Confianza científica: Muy alto
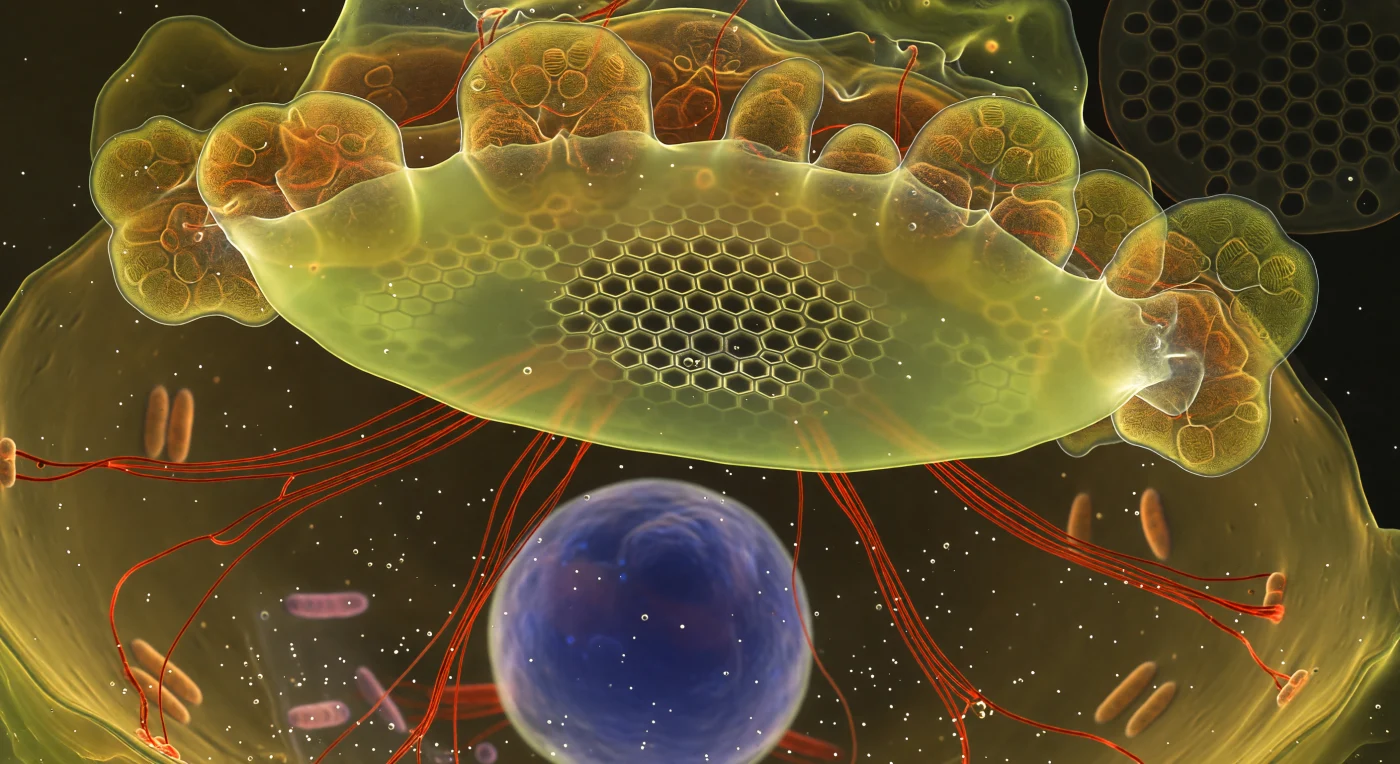
Flotas en la oscuridad interior de una diatomea viva, suspendido frente a una membrana translúcida que se extiende como un techo luminoso de celofán sobre todo tu campo visual: la Vesícula de Deposición de Sílice, impregnada de tinte PDMPO, irradia una fluorescencia amarillo-verdosa que ilumina el espacio circundante con la calidez de una catedral submarina. En su centro, la arquitectura de sílice ya ha tomado forma definitiva —hexágonos perfectos, arreolas oscuras enmarcadas por paredes finísimas que brillan blanco-dorado como metal incandescente— mientras hacia la periferia la geometría se deshace en una niebla luminosa de material a medio mineralizar, el vidrio biogénico aún negociando su estructura final. Cables de actina teñidos de rojo arterial irradian desde la base de la vesícula como tensores mecánicos, guiando con precisión molecular la forma de la valva que nace; cloroplastos ambarinos se curvan contra el techo de sílice como formaciones geológicas translúcidas, y en la profundidad del citoplasma, el núcleo flota como una luna de cobalto pálido rodeada de vacío viscoso. Estás dentro del acto mismo de la biomineralización —un proceso que dura horas pero que aquí se congela en un instante de violencia geométrica silenciosa, mientras la célula construye, desde adentro, el exoesqueleto de vidrio que la definirá.
