüst aus Glas, das sich ohne Unterbrechung in jeden Raumwinkel erstreckt. Das Licht hat keine Quelle mehr: Sonnenstrahlen, die von oben als kühles Blauweiß einfielen, wurden von Millionen fucoxanthinhaltiger Chloroplasten in jedem dieser gläsernen Trommeln absorbiert, gestreut und neu abgestrahlt, bis das Medium selbst leuchtet – ein warmes, schattenfreies Bernsteingrün, das aus dem Wasser selbst hervorquillt. Fast jede Zylinderoberfläche trägt einen dichten Besatz aus Bakterien, deren dunkle Zellkörper die optisch glatte Frustule zur matten, lichtschluckenden Pelzmasse verwandeln, während das Wasser dazwischen kein Wasser mehr ist, sondern eine kolloidale Suppe aus Polysaccharidsträngen, gelösten Pigmenten und viralen Partikeln, zähflüssig und von biologischer Dichte gesättigt. Es gibt keine Leerstelle: Jede Sichtlinie endet im nächsten Kettenglied, im nächsten Fächer aus Streben, in der nächsten Ansammlung bernsteingelber Trommeln – ein Labyrinth aus Gold und Glas und organischem Dunkel, das sich nach allen Seiten hin in sich selbst verschließt.

Der Blick geht über eine blassgrau-beige Ebene, die sich in einer Trübung aus schwebenden Partikeln verliert – ein Sediment, das über Hunderte von Millionen Jahren aus dem Sterben der Oberflächenblüten entstanden ist und zu einem tiefen, stillen Archiv aus biogenem Opal und Karbonatkalk geworden ist. Aus der Oberfläche ragen *Coscinodiscus*-Frusteln wie halb vergrabene Buntglasfenster hervor, ihre konzentrischen Ringe hexagonaler Areolae noch immer makellos erhalten in amorphem hydratisiertem Silikat, schimmernd in kaltem Aquamarin und verblichenem Gold ohne jedes Sonnenlicht – nur das diffuse Streulicht ihrer eigenen Interferenzfarben. Dazwischen liegen die zerfransten Bruchstücke von *Eucampia*-Ketten und pennate Valven, deren Ränder von dem leicht korrosiven Abyssalwasser kurz oberhalb der Lysokline zu Siliziumspitze aufgelöst werden, während weiße Foraminiferentests als winzige architektonische Ruinen im Sediment sitzen – die löslichsten bereits zu kalkigem Schleier zerfallen. Aus mehreren Punkten im Schlamm pulsieren langsam kalte blaue Biolumineszenzblitze bakterieller Kolonien, die durch die darüber liegenden Frusteln nach oben diffundieren und die Areolae für Bruchteile einer Sekunde wie Glasfaserfelder erleuchten, bevor die Dunkelheit sie wieder auslöscht. Weit in der Trübung erhebt sich eine blasse Sedimentwolke, wo ein Polychaet durch den Schlamm treibt und Jahrzehnte feiner Akkumulation in eine lautlose Wolke aus Frustulenfragmenten aufwirbelt, die langsam aufsteigt und jene bakteriellen Lichtblitze einfängt – jedes taumelnde Stück Diatomeenventil für einen Moment eine kleine, kalte Laterne, bevor das Schwarz es verschluckt.
