Confiance scientifique: Élevé

Au cœur de cette architecture minérale, le regard traverse trois coques concentriques de silice amorphe dont chaque pore hexagonal fonctionne comme une lentille taillée à l'échelle du micromètre, déviant la lumière océanique en halos prismatiques — violet pâle en bordure, aigue-marine au centre — qui se superposent en moirés géométriques jusqu'à donner l'impression d'habiter une cathédrale dont les voûtes se démultiplieraient à l'infini. Derrière soi, l'endoplasme nucléaire irradie une lueur ambrée et douce, chaleur organique qui contrebalance le bleu froid filtrant depuis les abysses extérieurs à travers les trois lattis progressivement plus ouverts. Douze épines triradiées traversent chaque sphère comme les nervures d'une croisée d'ogives impossible, guidant la lumière ambiante le long de leur axe à la façon d'une fibre optique cristalline avant de se perdre dans un indigo lointain. Cette structure — le squelette siliceux d'*Actinomma asteracanthion* — est le produit d'une biominéralisation conduite vésicule après vésicule, à raison de quelques heures de précipitation lente, dans un milieu où la viscosité domine toute inertie et où la gravité n'existe presque plus. Au-delà de la coque externe, la colonne d'eau se résout en brume particulaire de matière organique dissoute et de fantômes de frustules, chaque fenêtre hexagonale en offrant une version légèrement réfractée, comme si l'œil composé d'un organisme minéral regardait la mer se dissoudre dans la nuit.

Suspendu à deux cents micromètres de la surface de cette créature, vous faites face à l'une des architectures les plus improbables du vivant : un Acantharia, protiste pélagique dont le squelette n'est pas constitué de silice comme ses cousins radiolaires, mais de sulfate de strontium — la célestite — vingt épines cristallines disposées selon une symétrie absolue régie par la loi de Müller, impossible à obtenir par hasard biologique. Sous lumière polarisée croisée, la biréfringence de chaque épine transforme leur anisotropie cristalline en couleurs d'interférence distinctes — cyan électrique, magenta saturé, cobalt profond, or chaud — chaque teinte révélant l'orientation précise du réseau cristallin par rapport aux polariseurs, aucune épine ne partageant exactement la même couleur car aucune n'est orientée à l'identique dans l'espace tridimensionnel. Au centre de ce brasier géométrique, la capsule centrale est un ovale sombre et dense, halogène ambré des corps de zooxanthelles serrés contre sa paroi — algues symbiotiques de cinq à quinze micromètres dont les pigments chlorophylliens captent la lumière solaire descendante pour nourrir l'hôte en sucres photosynthétiques. Entre les épines, presque invisibles dans l'éclat polarisé, les câbles myonémaux tirés comme des filins d'orgue mécanique relient les spines à la capsule, capables de contracter l'ensemble du squelette en quelques secondes, ajustant la profondeur de flottaison de l'organisme dans une colonne d'eau que vous percevez maintenant comme un milieu visqueux et actif, peuplé de particules colloïdales qui scintillent faiblement dans le courant brownien.

À dix microns au-dessus de la surface, on se croit posé sur la paroi d'une planète de verre — le test siliceux d'une cellule de Sphaerozoum s'étend dans toutes les directions comme un dallage de cathédrale où chaque fenêtre hexagonale ou pentagonale est taillée dans une opale froide, les arêtes plus épaisses tirant sur l'or ambré, les struts les plus fins virant au bleu-gris translucide, et chaque nœud triple renvoyant l'éclairage oblique en un éclair d'argent dur qui découpe une ombre rasante dans le puits noir voisin. Depuis les rebords des pores et depuis les nœuds entre eux, une forêt d'axopodes jaillit verticalement dans le liquide au-dessus de nous : des tiges rigides d'un micron de diamètre, lisses comme du cristal, striées en long par les faisceaux de microtubules internes, et bordées d'une couture spectrale de lumière DIC — un fin ruban teal, ambre, magenta — qui court sur toute leur longueur avant de se perdre dans l'obscurité fuligineuse du milieu marin chargé de protéines et de complexes organiques en suspension brownienne. Le long de trois des axopodes les plus proches, des sphères topaze — vacuoles alimentaires emballant chacune une proie digérée dans une membrane dense et réfringente — rampent imperceptiblement vers la surface squelettique, portées par le flux cytoplasmique qui court sur l'armature invisible des microtubules, entourées d'un léger halo de microlentille là où leur indice de réfraction diffère de l'eau de mer, lanternes d'ambre suspendues dans une nuit minérale absolue.

Au fond de cette chambre intérieure, les parois de silice opale vous enveloppent comme les voûtes d'une cathédrale gothique sculptée dans de la glace marine — translucides, légèrement laiteuses, leurs rangées de pores elliptiques disposés en spirale laissent entrer l'océan sous forme de taches d'aquamarine tremblantes qui se superposent et interfèrent sur les nervures minérales, transformant toute la surface interne en une mosaïque vivante de lumière réfractée. Cette architecture n'est pas de verre ni de cristal mais de l'opale amorphe biogénique — de la silice hydratée (SiO₂·nH₂O) déposée par voie enzymatique à l'intérieur de vésicules de silicification, couche par couche, au cours des heures précédant votre arrivée. En levant les yeux à travers les cinq septa annulaires qui s'empilent vers le haut en perspective forcée, chaque foramen central agit comme un diaphragme naturel qui concentre et déplace vers le bleu la lumière de l'océan, jusqu'à ce que le cinquième anneau ne soit plus qu'un point incandescent de turquoise derrière lequel la colonne d'eau s'étend dans toutes les directions à des distances inimaginables depuis ici. À l'apex, l'épine apicale se découpe en parfait silhouette noire contre cet azur aveuglant — tige de silice dense, bord capturant un fin liseré de réfraction blanche — dernière structure solide avant l'immensité pélagique où les axopodes rayonnent déjà vers l'extérieur comme des filaments de lumière cristalline à la pêche dans la soupe bactérienne de l'océan ouvert.