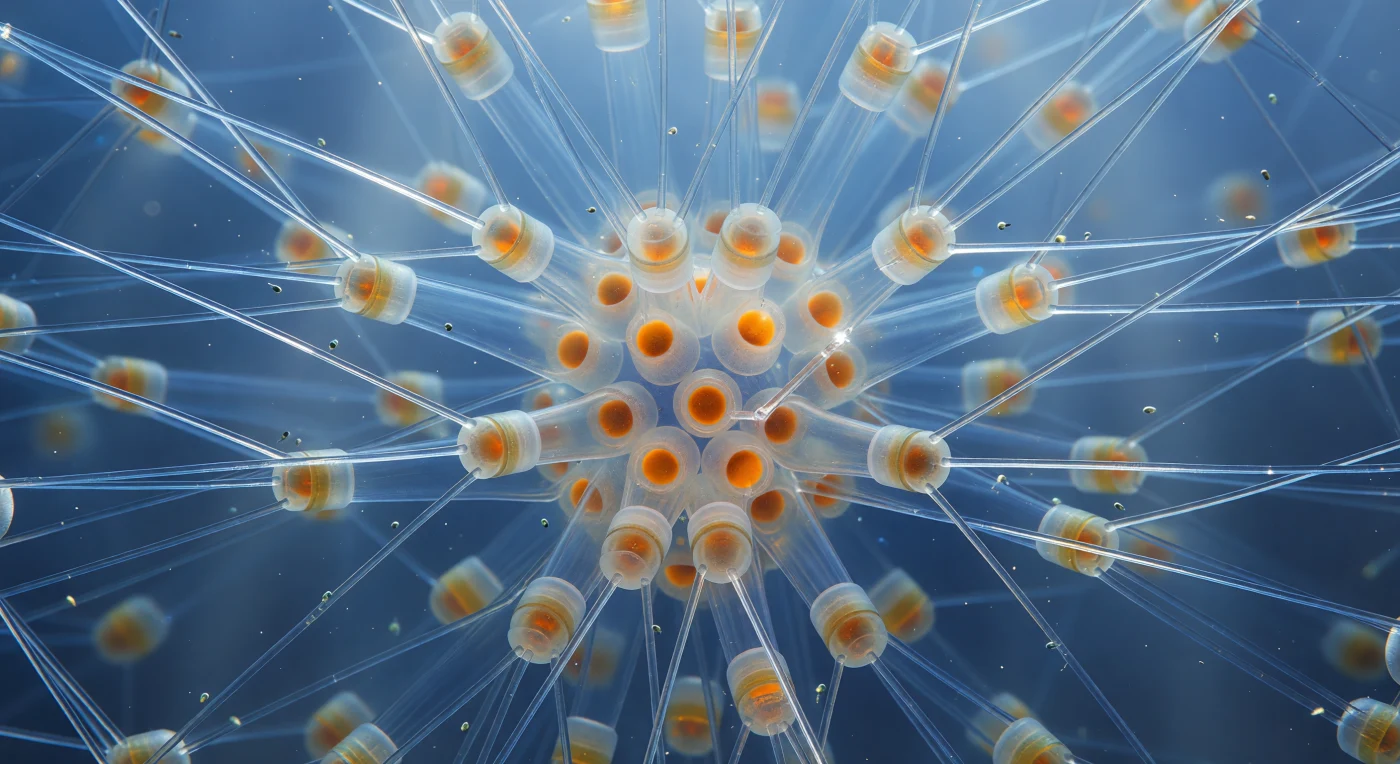
En el corazón geométrico de una colonia de *Chaetoceros*, el espectador flota suspendido en un interior oceánico de una arquitectura imposible: en todas las direcciones, agujas de sílice hueco se extienden desde los cuerpos celulares cilíndricos como las costillas de una catedral de vidrio, casi transparentes, sus paredes tan delgadas que el azul oceánico las atraviesa limpiamente, interrumpido apenas por filamentos dorados donde la luz superficial se concentra en cáusticas brillantes al refractarse en cada superficie curva. Estas setas —estructuras biogénicas de sílice amorfo hidratado secretadas por cada célula durante su desarrollo— se entrecruzan en celosías tridimensionales en forma de X entre los cuerpos celulares apilados, formando un andamiaje cristalino que se pierde en la penumbra índigo de la periferia de la colonia. En el núcleo denso, las esporas de reposo arden con un ámbar más saturado, esferas fuertemente silicificadas cuyas valvas engrosadas dispersan la luz de manera difusa, como brasas cálidas dentro de la geometría fría, mientras que sus cloroplastos ricos en fucoxantina presionan desde el interior con tonos siena quemado. Flagelados de escala nanométrica derivan entre las setas sobre corrientes invisibles, motas de vida orgánica suspendidas en el mismo espacio que esta estructura mineral milenaria, recordando que este bosque de vidrio biogénico es también un ecosistema en miniatura que sostiene cadenas tróficas enteras del océano abierto.

