Confiance scientifique: Très élevé
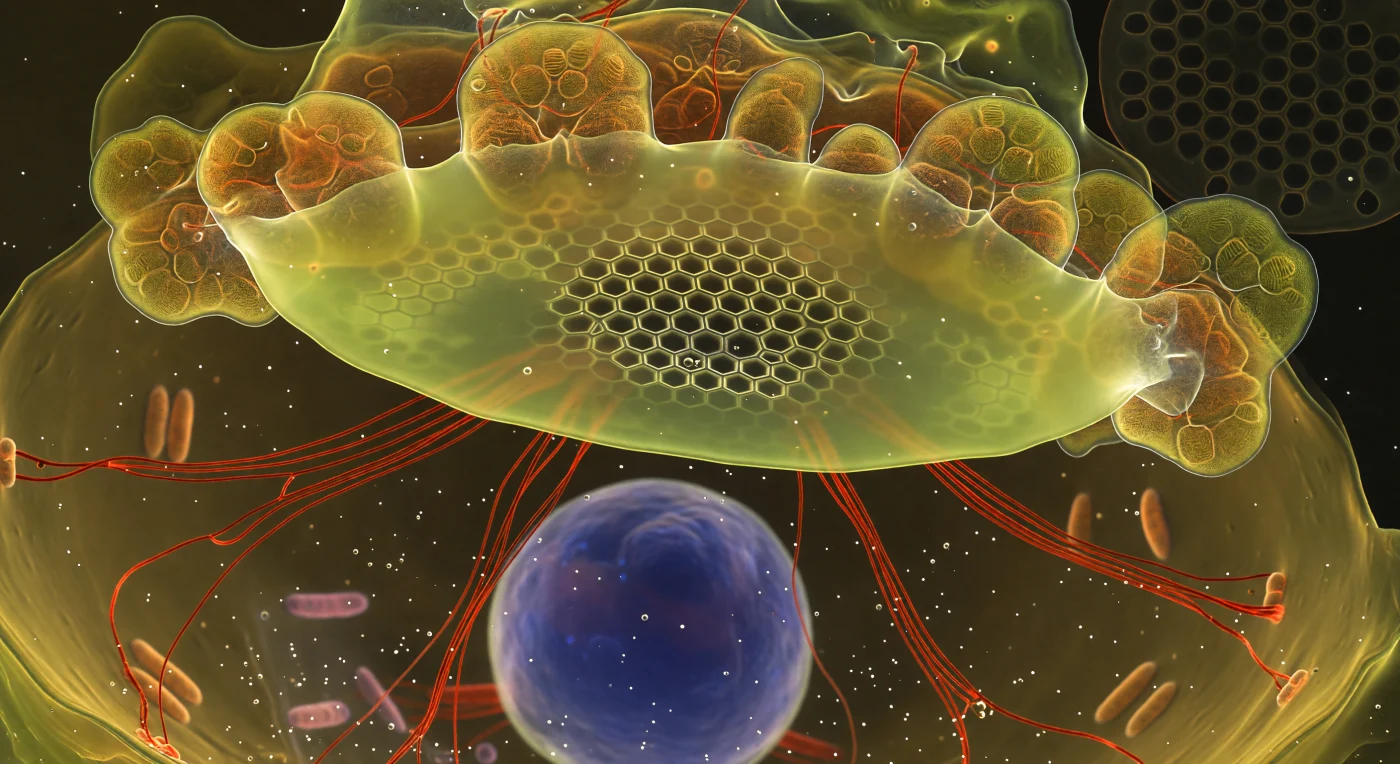
Dans le ventre d'une diatomée en train de se diviser, le spectateur flotte au cœur d'un événement de construction moléculaire d'une précision stupéfiante : la Vésicule de Dépôt Silicique s'étend comme un plafond de cellophane lumineuse en jaune-vert chartreuse, marquée par le fluorochrome PDMPO qui s'incorpore dans la silice fraîchement minéralisée, révélant en son centre un réseau hexagonal déjà cristallisé avec une netteté quasi-violente, tandis que sa périphérie se dissout dans un halo de géométries fantômes encore indécises. Des câbles d'actine F marqués à la phalloïdine rouge-cramoisi rayonnent en faisceaux tendus depuis la face inférieure de la vésicule, guidant avec une force mécanique précise l'expansion du futur squelette de silice vers les bords de la cellule, pendant que de grands lobes de chloroplastes ambré-dorés, translucides comme du verre de cathédrale, viennent presser leurs surfaces lamellaires contre ce plafond minéral en formation. Plus bas, dans le vide cytoplasmique chaud et encombré de gouttelettes lipidiques réfléchissantes, le noyau flotte comme une lune de cobalt pâle, sa lueur DAPI diffuse rappelant que toute cette chorégraphie architecturale est orchestrée depuis cet unique centre de commande — une cellule dont la paroi externe, cette fine dentelle de silice héritée du parent, dessine déjà en filigrane sombre le patron hexagonal que la nouvelle valve reproduit, à l'intérieur de l'intérieur.