Au-dessus de cette plaine sans horizon, suspendu à quelques millimètres seulement du fond de l'abîsse, le regard plonge sur une nécropole siliceuse d'une précision géométrique troublante : des centaines de tests radiolaires pressés dans l'argile grise, leurs lattices sphériques et coniques intactes ou fracturées, leurs pores perçant la lumière rasante d'électrons comme les fenêtres aveugles d'une cathédrale engloutie. L'éclairage oblique — froid, implacable, sans couleur — ne laisse aucune nuance intermédiaire : chaque surface tournée vers le faisceau surgit en blanc d'os, chaque creux s'effondre dans un noir absolu, révélant sans merci la granularité de l'opale amorphe, les microfissures propagées sous la compression géologique, les particules de silt logées dans des pores individuels comme des débris dans des vitres brisées. Ces organismes unicellulaires, qui fabriquaient leurs squelettes de silice par dépôt minéral intracellulaire en quelques heures à peine, ont coulé lentement — à raison d'un à dix mètres par jour — depuis la zone photique jusqu'ici, s'accumulant sur des millions d'années en une boue siliceuse qui recouvre des millions de kilomètres carrés des grands fonds océaniques. Ce que l'on contemple n'est pas un désert, mais une archive : une stratigraphie comprimée de vies mathématiques, chaque test un fossile d'une complexité architecturale que l'évolution a maintenue presque inchangée depuis le Cambrien.
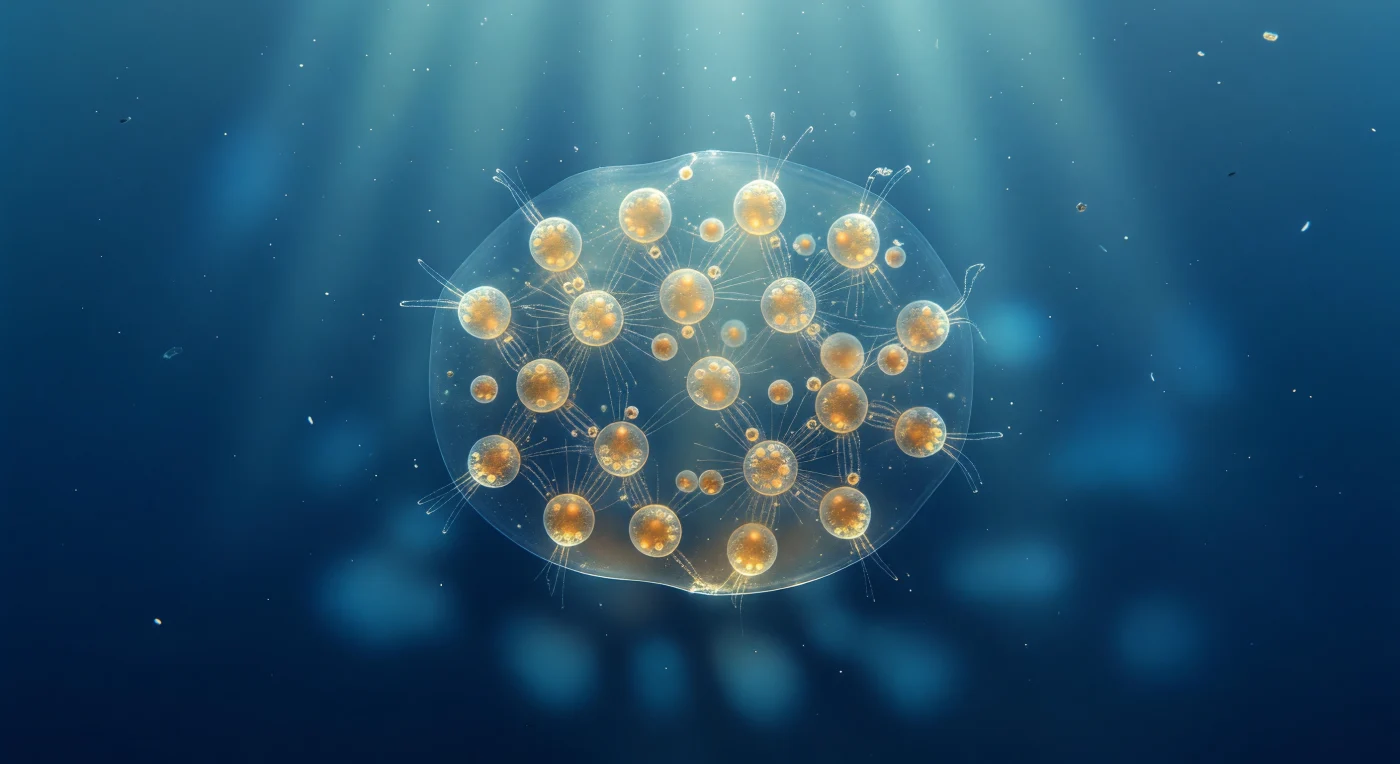
À un millimètre à peine de sa surface, la colonie de *Sphaerozoum* occupe tout le champ de vision comme une planète translucide suspendue dans un vide azuré — une ellipsoïde de mucilage gélatineux de huit millimètres, si proche de la transparence parfaite qu'elle se révèle moins par sa matière que par ce qu'elle contient : des dizaines de cellules individuelles distribuées comme des lanternes d'ambre chaud dans le gel, chacune d'environ trois cents micromètres, densément peuplées de dinoflagellés symbiotiques qui réchauffent la lumière tropicale descendante du vert aquamarine vers l'ocre et le miel. Ces cellules collodaires sont dépourvues de squelette siliceux — nul réseau géométrique, nulle cathédrale gothique — et leur flottabilité repose entièrement sur des vacuoles lipidiques qui captent la lumière en perles spéculaires irisées, conférant à chaque organisme une lueur opalescente autour de son noyau doré de symbiontes. Les axopodes rayonnent de chaque cellule en filaments cristallins presque invisibles jusqu'à ce qu'un rai de lumière les révèle comme des aiguilles d'argent s'étendant dans le gel commun, tissant un réseau filamenteux lâche qui relie l'intérieur de la colonie entière. Depuis les profondeurs indigo qui s'ouvrent en dessous, la colonie projette vers l'obscurité un caustique dapplé de lumière mouvante — fantôme optique entre invisibilité totale et rayonnement intérieur, vitrail à la dérive dans un silence bleu immense.