Flotando inmóvil sobre la válvula, el observador contempla una planicie circular de vidrio que se extiende hasta cada horizonte, esculpida en una teselación perfecta de cámaras hexagonales de sílice amorfa hidratada —cada una un cuenco de paredes transparentes empaquetado sin interrupción desde el centro del disco hasta su margen más lejano, captando la luz transmitida que asciende desde abajo en un espectro que va del ámbar cálido en el corazón del disco hasta el violeta-azul iridiscente donde las paredes de sílice adelgazan casi hasta desaparecer. Esta arquitectura no es casual: la *Coscinodiscus* ha depositado su frústula mediante vesículas de deposición de sílice —los SDV— en un proceso de biomineralización finamente regulado que organiza decenas de miles de areolas en anillos concéntricos, cada poro actuando como una lente diminuta que concentra la luz ascendente en un brillo suave en su base. Emergiendo de la planicie a intervalos regulares, los fultoportulae se alzan como chimeneas cilíndricas de vidrio hueco, estructuras de anclaje y secreción cuya función biológica conecta el interior celular con el océano exterior, y que aquí capturan el retrolluminado en destellos ámbar puros o en frías aureolas azules según el ángulo de difracción en sus paredes. El medio circundante —agua marina con su transparencia levemente cianada y sus velos de materia orgánica disuelta— no ofrece ni suelo ni cielo, solo esta vasta mandala de vidrio vivo suspendida en el agua, una ventana de catedral dispuesta en horizontal que irradia color e interferencia de películas delgadas como si albergara su propia fuente de luz interior. La estructura entera, con su lustro de vidrio opalino y sus hilos plateados de cresta entre cámaras, es simultáneamente esqueleto mineral y organismo vivo, condenada a sedimentar en los fondos oceánicos durante millones de años cuando la célula muera, preservando esta geometría perfecta como registro fósil en las capas de diatomita.

Suspendido apenas cinco micrómetros sobre una llanura de vidrio que se extiende hasta todos los horizontes, el observador contempla cómo una célula de *Navicula* de treinta micrómetros avanza lentamente hacia la derecha como una barcaza cruzando un puerto en calma, su casco biconvexo de sílice captando la luz oblicua en una cresta de plata-blanca que se funde en oro cálido y ámbar sobre sus flancos, mientras las finas estrías transversales de la valva generan un leve destello de interferencia en azules acerados y bronces. Desde el interior del casco translúcido, dos grandes cloroplastos cargados de fucoxantina irradian una luz dorado-ocre saturada que se filtra a través de las paredes de sílice como luz de vela a través de papel de arroz, proyectando en el suelo de vidrio dos lóbulos de calor ambarino que palpitan suavemente al ritmo de la corriente citoplasmática. Detrás de la célula, el canal de la rafe deposita una cinta de mucílago casi invisible —media micra de espesor— que refracta la luz del sustrato en un halo cremoso y dorado pálido, rastro bioquímico que adhiere al *Navicula* a su camino como una quilla silenciosa sobre el vidrio. Aferrada a esa estela, ocho micrómetros por detrás, una única bacteria en forma de bastón —dos micrómetros de longitud oscura y densa— permanece inmóvil como una coma sobre seda pálida, retenida por la misma química pegajosa que guía al diatomeo. La atmósfera de este mundo es la de una luminosidad oceánica fría: iluminación azul-blanca difusa desde arriba, calor ámbar emanando desde el interior del casco vivo, y el suelo de vidrio encendido desde abajo en colores de interferencia que recuerdan a una vidriera de catedral sumergida.

Flotas a la altura de los ojos frente a una columna viva de células de *Thalassiosira weissflogii* que desciende por el océano iluminado por el sol, cada disco de silicio translúcido —ancho como una plataforma de vidrio antiguo y cálido— presenta hacia ti su cara valvar como una rosa de catedral, con su geometría radial de areolas refractando la luz descendente en verdes y dorados iridiscentes que cambian con el más mínimo giro de perspectiva. A través de la sílice traslúcida se adivinan los lóbulos de los cloroplastos dispuestos como pétalos de un girasol ocre y ámbar, salpicados de gotitas lipídicas que brillan como cuentas de cristal suspendidas en resina. Entre cada par de células, una única fibrilla de quitina —tan delgada que solo existe cuando un haz de luz oblicua la enciende como un hilo de plata— une los fultoportulae célula a célula, manteniendo toda la cadena en una tensión arquitectónica tensa y helicoidal que gira lentamente a la deriva, mientras bacterias adheridas como motas oscuras recubren las superficies de sílice con una rugosidad biológica imperceptible. La cadena se desvanece en la penumbra azul a pocos diámetros celulares de profundidad, y un agregado de nieve marina —una maraña pálida de mucílago y fragmentos de frústulo roto— pasa a la deriva como un fantasma translúcido, retroluminado en sus bordes por las columnas de oro difuso que descienden desde la superficie. Todo el conjunto existe como un monumento vertical solitario en una catedral oceánica infinita, donde la escala de la arquitectura biogénica rivaliza con la vastedad azul que la rodea.