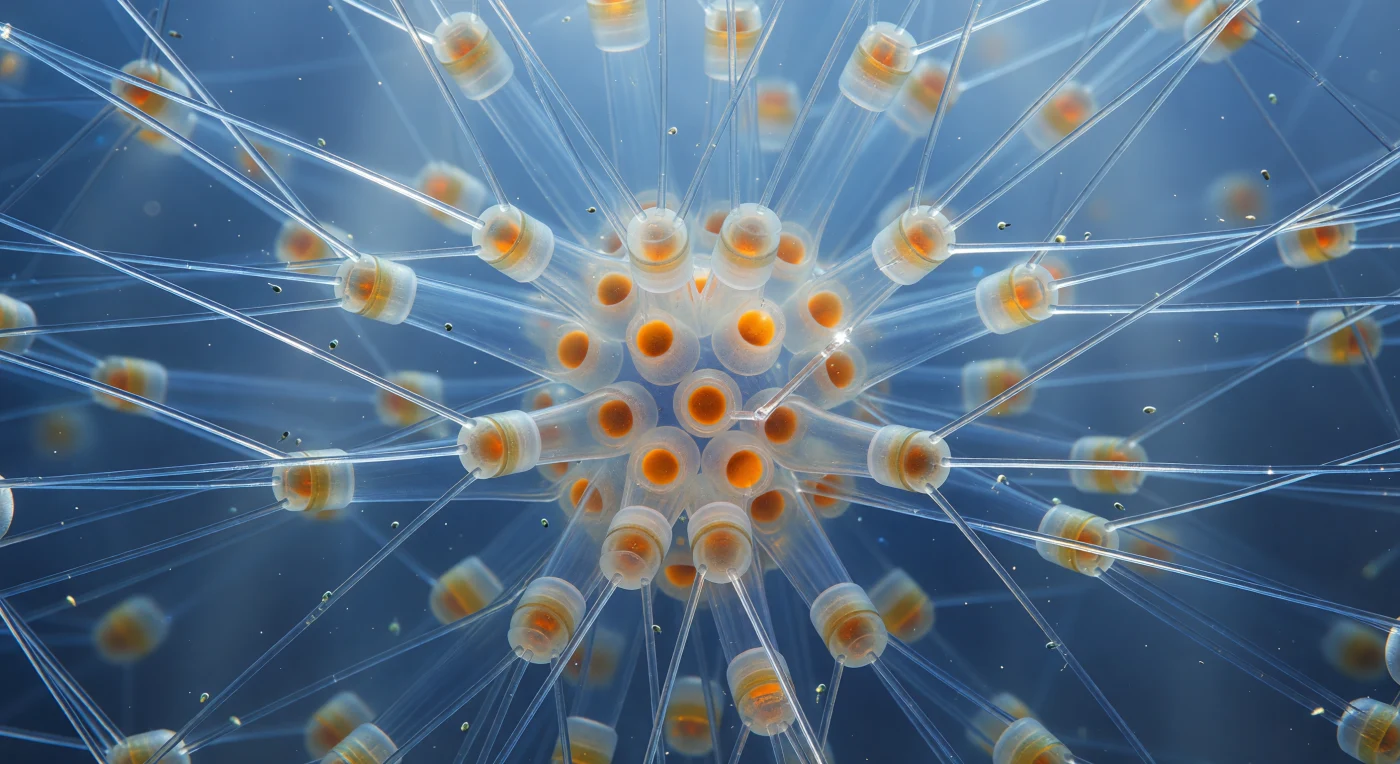
Au cœur de la colonie, vous flottez suspendu dans une cathédrale de verre vivant : dans toutes les directions, les soies siliceuses creuses de *Chaetoceros* s'élancent depuis les corps cellulaires cylindriques comme les nervures d'une architecture impossible, chaque aiguille presque transparente ne concentrant la lumière océanique filtrée qu'en de fines caustiques blanc-or là où ses parois incurvées jouent le rôle de lentilles. Ces setae — jusqu'à 200 µm de long, à peine plus épaisses qu'un fil de quartz soufflé — se croisent en treillis en X entre les cellules empilées, formant une charpente ouverte en trois dimensions qui s'estompe vers l'indigo profond à la périphérie de la colonie. Aux nœuds centraux, les spores de repos brûlent d'un ambre dense et chaud — sphères fortement silicifiées dont les valves épaissies diffusent la lumière comme des braises — tandis que les corps cellulaires eux-mêmes rayonnent d'un brun-sienna doux, trahissant les chloroplastes riches en fucoxanthine pressés contre leurs parois intérieures en verre dépoli. De minuscules flagellés gris-vert dérivent entre les soies portés par des courants invisibles, leurs passages révélés par d'infimes éclats lumineux, simples grains de poussière perdus dans ce lustre de silice biogénique suspendu dans l'océan froid.