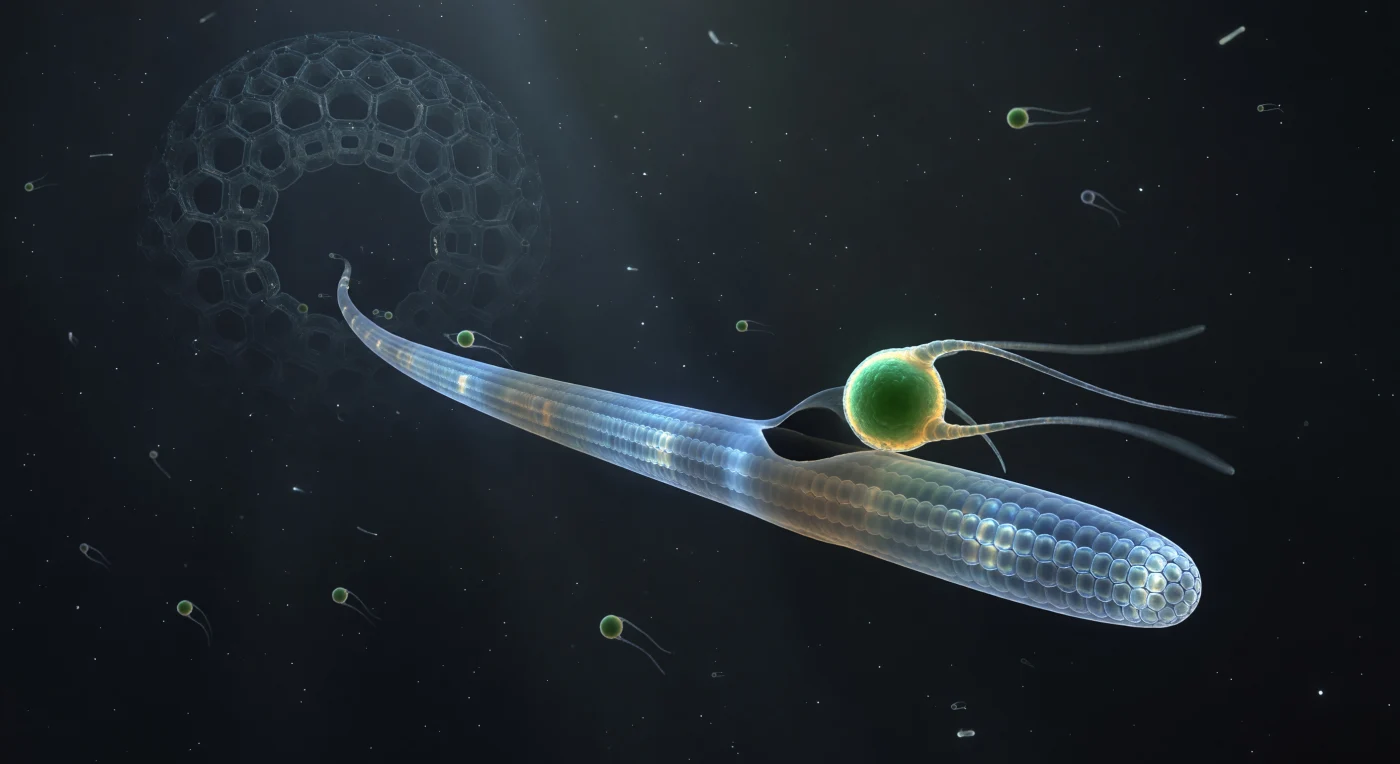
Dans l'obscurité totale d'une colonne d'eau océanique, on assiste à l'instant précis où une flagellée émeraude — Micromonas, à peine trois microns de diamètre — entre en contact avec un axopode radiolairien, cette longue lance de cytoplasme translucide soutenue en son cœur par des faisceaux de microtubules parallèles disposés en réseau hexagonal, visibles comme des fils de lumière froide courant sur toute la longueur de la structure. L'axopode, dont le diamètre représente à cette échelle celui d'un pont de verre tendu dans le vide, diffracte les photons ambiants en halos prismatiques qui lui confèrent une incandescence intérieure froide, tandis qu'au point de contact la membrane se déforme légèrement et qu'un croissant sombre — à peine une ombre, presque pas une matière — commence son premier degré d'arc autour de la proie lumineuse, signal biochimique d'une englobement en cours mais pas encore accompli. Les deux flagelles de la flagellée se figent dans leur battement, chacun une filature de lumière pure aux bords dissous dans l'ambiguïté brownienne, la chloroplaste brûlant d'une autofluorescence vert saturé comme une petite planète chaude prise dans l'étreinte inexorable d'un prédateur aux gestes lents. Plus loin dans l'axopode, vers l'obscurité cathédrale de la capsule centrale, des formes vésiculaires dérivent vers l'intérieur comme des lanternes dans un courant imperceptible — repas antérieurs déjà vacuolisés, leurs pigments autrefois vifs estompés en rouille et ocre sous l'action de la digestion en cours.