En la oscuridad absoluta, te encuentras suspendido frente a una constelación inmóvil de arquitecturas de vidrio que se extiende hasta los límites de tu percepción: frústulas de diatomeas dispuestas en el clásico arreglo radial de los microscopistas victorianos, cada una flotando en el vacío como una vidriera iluminada por una luz sin origen. El *Triceratium* que ocupa el centro de tu campo visual es una estructura triangular de sílice amorfa —ópalo-A hidratado, el mismo mineral que compone el nácar— cuyas paredes de apenas nanómetros de grosor fracturan la luz en franjas de interferencia que sangran del cobalto eléctrico al ámbar fundido; cada areola hexagonal actúa como una red de difracción independiente, proyectando coronas de color hacia el negro circundante como linternas perforadas. Más allá, un *Coscinodiscus* del tamaño de un coliseo arde en ámbar citrino, sus anillos concéntricos de cámaras hexagonales idénticos en proporción a una ciudad vista desde el aire, mientras un *Pleurosigma* sigmoideo —su cuerpo curvado como un trazo de caligrafía en turmalina verde— cambia de verde azulado a amatista con cada variación de perspectiva, pues su grilla oblicua de areolas funciona como una red de difracción cruzada. Lo que contemplas no es una preparación microscópica sino un universo arquitectónico completo: estructuras biológicas que la evolución perfeccionó durante doscientos millones de años para gestionar el intercambio de nutrientes y luz, ahora reducidas —o más bien, reveladas— a su esqueleto mineral puro, limpiado con ácido, eterno.

Suspendido sobre una vasta llanura esculpida que se extiende hasta el horizonte en todas las direcciones, el observador contempla desde lo alto una superficie de sílice amorfa que la luz polarizada del contraste diferencial de interferencia transforma en un paisaje de metal trabajado: las costae transversales avanzan como crestas paralelas de piedra pulida a lo largo de todo el continente valvar, separadas por bahías rectangulares donde el frente de onda polarizado oscila del blanco hueso al gris paloma. Por el eje central corre el esternón como una autopista luminosa de luz casi blanca, interrumpida en el ecuador y en cada polo por la hendidura del rafe — una grieta de oscuridad absoluta tallada con precisión molecular, con nódulos que asoman como cantos rodados pulidos que capturan la luz asimétrica del DIC en una cara y la sombra azul fría en la otra. Bajo toda esa arquitectura mineral, dos enormes lóbulos de cloroplasto en forma de H irradian un oro ámbar cálido a través de la pared de sílice translúcida como brasas vistas a través de vidrio, y entre ellos el núcleo descansa como un óvalo opalescente levemente luminoso, creando el contrapunto entre biología cálida y mineral frío que confiere a este paisaje de ciento cincuenta micrómetros la escala y la autoridad de un mundo completo.

La superficie ante ti no es barro — es una metrópolis viviente de bronce y ámbar que se extiende hasta cada horizonte, una alfombra de frústulos de sílice empaquetados tan estrechamente que sus flancos pulidos se tocan en una lámina dorada continua, capturando la luz rasante de la mañana como millones de espejos cóncavos en miniatura. Este tapiz es un biofilm microfitobentónico, una comunidad dominada por diatomeas pennadas que migran activamente hacia la superficie durante la bajamar, guiadas por gradientes de luz y humedad a través de la matriz de polímeros extracelulares —EPS— que segrega sus rafes, ese surco de cincuenta a doscientos nanómetros de anchura por el que fluye el mucílago impulsor. Las crestas de tono ámbar más intenso revelan dónde poblaciones enteras de células han ascendido en masa, engrosando la película viva en cordilleras de pigmento y vidrio que palpitan con una luminosidad casi metálica, mientras las células de Gyrosigma serpentean entre ellas como canoas de caoba pulida navegando un río de glicerina opalescente, y los granos de arena emergen como cordilleras recubiertas de valvas de Cocconeis adheridas de manera permanente, cuyas estrías de sílice actúan como redes de difracción que dispersan la luz solar en abanicos espectrales de azul frío y oro cálido. Las gotas de agua de mar en retirada se aferran entre las crestas como lupas temporales, magnificando el tapiz dorado bajo ellas antes de evaporarse, y hacia el horizonte lejano el mat se sumerge bajo láminas delgadas de agua turbia que viran su color del oro brillante al olivino profundo, dibujando la frontera exacta entre el mundo aéreo y el intermareal que este biofilm lleva millones de años conquistando y abandonando con cada marea.