Suspendu immobile au-dessus d'elle, le regard plonge vers le bas sur une plaine circulaire de verre qui s'étend jusqu'à chaque horizon — une valve de diatomée géante du genre *Coscinodiscus*, dont la surface sculptée en tessellation hexagonale couvre l'intégralité du champ visuel, chaque chambre de silice amorphe hydratée rayonnant d'une lumière ambrée chaude au cœur du disque, qui vire progressivement au violet-bleu irisé vers la marge où les parois s'amincissent jusqu'à la quasi-transparence, produisant une interférence chromatique typique des films minces de silice opaline. Les aréoles — ces cellules hexagonales de deux cents à cinq cents nanomètres de diamètre, serrées les unes contre les autres en anneaux concentriques selon un ordre radial taxonomiquement précis — agissent chacune comme une microlentille, concentrant la lumière transmise en un point brillant à leur plancher, de sorte que le disque entier se lit comme dix mille cellules lumineuses, une rosace de cathédrale posée à plat dans une eau de mer d'un bleu laiteux. Jaillissant du sol à intervalles réguliers, les fultoportulae — ces processus tubulaires à parois structurées, caractéristiques fonctionnelles des diatomées centriques assurant la sécrétion de mucilage et la flottaison — s'élèvent en cheminées de verre creux dont les bords diffractent la lumière en liserés bleu-froid, projetant des ombres elliptiques douces sur les aréoles voisines et révélant la profondeur topographique subtile de cette architecture biogénique. L'ensemble de la structure — produit d'une déposition siliceuse contrôlée en quelques heures à l'intérieur de vésicules organiques spécialisées — rayonne comme un vitrail suspendu dans l'obscurité bleue de l'océan, monument de précision nanométrique à l'échelle d'une ville entière vue depuis les airs.

À cinq micromètres au-dessus du verre poli, on contemple l'avancée silencieuse d'une cellule de *Navicula* qui traverse ce monde comme une barque de quartz fumé glissant sur une mer immobile — sa coque biconvexe de silice amorphe captant la lumière oblique en un fin croissant argenté qui se fond dans l'ambre chaud de ses flancs, les stries transversales de la valve diffractant une lueur d'interférence en bleu acier et bronze à peine perceptible. À l'intérieur de cette coque translucide épaisse de quelques centaines de nanomètres, deux chloroplastes chargés de fucoxanthine brûlent comme des lanternes d'ambre, leur lumière dorée filtrant à travers la paroi de silice et se déposant en deux lobes oblongs sur le substrat de verre, trahissant la présence d'une machinerie photosynthétique vivante confinée dans une enceinte minérale. Derrière la cellule, extrudé par la fente raphe — couture quasi invisible longue de trente micromètres —, un ruban de mucilage glycoprotéique repose à plat contre le verre en une traîne pâle et réfringente d'un demi-micromètre d'épaisseur à peine, mécanique d'adhésion et de propulsion confondues dans une même sécrétion. Accroché à ce sillage organique comme une virgule sombre sur soie claire, un bâtonnet bactérien de deux micromètres témoigne que même ce chemin éphémère est déjà une niche, une ressource, un territoire colonisé dans l'économie invisible de la pellicule marine.

Au niveau du regard, une colonne de cellules de *Thalassiosira weissflogii* descend dans l'océan de surface baigné de lumière : chaque disque de silice, large d'une centaine de microns, présente sa face comme une rosace de cathédrale, ses aréoles hexagonales difractant la lumière descendante en irisations vertes et dorées sur une trame d'ambre translucide, pendant que les lobes chloroplastiques dorés rayonnent à l'intérieur comme les pétales d'un soleil enchâssé dans du verre antique. Entre chaque paire de cellules, une fibrille de chitine — émise par les fultoportulae et si fine qu'elle demeure presque invisible — s'illumine par instants en un filament argenté lorsqu'un rai de lumière oblique la traverse, maintenant la chaîne entière dans une tension architecturale souple qui lui confère une légère torsion hélicoïdale. À la surface de plusieurs cellules, des bactéries s'accrochent en petites colonies sombres le long des crêtes siliceuses, rappelant que la frustule n'est pas une surface inerte mais un habitat colonisé. Vers le bas, la chaîne se perd dans le bleu volumétrique de la colonne d'eau, son dernier disque se dissolvant en une lueur ambrée fantôme, tandis qu'un agrégat de neige marine dérive en silence sur la gauche — voile translucide de mucilage et de fragments frustulaires — dans la lumière omni-directionnelle de ce bleu cathédral, immense et silencieux.