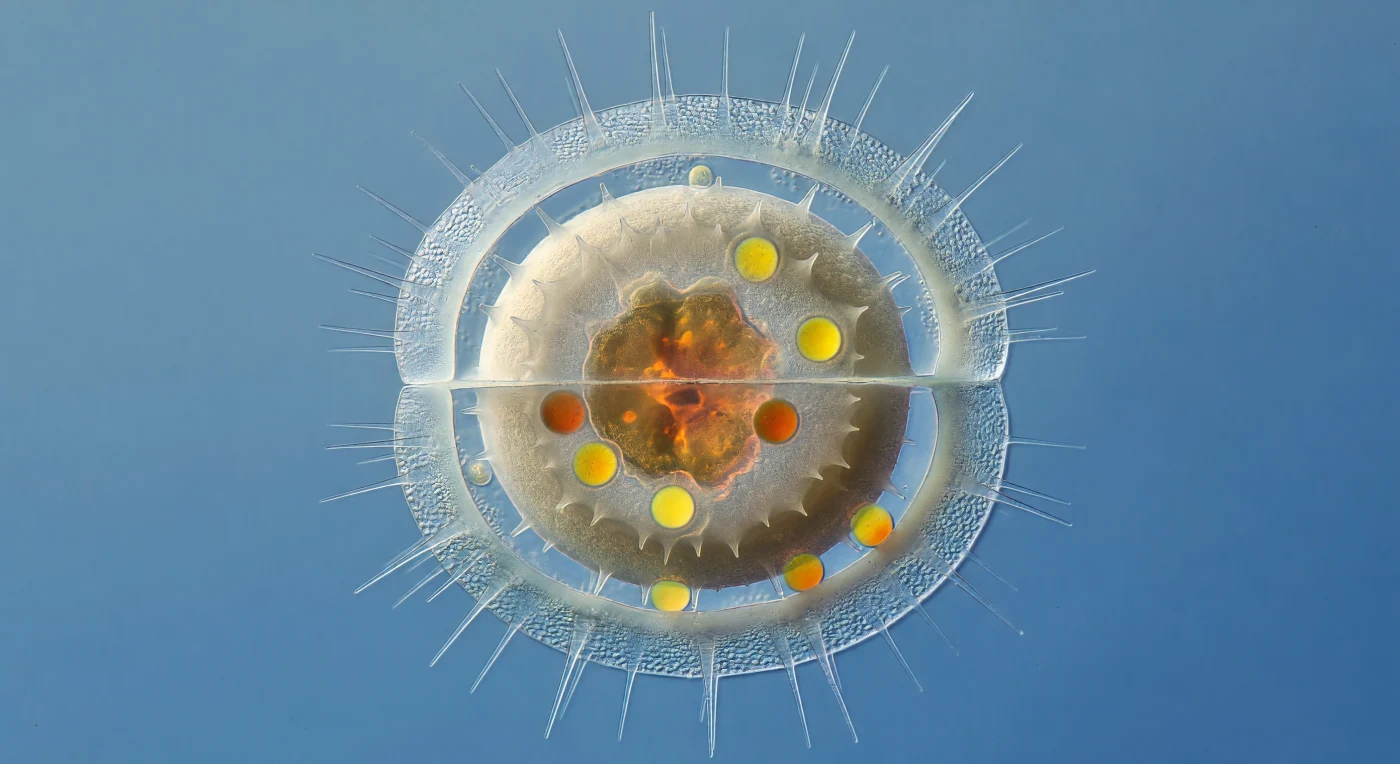
Die Welt, die sich dem Betrachter hier offenbart, ist ein Kontrast zwischen Vergehen und Entstehung: Die äußere Frustule einer *Chaetoceros*-Zelle, kaum mehr als ein Hauch aus hydratisiertem opalem Silizium, umschließt wie ein verblassender Glasschrein das Innenleben einer werdenden Dauerspore. Lange hohle Setae strahlen wie nadelfeine Lanzetten aus der Gürtelregion ins umgebende Wasser, während die dünner gewordenen Schalenwände — einst belebt von Interferenzfarben — nun milchig-blass wie bereiftes Glas wirken. Im Inneren dominiert die Ruhespore mit ihrer zwei Mikrometer dicken, matt-elfenbeinfarbenen Silikatschale das gesamte Bild: ihre granulierte Oberfläche mit stummeligen Stacheln steht in extremem Tonkontrast zur zarten Transparenz des sterbenden Muttergehäuses. Verdichtet im Kern der Spore leuchten ein kollabierter Chloroplastlappen in tiefem Bernstein und mehrere kugelrunde Lipidtröpfchen in glutendem Orange-Gelb — komprimierte biochemische Energie, versiegelt in einer schweren Keramikschale, die selbst wiederum in einem erlöschenden Glaskasten ruht, treibend im kalten Blau des Küstenwassers.

Der Blick öffnet sich in alle Richtungen auf ein endloses, kreideweißes Mosaik aus Kieselglas und Mineralschatten – man hängt reglos im polierten Querschnitt eines miozänen Diatomitefelsens, eingeschlossen in einer Stille, die seit zehn Millionen Jahren ungebrochen ist. Direkt vor einem stapeln sich *Melosira*-Valven wie eine Säule matter Münzen, jede Scheibe vierzig Mikrometer im Durchmesser, die konzentrischen Radiärstriae noch gestochen scharf ins Gestein eingeschrieben; links erhebt sich ein *Stephanopyxis*-Valve als riesiges Kathedralenfenster aus hexagonalen Silikalokuli, erschreckend dünn für etwas, das die Miozänzeit überdauert hat. Zwischen diesen Architekturen der biogenen Kieselsäure klaffen absolute Dunkelzonen – Porenvoids, aus denen einstmals Porenwasser floss und die nun als kalte Negativräume den hellen Silikaglanz der umliegenden Frusteln verstärken. Ein Bruchstück eines fossilen Fischschuppenrandes zieht sich als bernsteinfarbene Kurve durch das weiße Feldgefüge, das einzige organische Leuchten in dieser mineralen Welt. Was hier erhalten ist, bildet ein stratigraphisches Archiv biologischer Präzision: Tausende identifizierbarer Arten, manche zerbrochen, manche makellos, alle auf mikroskopischer Auflösung in Stein eingefroren – ein Sedimentgedächtnis, das Taxonomie in Geologie übersetzt hat.