Suspendido en el agua verdácea y ambarina del epilimnion de un lago tranquilo, el ojo encuentra ante sí una estructura de geometría casi imposible: la colonia de *Asterionella formosa*, una estrella de ocho brazos de poco más de doscientas micras de diámetro, construida enteramente en sílice biógenica amorfa, cada brazo una varilla translúcida que se curva levemente desde el nudo central de mucílago como si hubiera sido tallada en vidrio de borosilicato por una mano de precisión inhumana. La luz descendente del lago se fragmenta al atravesar las paredes de cada frústulo —apenas un par de micras de grosor— y reaparece como halos de interferencia dorados y plateados que bordean cada estrías y cada areola, mientras dos cloroplastos ricos en fucoxantina recorren la longitud de cada brazo como lingotes de ámbar encendidos desde dentro, bañando el agua circundante en una luminiscencia parda y cálida. El cojín de mucílago central retiene los ocho brazos en una roseta mediante enlaces biopoliméricos invisibles, y la colonia entera gira con la lentitud de un mecanismo de relojería sumergido, respondiendo a las fuerzas de arrastre viscoso de un medio que, a esta escala, se comporta más como jarabe que como agua. Al fondo, desmidias de *Staurastrum* de tres lóbulos derivan como linternas de jade, y filamentos de cianobacterias se disuelven en la penumbra del lago como hilos de humo azulado, recordando que esta arquitectura de cristal vivo no es una rareza sino el tejido invisible del que está hecha la productividad de los lagos del mundo.

En este instante de quietud acuática, el espectador flota frente a un acto de renacimiento que apenas comienza: a cada lado, las frustrulas vacías de *Fragilariopsis* derivan con la parsimonia de armaduras abandonadas, sus superficies estriadas captando la luz transmitida en destellos de azul acero y verde fantasmal —interferencia óptica en paredes de sílice hidratado de apenas centenas de nanómetros de grosor— mientras sus bandas del cíngulo se separan con una lentitud casi geológica. Entre esos cascos rotos y oscuros, la auxospora domina el campo visual como un mundo propio: una esfera translúcida que ya quintuplica el diámetro de las células que la generaron, su membrana tensa y perlada cruzada por las bandas transversales del perizonium —arcos de sílice apenas resueltos, trazados como meridianos en plata— que centellean en oro iridiscente allí donde la luz los roza en ángulo oblicuo. En su interior, una masa continua de cloroplastos dorados prensa contra la membrana desde adentro, salpicada de glóbulos lipídicos que arden como soles miniaturizados en ámbar, mientras una gran vacuola central —perfectamente transparente, ligeramente más refractiva que el citoplasma circundante— actúa como una lente viva que empuja todo el contenido celular hacia afuera, hacia el límite en expansión. Este proceso de sexual rejuvenecimiento —la auxosporulación— existe porque la reproducción vegetativa por bipartición reduce progresivamente el tamaño celular, y solo a través de esta esfera efímera, protegida por su perizonium de sílice de nueva síntesis, el linaje recupera su tamaño máximo y reinicia el ciclo.

Te encuentras suspendido dentro de un canal de salmuera de doscientas micras que atraviesa el hielo marino antártico como una fisura en un geoda de vidrio polar, sus paredes de hielo policristalino irradiando una luz cobalto profunda que se fragmenta en cuñas de cian, violeta pálido y plata fría según el ángulo de cada faceta cristalográfica, convirtiendo el espacio en una catedral de prismas apilados a temperatura de menos dos grados. La salmuera hipersalina que te rodea es ligeramente viscosa y lleva una tenue tonalidad ámbar por la materia orgánica disuelta, refractando la luz de manera distinta al agua pura, mientras largas cadenas de *Fragilariopsis cylindrus* —cilindros de sílice de apenas ocho micras— derivan en arcos suaves a través del canal, cada frústulo brillando desde su interior con un oro bruñido cálido donde los cloroplastos cargados de fucoxantina arden como brasas comprimidas contra el azul glacial circundante. Cada cadena está envuelta en una vaina casi invisible de gel EPS anticongelante que difumina los contornos como vaho sobre cristal frío, y sus hebras de mucílago más finas se adhieren a las caras de hielo como hilos de seda. Por debajo, el suelo del canal desaparece bajo una alfombra densa y texturada de biomasa acumulada —resina de ámbar oscuro salpicada de destellos dorados de cloroplastos aún activos— mientras desde arriba los grandes cristales del techo actúan como espejos divisores de haz, lanzando barras diagonales de luz polar tenue que iluminan cadenas individuales con precisión escultórica antes de desvanecerse en la sombra azul-negra de las profundidades del canal.