Dans l'obscurité absolue, vous flottez au centre d'une constellation aplatie sur un seul plan focal, entouré de frustules de diatomées disposées selon l'art des diatomistes victoriens : des architectures de silice amorphe hydratée — l'opale-A biogénique — suspendues dans le vide comme des reliquaires de vitrail éclairés de l'intérieur par une lumière froide et sans source. Directement devant vous, un *Triceratium* triangulaire s'étend sur plusieurs dizaines de longueurs de corps, ses parois de verre épaisses de quelques nanomètres seulement fracturant la lumière en franges d'interférence qui saignent du cobalt électrique vers l'ambre fondu, chaque aréole hexagonale agissant comme un réseau de diffraction miniature projetant des couronnes colorées dans le noir environnant. Plus loin, un *Coscinodiscus* aussi vaste qu'un dôme de colisée rayonne en citrine profonde, ses fultoportulae — ces cheminées de silice caractéristiques des diatomées centriques — projetant de fines colonnes de lumière réfractée dans le vide, tandis qu'un *Arachnoidiscus* irradie, rayon par rayon, comme un lustre de nacre rose pâle. Tout près, une *Pleurosigma* pennate courbe son corps sigmoïde à portée de main, sa grille d'aréoles obliques formant un réseau de diffraction croisé qui fait onduler des vagues de teinte sarcelle à améthyste à chaque imperceptible déplacement du regard, la fente raphéenne brillant d'un blanc argenté là où la paroi de silice s'amincit jusqu'à la quasi-transparence. Ces squelettes minéraux acido-nettoyés, dépouillés de toute matière organique, révèlent la pureté structurelle d'une architecture biologique capable de traverser des millions d'années dans les sédiments — et ce qui ressemble ici à une constellation de gemmes microscopiques est en réalité l'ossature d'organismes qui produisent près d'un quart de l'oxygène atmosphérique terrestre.

Suspendu au zénith d'un continent de verre pétrifié, le regard plonge verticalement sur la valve d'une *Pinnularia nobilis* de cent cinquante micromètres — une plaine convexe de silice amorphe hydratée que la lumière polarisée en contraste interférentiel différentiel cisaille en relief d'orfèvrerie, révélant chaque costae transversale comme un rempart de métal battu surgissant d'une vallée d'ombre bleu-gris. Le sternum axial traverse le paysage du pôle au pôle tel un boulevard de lumière blanche architecturalement bilatéral, interrompu aux nœuds central et polaires par les renflements sculptés du raphé — cette fente de cinquante à deux cents nanomètres de large taillée avec une précision moléculaire dans l'épaisseur même du verre, béant comme un canyon dont les parois descendent vers une obscurité absolue. Sous cette armure minérale froide et métallique, les deux lobes chloroplastiques en H irradient par transparence une chaleur ambrée chargée de fucoxanthine, une lumière d'intérieur organique qui contraste avec la froideur sculpturale de la silice — deux mondes superposés, le biologique et le minéral, séparés par quelques micromètres de paroi translucide. Entre eux, le noyau opalescent flotte comme une lanterne givrée, et l'ensemble du plateau est figé dans cet instant de précision photonique où la physique de l'onde lumineuse révèle, en un seul regard rasant, toute l'architecture d'une cellule qui fabrique sa propre cathédrale de verre.

À un centimètre au-dessus de la vasière intertidale découverte par la marée basse, le sol n'est pas de la vase — c'est une cité vivante de bronze et d'ambre, un tapis continu de frustules pennées serrées les unes contre les autres comme des feuilles de verre martelé, captant la lumière rasante du matin en un chatoiement métallique profond qui ondule en crêtes lentes là où des populations entières de cellules ont migré vers la surface en masse, épaississant le biofilm en bourrelets d'ambre concentré plus lumineux que les marges translucides qui les entourent. Ce tapis n'est pas une simple pellicule pigmentée : c'est une architecture biogénique de silice amorphe hydratée, chaque frustule constituant une coquille à deux valves emboîtées dont les parois nanoporeuses — quelques centaines de nanomètres d'épaisseur à peine — diffractent la lumière visible en éventails spectraux pâles aux bords des valves de Cocconeis aplaties sur les grains de sable, leurs aréoles régulièrement espacées agissant comme des réseaux de diffraction naturels. Dans la matrice de polymères extracellulaires transparents qui lie l'ensemble — un gel aux reflets opalescents, chargé de fibrilles mucilagineuses — une cellule de Gyrosigma progresse en courbe sinueuse, sa fente raphéale en contact avec le substrat, propulsée par le flux d'exsudats muqueux à raison de quelques micromètres par seconde, laissant derrière elle un sillage à peine visible dans la lumière. Des gouttes d'eau de mer en retrait s'accrochent entre les crêtes du mat, magnifiant par lentille les organismes dessous en une mosaïque légèrement distordue d'or et d'ambre, avant de s'évaporer ou de glisser sous un souffle de vent marin — témoins éphémères du passage entre deux mondes, l'aquatique et l'aérien, que ces diatomées traversent chaque jour de marée avec une indifférence chimique absolue.

Suspendu dans la lumière glauque et ambrée de l'épilimnion, vous faites face à une colonie d'*Asterionella formosa* : une étoile de verre à huit bras d'environ 200 µm d'envergure, chaque bras une tige de silice biogénique aussi translucide que du borosilicate poli, légèrement incurvée comme un arc tendu, ses stries d'aréoles captant la lumière descendante en halos d'interférence dorés et argentés. Au centre, un nœud de mucilage ambré — dense comme de la résine de pin ancienne — soude les huit frustules en une rosette maintenue par des liens biopolymères invisibles, pendant que deux chloroplastes chargés de fucoxanthine courent sur toute la longueur de chaque bras, irradiant une chaleur dorée-brune à travers la silice comme des braises enfouies dans du verre. La colonie pivote avec une lenteur imperceptible, soumise à la viscosité intime du milieu aqueux — un fluide si résistant à cette échelle que chaque micron de déplacement exige une force mesurable contre la pression uniforme des molécules d'eau. Dans le fond lumineux, des desmides *Staurastrum* à trois lobes dérivent comme des lanternes de jade, et des filaments cyanobactériens tracent des traînées bleu-vert diffuses dans la colonne d'eau, rappelant que cette architecture de silice biologique s'inscrit dans un écosystème entier baigné de chimie lente et de photosynthèse silencieuse.