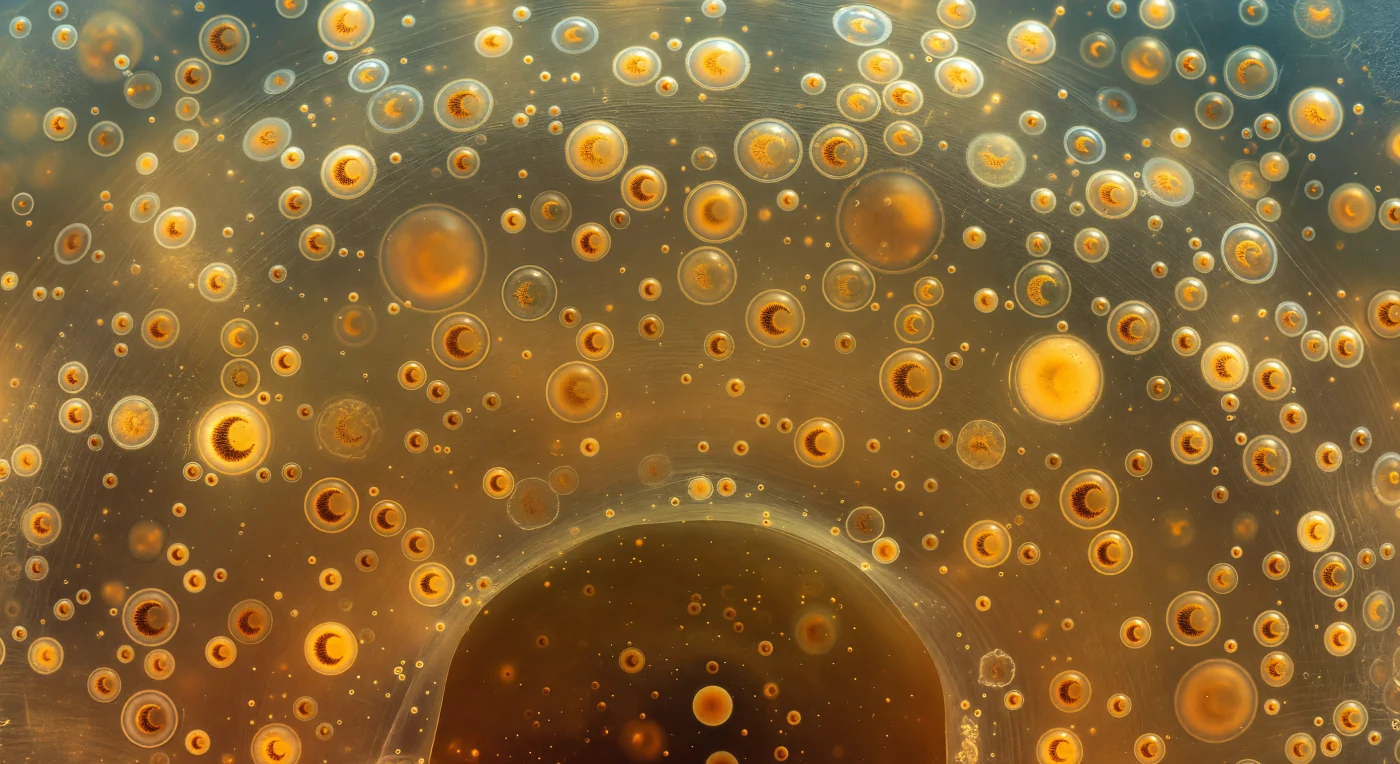
Au sein de la calymma d'un Collodaire colonial, le regard ne trouve nulle limite nette : dans toutes les directions flottent des zooxanthelles dorées, sphères de dix à quinze micromètres suspendues dans l'ectoplasme viscoélastique comme des lanternes d'ambre dans un air parfaitement immobile, chacune portant en son centre un noyau en croissant — arc ochre-brun, dense et légèrement granuleux — distinctement lisible à cette proximité. Le milieu lui-même n'est ni liquide ni solide, mais un gel de glycoprotéines d'une translucidité absolue, parcouru d'une fine dentelle de filaments à peine résolus que seule la lumière rasante révèle, et ponctué de vacuoles lipidiques sphériques de trente à quarante micromètres dont la paroi courbe la lumière ambiante en une caustique annulaire dorée sur tout ce qui flotte derrière elles. La lumière descend de quelque part au-dessus — direction devinée plutôt que vue —, filtrée par des couches successives de cellules symbiotiques et de vacuoles jusqu'à se transformer en une luminescence ambrée diffuse qui semble émaner du milieu lui-même plutôt que de le traverser. Plus loin, les zooxanthelles se multiplient en une brume chaude qui absorbe la profondeur comme du velours absorbe la lumière, tandis que la silhouette de la capsule centrale se devine à peine au loin — membrane organique incurvée, sombre, scellant l'endoplasme comme la paroi d'un monde clos illuminé par une braise enfouie dans du miel.