Desde una posición elevada sobre la válvula inclinada, el observador se enfrenta a una llanura de geometría imposible: miles de hexágonos perfectos se extienden hacia todos los horizontes sin interrupción visible, cada pozo tallado con una precisión que ninguna herramienta humana podría reproducir, sus paredes de sílice opalino captando la iluminación oblicua del haz de electrones hasta convertir cada arista en una línea de porcelana blanca y cada interior en negro absoluto. Este es el esqueleto vítreo de un organismo unicelular —la válvula de un *Coscinodiscus wailesii*— construida en cuestión de horas por vesículas intracelulares que depositan sílice amorfa hidratada (SiO₂·nH₂O) con una fidelidad estructural que desafía toda intuición biológica: los 400 nanómetros de anchura de cada areola y los 200 de profundidad no son tolerancias aproximadas sino dimensiones reproducidas célula tras célula, generación tras generación. Los torreones de los fultoportulae emergen del campo hexagonal como chimeneas industriales huecas, sus tres poros satélite agrupados en la base como arcos de una cripta, mientras que en el margen curvo de la válvula las rimoportulae abren hendiduras oscuras en el precipicio de sílice —estructuras que en vida secretan mucílago y coordinan la flotabilidad de la célula en la columna de agua oceánica. Todo este mundo —esta arquitectura de vidrio biológico capaz de preservarse intacta durante millones de años en el sedimento marino— pertenece a un ser cuya existencia completa transcurre en unas pocas decenas de horas.

Sobre la superficie de una lámina de kelp gigante (*Macrocystis*), extendida como un continente de ámbar y verde oliva bajo la tenue luz filtrada del agua poco profunda, prospera una civilización silenciosa de diatomeas. Las colonias de *Licmophora* se elevan sobre tallos de mucílago como abanicos áureos desplegados hacia la luz, sus células de sílice translúcida —organizadas en arcos flabelados de treinta a sesenta micrómetros de anchura— captando los destellos cambiantes del sol en colores de interferencia: ámbar cálido, cobre y destellos de verde azulado. Entre ellas, las valvas de *Cocconeis* yacen casi invisibles, aplastadas contra la superficie del alga como escudos elípticos de cristal, mientras los ribbons de *Rhabdonema* se tuercen en espirales geométricos que atrapan la luz en sus aristas plateadas. En el plano medio, una cicatriz pálida rasgada en la matriz de polisacáridos extracelulares delata el paso reciente de un anfípodo ramoneador, cuyas mandíbulas aún trabajan en el borde del biofilm, enviando ondas de presión que hacen temblar al unísono todos los abanicos de *Licmophora* como antenas doradas sintonizadas con la misma frecuencia lenta del océano.

Suspendido en el interior de esta cámara arqueada de calcita, el visitante se encuentra rodeado por paredes de un blanco marfil suavemente traslúcido que irradian una luminosidad difusa, como si la luz del fondo marino de arena carbonatada filtrara a través de la propia piedra y convirtiera las paredes en su fuente. El espacio está parcialmente ocupado por un citoplasma pálido y granular —gel vivo de tonos beige grisáceo— del que emergen finos filamentos pseudopodiales que se deslizan hacia el exterior a través de los poros del caparazón como hebras de vidrio estirado, conectando este refugio encerrado con el agua de mar abierta más allá. Distribuidas por toda esa matriz viva descansan las células de *Nitzschia*, cada una una frústula pennada de sílice amorfa de veinte a cuarenta micrómetros de longitud, con sus cloroplastos ricos en fucoxantina irradiando un ámbar dorado intenso —cálido como resina translúcida sostenida ante una llama— mientras permanecen inmóviles en estado simbionte protegido dentro de su huésped foraminífero. Esta relación es una de las asociaciones fotosintéticas más refinadas del océano: el foraminífero *Amphistegina* alberga activamente a estas diatomeas como simbiontes funcionales, aprovechando su fotosíntesis para enriquecer el carbono orgánico disponible en los sedimentos someros iluminados por el sol de las plataformas de carbonato tropical. La atmósfera general es de una quietud filtrada y fecunda: ni corriente, ni turbulencia, solo el brillo suave de la calcita, las constelaciones ámbar de los cloroplastos y la granularidad silenciosa de un mundo interior que sostiene, en calma productiva, a sus huéspedes fotosintéticos.