Entre deux carapaces brisées de *Fragilariopsis*, le regard est immédiatement capturé par une sphère translucide d'une ampleur qui écrase tout le reste du champ de vision — l'auxospore en pleine expansion, dont la membrane tendue et nacrée reflète par endroits de fugitives irisations dorées là où les bandes siliceuses du périzonium interceptent la lumière transmise. Les deux frustules abandonnées flottent de part et d'autre comme des armures dépouillées, leurs parois de silice striées de côtes parallèles impeccablement régulières, leurs bords fracturés en courbes conchoïdales qui accrochent la lumière ambiante en minuscules éclats bleu-argent — témoins géologiques d'un organisme qui a choisi de tout sacrifier à la croissance. À l'intérieur de l'auxospore, une masse chloroplastique dorée d'une densité presque solide tapisse la membrane de l'intérieur, parsemée de gouttelettes lipidiques qui brillent comme de petits soleils enchâssés dans de l'ambre, tandis qu'une large vacuole centrale creuse un espace de transparence parfaite, sa courbure infléchissant légèrement les rayons lumineux comme une lentille vivante. Ce que l'on contemple ici est l'une des rares occasions où une cellule unicellulaire transgresse les limites de sa propre architecture héritée, s'affranchissant momentanément de la contrainte minérale de la silice pour retrouver une liberté de forme éphémère avant de forger une nouvelle coque.

Dans les entrailles de la glace antarctique, vous flottez à l'intérieur d'un couloir de saumure hypersaline, cerné de parois de glace polycristalline qui irradient en bleu cobalt et en aigue-marine laiteuse, chaque facette cristallographique déviant la faible lumière polaire en coins de cyan, de violet pâle et d'argent froid, transformant cet espace confiné en une cathédrale de verre taillé. Suspendues dans cette colonne de liquide légèrement visqueux et teinté d'ambre, des chaînes de *Fragilariopsis cylindrus* dérivent en arcs et en spirales — de minuscules cylindres de silice de 8 micromètres reliés bout à bout, chaque frustule quasi invisible laissant rayonner de l'intérieur un chloroplaste d'un or sombre et safrané, comme une braise comprimée enfermée dans une lanterne fantôme. Autour de chaque chaîne, un manteau de gel d'EPS antigel enveloppe les cellules d'un halo gélatineux transparent, estompant légèrement leurs contours et capturant la lumière environnante en une couronne prismatique à peine perceptible. Le sol du canal disparaît sous une couche épaisse de biomasse diatomée accumulée — un tapis de résine ambrée et de bronze terni parcouru d'éclats d'or vif là où des chloroplastes demeurent encore actifs — tandis qu'au-dessus, les grandes faces cristallines de la glace redistribuent la lumière en longs faisceaux diagonaux qui illuminent une chaîne, puis une autre, avant de se noyer dans l'obscurité bleu-nuit des profondeurs du canal.

Au-dessus de cette plaine de verre dépoli, le regard se perd dans une géométrie qui déborde toute échelle humaine : des milliers de puits hexagonaux s'étendent à perte de vue, chacun taillé avec une précision absolue dans la silice amorphe d'une valve de *Coscinodiscus wailesii*, leurs parois illuminées en blanc éclatant par un faisceau rasant qui noie chaque fond dans une obscurité totale. Au plancher de ces alvéoles, le velum — une dentelle de silice dont les poroides sont inférieurs à la longueur d'onde de toute lumière visible — tisse une rosace presque invisible, témoignage d'un système de filtration moléculaire façonné par la cellule elle-même lors de la silicification en vésicule SDV. Ça et là, les fultoportulae se dressent comme des cheminées industrielles au-dessus du plateau : colonnes creuses à parois translucides, flanquées chacune de trois pores satellites, conduits par lesquels la cellule sécrétait ses fibrilles de chitine vers l'océan qui l'entourait. À la marge du disque, là où la valve bascule en manteau, les rimoportulae découpent la falaise de silice en fentes sombres et nettes — ouvertures labiales sur des chambres intérieures invisibles — avant que la surface n'abandonne tout ornement pour les bandes lisses du cingulum, cercles de verre nu qui s'éloignent dans une brume d'électrons, rappelant que cette architecture monumentale est l'œuvre d'une cellule unicellulaire de moins d'un demi-millimètre.