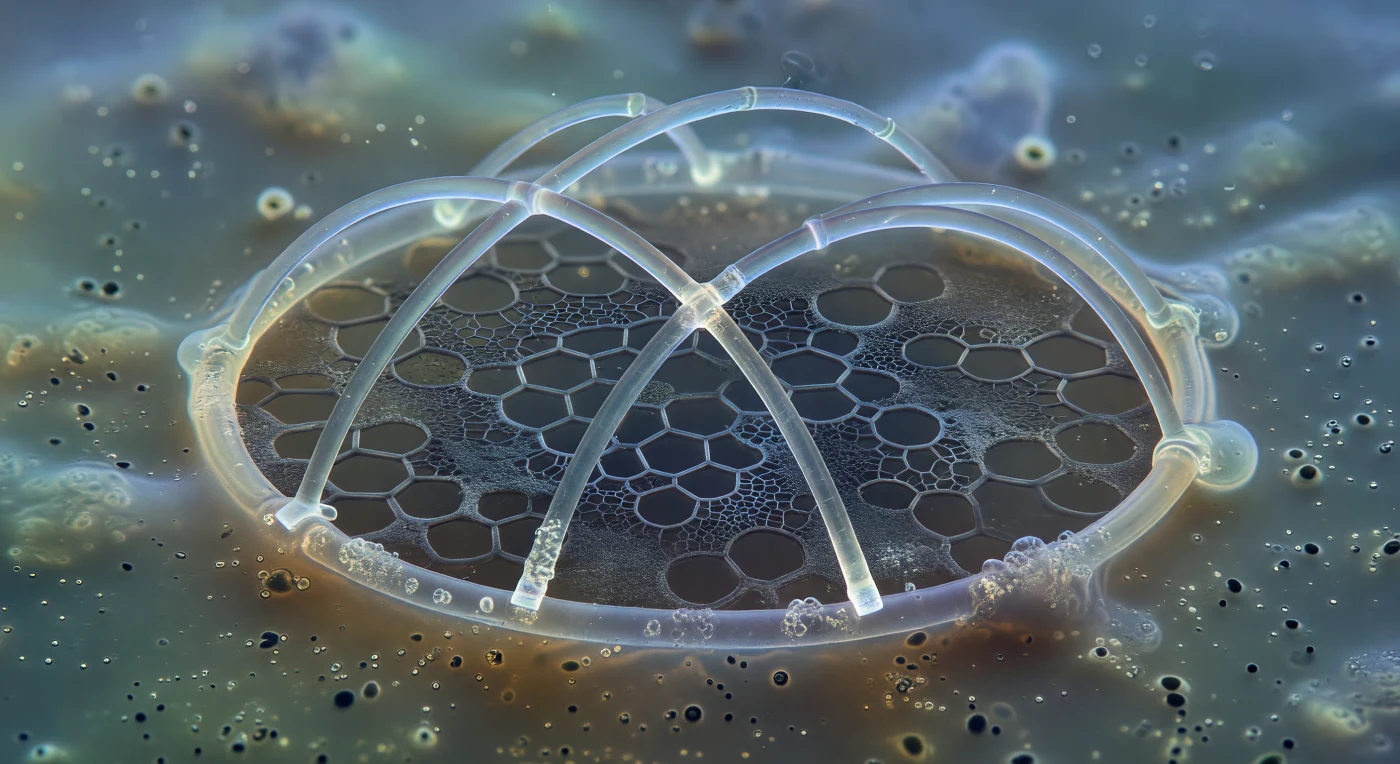
Au sein d'un milieu aqueux chargé comme de la glycérine, un anneau sagittal de *Pterocorys* s'élève en arcs lumineux et vitreux, ses quelques microns de silice déposée brillant d'un éclat blanc-or cerné d'halos de contraste de phase, pareils aux nervures d'une cathédrale à peine sortie du chantier. Deux barres primaires traversent l'intérieur de l'anneau à des angles précis, leurs extrémités fraîchement formées encore entourées du scintillement fantomatique des vésicules de dépôt siliceux. Derrière la minéralité lumineuse, à peine perceptible, le réseau protéique organique s'étend comme du givre sur du verre sombre — une dentelle de glycoprotéines mate et grise, qui précède le minéral et en trace la géométrie future : des hexagones négatifs déjà inscrits dans la matière organique, vides et en attente. Le cytoplasme ambiant, trouble et agité, se perd progressivement dans un halo aquamarine pâle, la calymma extracapsulaire noyant toute frontière nette entre la cellule et l'océan ouvert. Ce squelette naissant, patient et transparent, déposera les fondations d'une structure capable de traverser cinq cents millions d'années de temps géologique, bien après la disparition de la cellule qui le construit.

Vous êtes plaqué contre la paroi intérieure d'une vésicule de dépôt siliceux — la silicalemma se dessine au-dessus de vous comme une double ligne d'obscurité absolue, précise au rasoir, épousant une courbure dont le rayon suggère une sphère d'une ampleur presque inconcevable à votre échelle. Devant vous, le front de gel siliceux avance sur son échafaudage de protéines silaffine avec la lenteur délibérée d'une coulée volcanique qui refroidit : masse pâle et grise, semi-transparente en ses marges, ses bords effilochés s'enfoncent dans la trame de fibrilles rouille-terracotta comme du givre qui cristallise sur du grillage métallique. Ces protéines fibrillaires constituent le gabarit organique indispensable à la biominéralisation — sans ce réseau chargé positivement qui concentre l'acide silicique et catalyse sa polymérisation, nulle architecture siliceuse ne pourrait émerger avec la précision géométrique caractéristique des radiolaires. Au-delà de la membrane, dans le cytoplasme ambré qui baigne l'extérieur du sac, des formes lumineuses — un globule cobalt qui pourrait être une mitochondrie, une goutte lipidique jaune d'ambre aplatie contre la paroi — flottent comme des lanternes derrière du verre dépoli, rappelant que cette fonderie minérale silencieuse est aussi, irréductiblement, vivante.