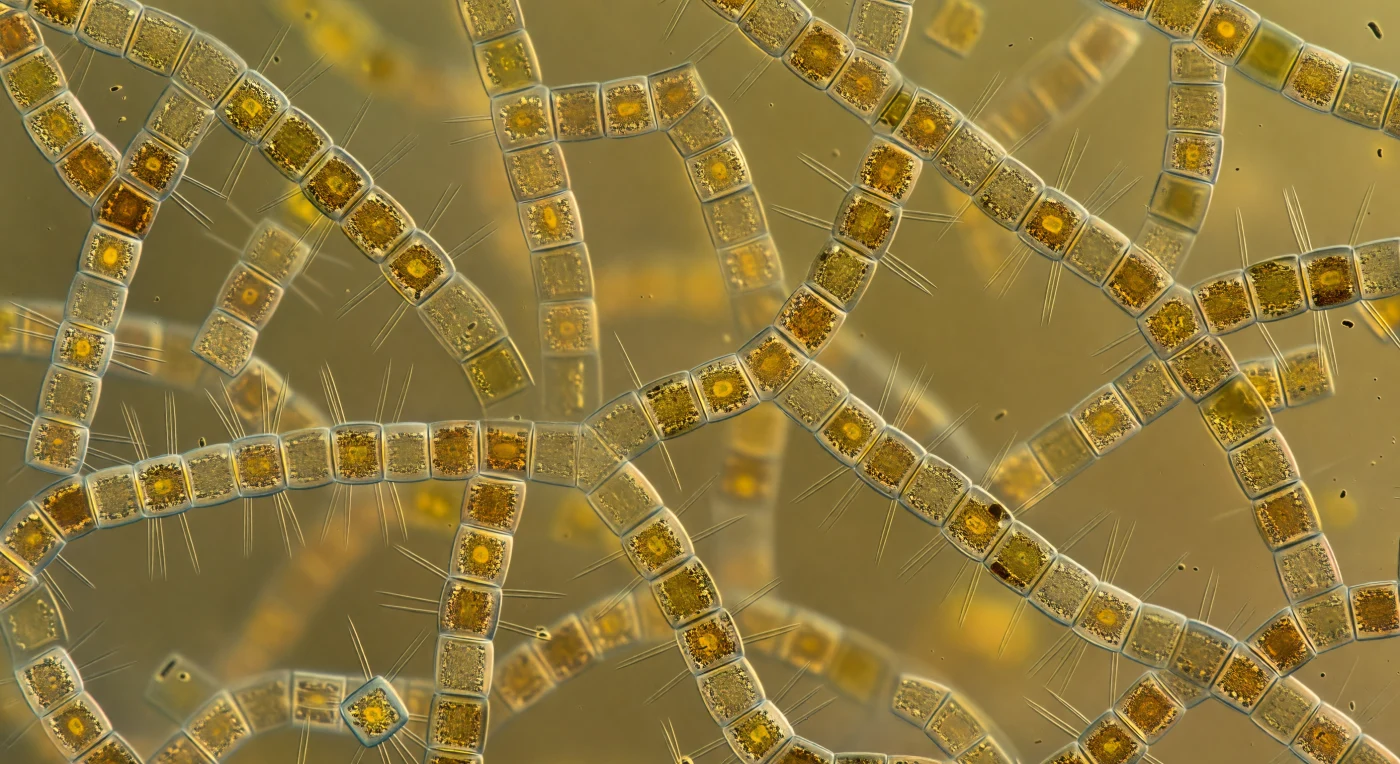
Te encuentras suspendido en el corazón de una catedral ámbar viva, donde cadenas de *Skeletonema costatum* —cilindros de sílice de apenas cinco a ocho micrómetros de diámetro— se extienden en todas las direcciones conectados por varillas fultoportulares tan delgadas como agujas de cristal, entretejiendo una jaula torácica tridimensional sin comienzo ni fin visible. Cada célula arde desde dentro: los cloroplastos cargados de fucoxantina presionan contra las paredes internas de la frústula y convierten la luz solar superficial en una luminosidad ámbar-dorada que no procede de ningún punto concreto sino del medio mismo, fotón a fotón dispersado, absorbido y reemitido por millones de faroles biológicos idénticos hasta que la iluminación llega igual desde todos los ángulos y elimina casi toda sombra. Las cadenas más lejanas —apenas medio milímetro por delante— ya se disuelven en una neblina cálida de oro y oliva oscuro, haciendo que la visibilidad colapse en lo que debería ser agua oceánica abierta. Sobre casi toda superficie cilíndrica crece una capa viva de bacterias en forma de coma y bastón, tan densa que transforma las paredes ópticamente lisas de la sílice en una pelambre de marrón oscuro que absorbe la luz allí donde el vidrio la habría dispersado. El agua entre las cadenas no está vacía sino colmada de polisacáridos extracelulares, pigmento disuelto y partículas virales que espesan el medio hasta hacerlo casi gelatinoso, convirtiendo este florecimiento primaveral en uno de los eventos de productividad más explosivos del océano: en pocas semanas estas células habrán bombeado toneladas de carbono hacia el fondo marino, construyendo su abundancia con silicio disuelto y luz.