Vous planez à peine au-dessus d'une lame de kelp géant qui s'étend comme un continent ondulant d'olive et d'ambre, sa surface une mosaïque de cellules végétales dont les parois luisent sous la lumière filtrée de la zone subtidale, des rideaux caustiques bleu-vert balayant tout le paysage en pulsations lentes. Depuis ce substrat laqué d'une matrice mucilaginaire aux reflets irisés dorés et cuivrés — un biofilm d'exopolysaccharides sécrété par des dizaines d'espèces de diatomées —, des colonies de *Licmophora* s'élèvent sur leurs tiges de mucilage comme des éventails de paon déployés vers la lumière, leurs vingt à quarante cellules siliceuses arrangées en gerbes flabellées d'un or chaud, striées avec une précision nanométrique, vibrant à l'unisson à chaque impulsion du courant. Entre elles, les valves elliptiques de *Cocconeis* restent presque invisibles, plaquées à fleur de kelp, tandis que des rubans de *Rhabdonema* spiralent en chaînes en zigzag dont les arêtes captent la lumière comme des fils d'argent. Au loin, une cicatrice pâle entaille le biofilm — la trace encore fraîche d'un amphipode brouteur dont les mandibules soulèvent doucement les franges de mucilage déchiré, envoyant des ondes de pression qui font frémir chaque éventail de *Licmophora* comme des antennes d'or accordées à la même fréquence lente.

À l'intérieur de la chambre calcaire d'un foraminifère *Amphistegina*, les parois incurvées s'élèvent en voûte sur une centaine de micromètres, leur calcite translucide transmettant depuis le fond sableux une lumière douce et diffuse qui baigne tout l'espace d'un halo ivoire sans source définie. Le cytoplasme de l'hôte occupe le bas de la chambre comme une eau gélifiée et pâle, traversée de fins filaments pseudopodaux qui s'étirent en fils de verre argenté vers les ouvertures en arceaux sombres de la paroi. Parmi ce gel vivant reposent les cellules de *Nitzschia*, diatomées symbiotiques pennées de vingt à quarante micromètres, leurs frustules de silice amorphe renvoyant un reflet irisé bleuté tandis que leurs chloroplastes chargés de fucoxanthine rayonnent d'un ambre chaud, entre vieux miel et cuivre bruni. Ces cellules sont ici non mobiles, tenues en suspension productive dans le cytoplasme de leur hôte, leurs raphes à peine lisibles en fines lignes sombres sur chaque valve, et leurs gouttelettes lipidiques scintillent comme de petites perles de lumière vive perdues dans la chaleur ambrée des plastes. L'ensemble forme un monde clos et filtré où la turbulence de la colonne d'eau est totalement absente, remplacée par ce calme calcaire lumineux et la constellation dorée d'une photosynthèse abritée.
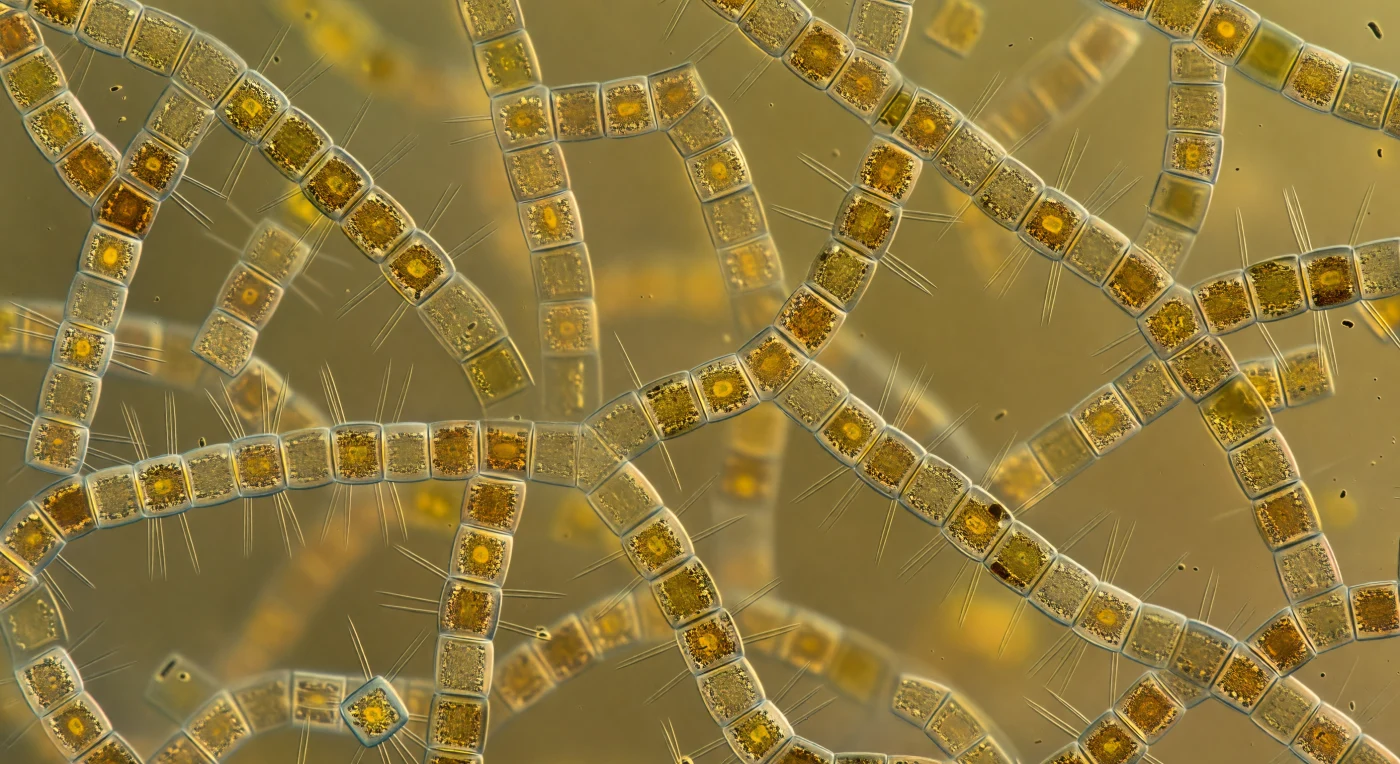