Sous vos pieds s'étend un plain courbe de silice grise pâle, percé d'une tessellation quasi parfaite d'hexagones dont chaque ouverture plonge dans un noir absolu — vous vous tenez sur la coquille externe d'un *Hexacontium*, radiolaire sphérulaire dont le test amorphe d'opale siliceuse mesure à peine deux cents micromètres de diamètre, un monde entier contenu dans l'épaisseur de deux cheveux humains. La surface, amorphe et non cristalline, tient davantage du verre borosilicaté givré que du minéral feuilleté : chaque barre de silice reliant les pores fait trois micromètres de largeur, et aux nœuds d'intersection, de petites apophyses nodulaires gonflent comme des perles soudées à chaud, captant la lumière rasante venue de la gauche et projetant des reflets spéculaires froids contre le gris mat général. Six épines triradiées surgissent de la trame à intervalles réguliers, s'élevant quarante à cinquante micromètres au-dessus du sol — des obélisques de silice extrudée dont la base fusionne sans couture dans les nœuds de la lattice et dont la pointe se perd vers le haut comme un gratte-ciel vu depuis le caniveau, leurs faces éclairées luisant d'un blanc argenté tranché, leurs faces ombragées dans le néant. À travers les pores, dans l'obscurité partielle en dessous, on devine une seconde sphère concentrique dont les hexagones décalés créent un moiré de vides emboîtés — l'architecture en poupées russes propre au genre, déposée couche par couche dans des vésicules intracytoplasmiques au fil des heures, tandis que l'organisme dérivait vivant dans les eaux pélagiques il y a peut-être des millions d'années.

Au cœur de la zone mésopélagique, à cinq cents mètres sous la surface, le regard se perd dans une procession silencieuse de tests nassellaires qui descendent de toutes parts — cônes à plusieurs chambres, formes en pagode, casques grillagés — leurs lattis de silice amorphe captant les derniers photons bleutés filtrés depuis le monde du dessus et les redistribuant en éclats froids le long de chaque barreau et de chaque bordure de pore. Les cellules encore vivantes pressent leur cytoplasme ambré et ocre contre les parois intérieures du squelette, faisant brûler chaque test habité comme une lanterne de verre coloré au sein d'un univers minéral et glacial, tandis que les tests vides — nombreux, et en route vers les sédiments — ne renvoient qu'un reflet argent pâle, leurs chambres creuses ouvertes sur l'obscurité comme des orbites dans un crâne de dentelle siliceuse. Entre ces architectures dérivantes s'intercalent des flocons de neige marine — mucus transparent, matière organique morte, cytoplasme radiolairien délié — qui tourbillonnent en spirale lente, captent une bioluminescence venue de nulle part identifiable et la restituent en lueur nacrée avant de s'accrocher un instant à une épine saillante puis de se libérer doucement. Chaque test en sédimentation transporte en lui une portion de l'histoire géologique du vivant : depuis le Cambrien, ces squelettes de quelques centaines de micromètres s'accumulent sur les fonds océaniques en boues radiolaritiques épaisses de kilomètres, témoins siliceux d'un flux ininterrompu de matière organique vers les abysses.

Sous la lumière polarisée du microscope pétrographique, le regard plonge dans un champ de ruines siliceuses qui s'étend à l'infini — une mosaïque comprimée de tests recrystallisés en quartz microcristallin et en calcédoine, sphères écrasées, cônes déformés et ovales affaissés soudés les uns aux autres par cent cinquante millions d'années de pression lithostatique. La lumière transmise monte d'en bas en un flux froid et uniforme, diffusant à travers toute la masse minéralisée pour conférer à chaque surface une lueur intérieure spectrale — non pas brillante, mais suffuse, comme du clair de lune figé dans du verre dépoli — tandis que de minuscules cristallites de quartz captent la lumière polarisée à des orientations légèrement différentes, produisant au sein d'un même test un subtil chatoiement de gris argent et de crème pâle. Entre ces formes fantômes, le ciment interstiel se teinte de rouille profonde et de cramoisi sombre là où des oxydes de fer ont infiltré les anciens espaces poreux emplis d'eau de mer, traçant en négatif la géographie d'un océan disparu. Ces radiolaires — microplancton siliceux qui peuplaient la colonne d'eau jurassique avant de sombrer lentement vers les abysses — ont accumulé leurs tests sur le fond océanique jusqu'à former d'épaisses couches de radiolarite, archives minérales d'une biosphère pélagique que la diagenèse a transformée en ce paysage de pierre phosphorescente et de temps comprimé.

Dans l'obscurité absolue créée par les polariseurs croisés, vous flottez au cœur d'un ciel nocturne miniaturisé sur deux millimètres de réalité : des dizaines de radiolaires Acantharia s'embrasent simultanément, chacun irradiant une signature chromatique distincte — magenta et or pour l'un, sarcelle électrique et violet profond pour un autre — leurs vingt spicules de sulfate de strontium convertissant la lumière polarisée en couleurs d'interférence pures grâce à leur biréfringence cristalline, chaque tige minérale se comportant comme un prisme à géométrie icosaédrique imposée par la logique biologique de l'espèce. Entre ces détonations chromatiques lentes dérivent les tests des Spumellaria, dont la silice opale amorphe reste aveugle au champ polarisé et apparaît comme une architecture fantôme de dentelle grise — cages sphériques concentriques percées de pores hexagonaux, reliées par des struts radiaux, cathédrales gothiques rendues en brume d'argent froid. Là où une épine acanthaire effleure un test spumellaire, un bref point d'ambre surgit, le matériau biréfringent imposant sa couleur au seul point de contact géométrique. Cette scène, qui dans l'espace réel tiendrait dans la paume d'une main, se lit ici comme un volume cosmique habité, les axopodes invisibles des cellules les plus proches frôlant votre présence tandis que les organismes les plus lointains se dissolvent en piqûres de couleur pure avalées par le noir.

Dans le cône d'un projecteur submersible, *Aulacantha scolymantha* surgit de l'obscurité à deux mille mètres de fond comme une étoile à part entière — ses cinq cents spicules siliceux creux irradient en tout sens depuis un corps à peine plus large qu'un cheveu humain, conduisant la lumière bleue-blanche jusqu'à leurs pointes distales où elle ressort en points froids, une constellation sphérique suspendue dans un vide qui ne connaît ni fond ni paroi. Au cœur de la créature, le phaeodium — masse brune et opaque de matières partiellement digérées — refuse toute transmission lumineuse, donnant à l'organisme un noyau solide et étrange autour duquel le maillage siliceux de la capsule centrale n'existe que par instants, comme de l'argent fugace arraché à l'obscurité par le faisceau. La calymma, cytoplasme extracapsulaire gorgé de vacuoles, diffuse la lumière en une lueur douce entre ce noyau sombre et la forêt de spicules, un halo de rétrodiffusion froide qui prête à l'ensemble une présence presque gazeuse. À cette échelle, la pression de deux cents atmosphères n'est pas une abstraction mais une réalité physique qui comprime le milieu lui-même, et le tremblement brownien aux extrémités des spicules — microtubules et silice oscillant au seuil de la résolution — ressemble moins à une réaction biologique qu'à une respiration du froid. Au-delà du halo illuminé, le noir est structural : non pas une absence, mais une matière que la lumière teste et refuse d'entrer, l'organisme seul maître de ses coordonnées dans un espace qui n'accepte aucune autre mesure.
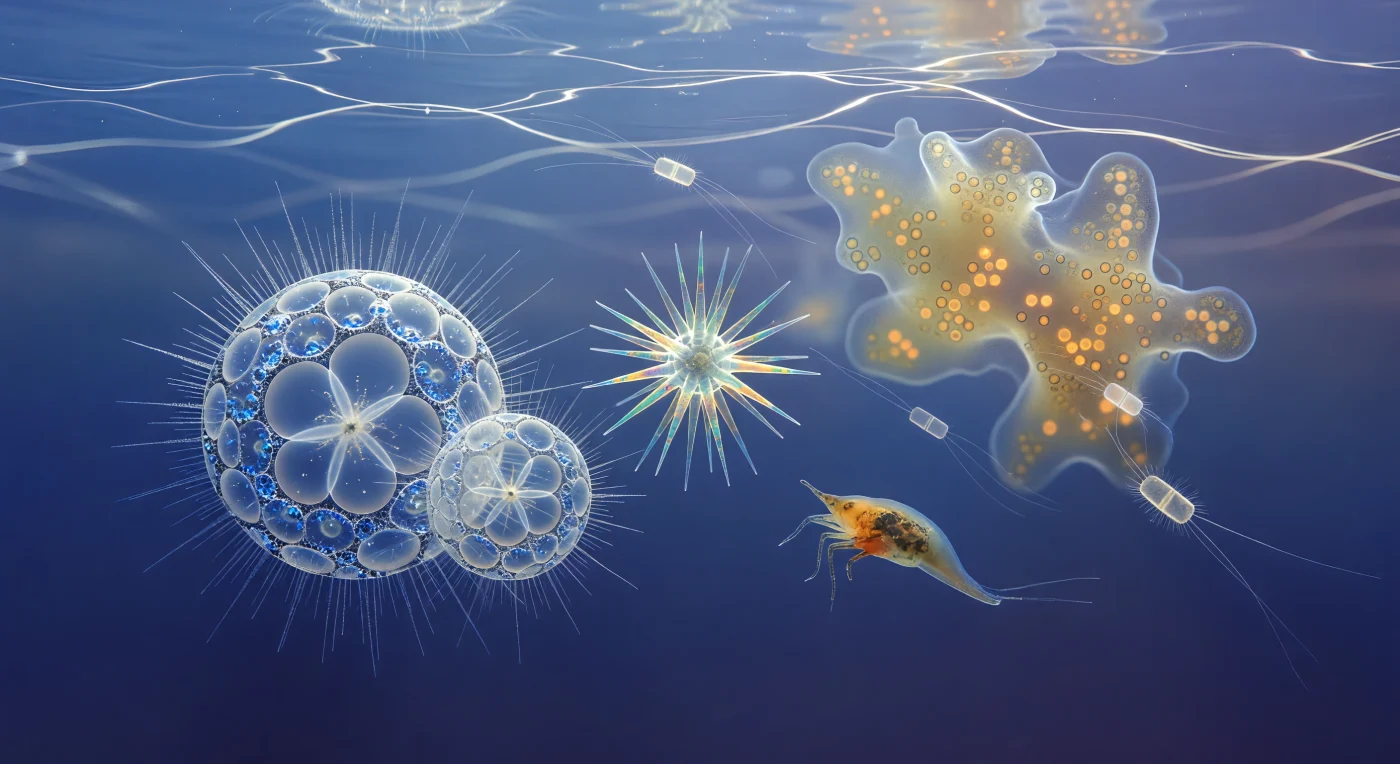
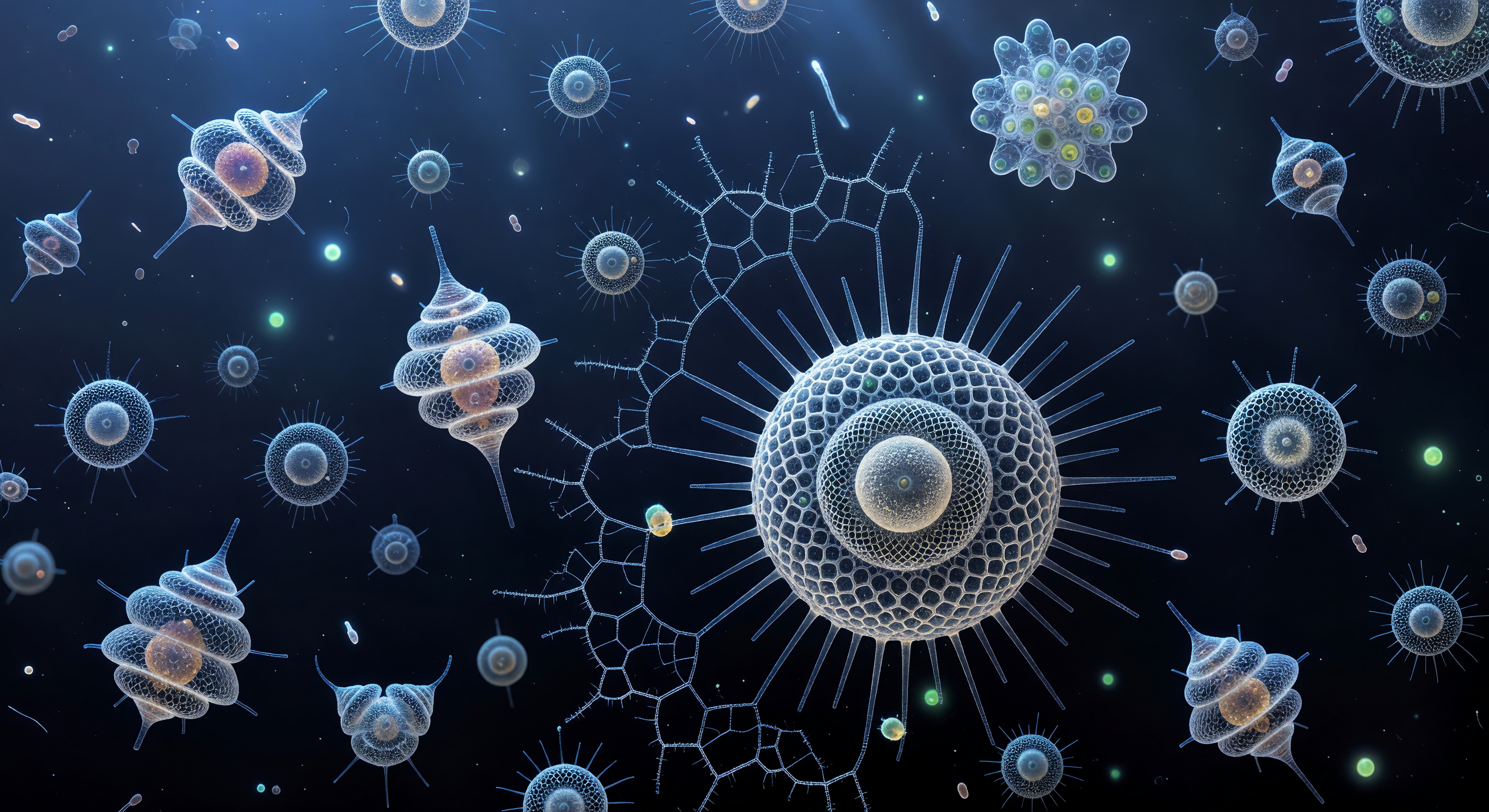