Au cœur de la floraison printanière, vous flottez immobile dans une cathédrale ambrée qui n'a ni murs ni plafond : des tours cylindriques de *Skeletonema costatum* s'empilent dans chaque direction, leurs frustules de verre gossamer reliés par des nervures de silice rayonnantes — les fultoportulae — qui s'entrecroisent comme les armatures d'une cage thoracique explosée, formant un échafaudage tridimensionnel sans interruption visible. Chaque tambour cellulaire, large de cinq à huit micromètres, brûle de l'intérieur d'une chaleur dorée : les chloroplastes chargés de fucoxanthine pressés contre les parois internes transforment chaque cellule en une lanterne d'ambre, si nombreuses que la lumière solaire d'origine — froide et bleu-blanche — a été entièrement digérée par le milieu et renaît comme une radiance volumétrique chaude venue de partout à la fois, sans ombre, sans source localisable. Une fourrure sombre de bactéries en bâtonnets tapisse presque chaque surface de silice, et l'eau elle-même n'est pas vide mais épaissie en une soupe colloïdale de polysaccharides extracellulaires et de matière dissoute, si dense que les chaînes distantes — à peine un demi-millimètre — se dissolvent déjà dans le brouillard ambré avant d'atteindre votre regard. Il n'existe ici aucun arrière-plan libre, aucune ligne de fuite : chaque axe se ferme sur un autre amas de verre et d'or biologique, le milieu entier gorgé de vie jusqu'à la saturation, photosynthétisant en silence dans chaque micromètre cube illuminé.

Au ras du plancher océanique de l'Austral, à trois mille mètres de profondeur, le regard s'étend sur une plaine gris-beige construite frustule par frustule depuis des millions d'années de floraisons en surface : une boue siliceuse diatomifère dont chaque centimètre carré recèle plus d'histoire géologique que l'œil ne peut en saisir. Les valves circulaires de *Coscinodiscus*, larges de quatre-vingts à cent micromètres, émergent de la vase à des angles aléatoires comme des vitraux à demi enfouis dans la cendre, leurs aréoles hexagonales en opal-A encore parfaitement ordonnées captant une iridescence fantôme — aquamarine pâle, ivoire froid — issue de leur propre structure nanoporeuse plutôt que de toute lumière solaire. Entre ces disques, des fragments de chaînes d'*Eucampia* et des valves pennées de *Fragilariopsis* aux marges progressivement dissoutes par les eaux légèrement corrosives proches de la lysocline témoignent d'une dissolution en cours, la frontière entre le verre biogénique et le sédiment s'effaçant en un dégradé de silice de plus en plus lacunaire. Des colonies bactériennes enfouies dans la boue émettent par intermittence de brèves pulsations de bioluminescence bleue qui traversent les frustules translucides et illuminent leurs aréoles comme des réseaux de fibres optiques l'espace d'une fraction de seconde, tandis qu'au loin un polychète progresse sous la surface, soulevant un panache lent de fragments de valves et de poussière carbonatée qui monte en silence dans la colonne d'eau noire, chaque éclat de diatomée devenant brièvement une petite lanterne froide avant que l'obscurité ne l'absorbe.