Suspendu à trente mètres de profondeur dans le Pacifique tropical, le regard embrasse un univers où la matière vivante et la lumière se confondent en une chorégraphie silencieuse : au-dessus, la surface de l'océan est un miroir d'argent fracturé dont les caustiques descendent en tresses blanches ondulantes, balayant lentement des architectures d'une complexité vertigineuse. Deux individus de Sphaerozoum flottent à portée immédiate, leurs lattices de silice amorphe évoquant des cathédrales gothiques suspendues dans le bleu, chaque pore hexagonal agissant comme une microlentille qui disperse la lumière en étincelles prismatiques froides, tandis que leurs couronnes d'axopodes — filaments rigidifiés par des faisceaux de microtubules cristallins — s'enflamment brièvement en halos d'argent chaque fois qu'une caustique les traverse. Derrière eux, une Acantharia tourne lentement sur elle-même, ses vingt spicules de sulfate de strontium disposés en géométrie icosaédrique révélant leur biréfringence sous la lumière oblique : ils changent d'amber en sarcelle électrique en rose irisé au fil de la rotation, pendant que dans le lointain la colonie collodaire s'étale comme une nébuleuse ambrée, sa matrice gélatineux commune illuminée de l'intérieur par des milliers de dinoflagellés symbiotiques dont la chaleur orangée contraste avec le froid bleu indigo de l'eau libre. Entre toutes ces structures dérivent des diatomées Chaetoceros — leurs frustules de silice comme des paires de vitres givrées traçant des aiguilles d'argent éphémères —, et un nauplius de copépode, monstrueux à cette échelle, bat ses appendices en arcs rythmiques dans une eau qui n'est jamais vraiment vide : saturée de brume bactérienne et d'organiques dissous, elle donne à chaque lointain une légère auréole de flou lumineux où la physique de la viscosité, et non de l'inertie, gouverne chaque mouvement.