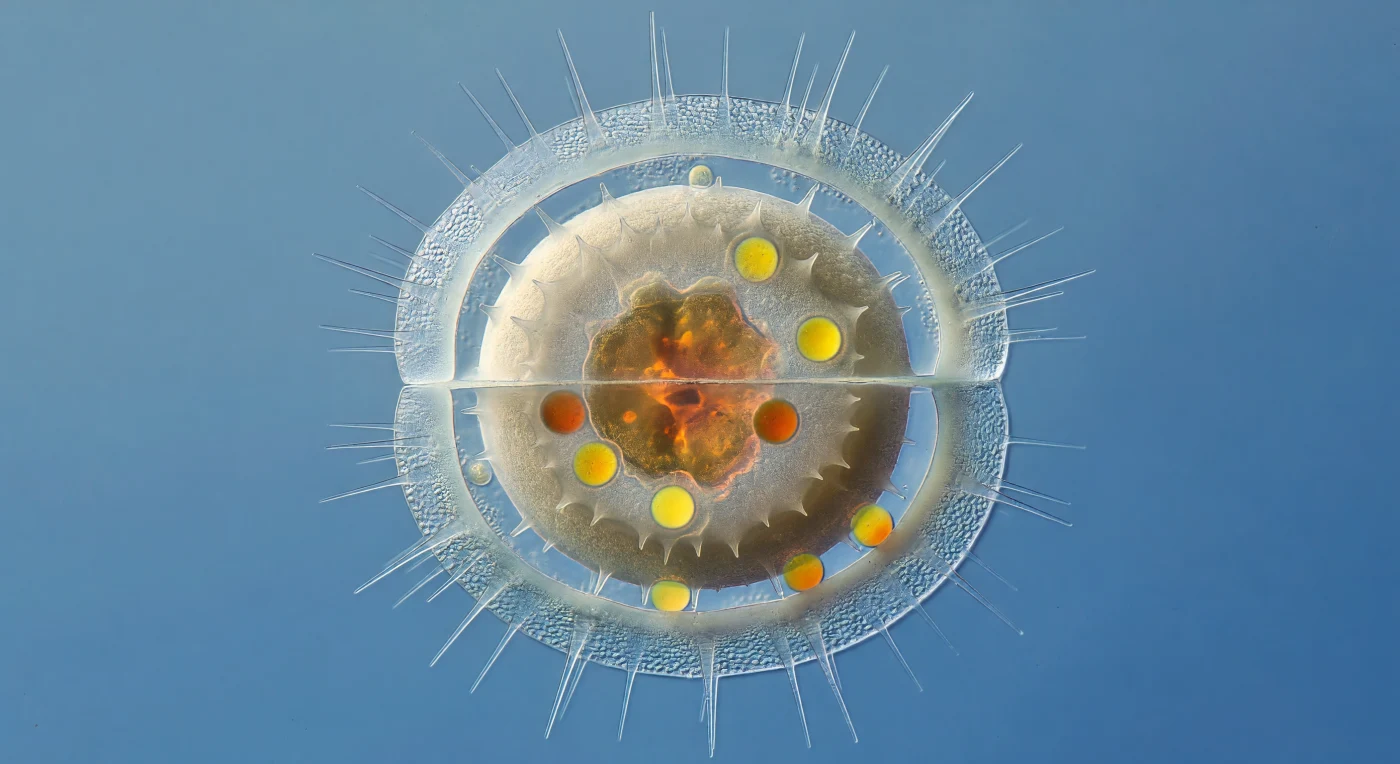
À dix micromètres de la paroi externe, le regard plonge dans l'intérieur d'une frustule de *Chaetoceros* mourante — une lanterne de silice hydratée si mince qu'elle en est presque immatérielle, ses parois opalines réduites à des feuillets de cent nanomètres à peine, ses longues setae creuses rayonnant vers l'obscurité comme des aiguilles de verre oubliées. À l'intérieur de cette enveloppe fantôme gonfle la spore de repos : une masse d'opale granuleuse aux parois de deux micromètres d'épaisseur, opaque et mate comme de la céramique non émaillée, dont les épines émoussées absorbent toute la lumière que la cellule mère laisse encore filtrer. Ce contraste radical entre la translucidité du parent et la densité de l'endospore illustre une stratégie de survie extrême — lorsque les nutriments s'épuisent, la cellule redirige toutes ses ressources vers la construction d'une armure siliceuse capable de résister des mois, voire des années, dans les sédiments. Pourtant, derrière ces murs austères, les contenus du spore rayonnent : une masse de chloroplastes contractés en ambre brûlé, et des gouttelettes lipidiques sphériques d'un jaune-orange incandescent, réserves d'énergie scellées comme des braises dans une boîte d'huître, en suspens dans l'eau côtière froide et cobalt qui entoure tout.

Vous êtes enchâssé dans la chair minérale d'une falaise de diatomite du Miocène, suspendu dans une coupe polie où dix millions d'années tiennent dans chaque centimètre carré de roche blanche. Autour de vous s'étend une mosaïque dense et pâle de frustules fossilisées : des colonnes de valves de *Melosira* empilées comme des pièces de silice, leurs stries concentriques encore résolues dans la pierre ; un disque de *Stephanopyxis* large comme une cathédrale en coupe transversale, ses loculi hexagonaux formant une paroi en nid d'abeilles de verre opaque ; des *Nitzschia* pennées fichées en oblique, leurs raphés striés à peine voilés par la diagenèse sans en être effacés. Les espaces poraux entre les cellules comprimées apparaissent comme une obscurité absolue, des lacunes de néant qui font rayonner par contraste la silice environnante d'un blanc ivoire froid et diffus — la lumière semble sourdre de la matrice elle-même plutôt que de lui parvenir de l'extérieur. Près du bas du champ, un fragment d'écaille fossile de poisson courbe sa surface lisse à travers la mosaïque, seule chaleur ambrée dans un monde par ailleurs entièrement minéral, témoin silencieux que ce compactage de géométries biologiques nano-résolues fut jadis, avant sa lithification, un fond océanique vivant.