Suspendido a escasos centímetros sobre el fondo del Océano Austral, el observador contempla una llanura de tonos gris-beige que se extiende hasta perderse en una neblina de partículas: una acumulación construida durante cien millones de años de floraciones superficiales, depositada en capas de ooze silíceo compuesto casi íntegramente por las estructuras mineralizadas de diatomeas muertas. Desde el sedimento emergen frustulas de *Coscinodiscus* inclinadas a ángulos aleatorios —discos de sílice amorfo hidratado de hasta cien micrómetros de diámetro— cuyas areolas hexagonales, aún perfectamente ordenadas, captan la mínima dispersión de luz disponible y la devuelven como irisaciones fantasmales en aguamarina pálido y marfil frío, la ingeniería de una célula viva conservada como ópalo biogénico durante tiempo geológico. Entre ellos, fragmentos de cadenas de *Eucampia* y valvas pennadas de *Fragilariopsis* muestran márgenes disueltos y estriados apenas sugeridos, consumidos lentamente por el agua abisalmente corrosiva que roza la lisoclina, mientras esferas blancas de caparazones de foraminíferos se desmoronan en manchas calcáreas entre la translucidez vítrea del material silíceo. Desde varios puntos del ooze, colonias bacterianas emiten pulsos de bioluminiscencia azul-fría que ascienden a través de las frustulas superpuestas y las convierten brevemente en ventanales luminosos, y hacia el horizonte brumoso una pluma pálida de sedimento se eleva sin sonido donde un poliqueto excava su camino bajo la superficie, liberando fragmentos de valvas y polvo de carbonato que suben girando lentamente, cada pieza de sílice convertida por un instante en una pequeña linterna fría antes de que la oscuridad lo devore todo.
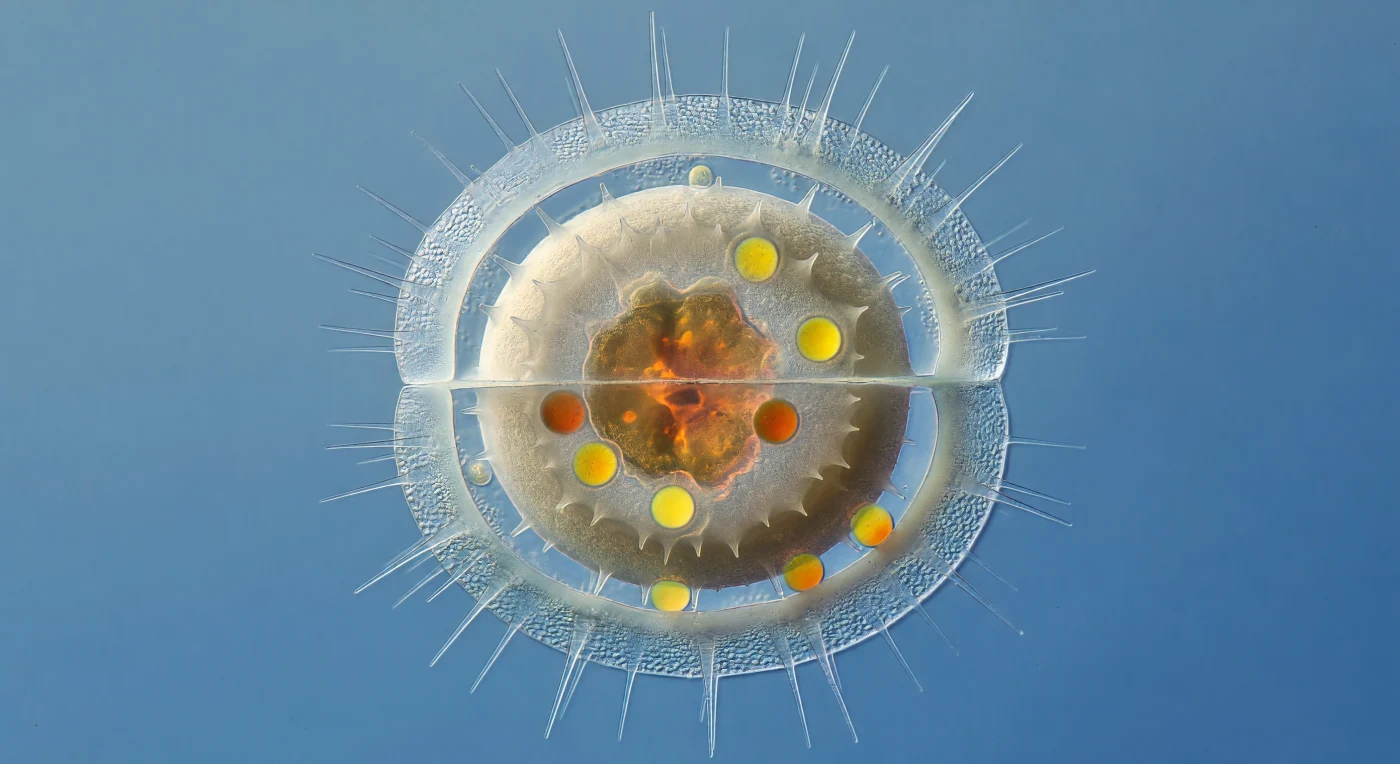
Suspendido a apenas diez micrómetros de la pared exterior de una frústula moribunda de *Chaetoceros*, el observador contempla una de las transformaciones más íntimas del plancton oceánico: el nacimiento de una espora de reposo dentro de la carcasa casi diáfana de la célula madre. Las paredes de sílice de la célula parental se han adelgazado hasta convertirse en láminas de ópalo hidratado de apenas cien nanómetros, translúcidas como escarcha sobre vidrio, mientras las largas setas huecas irradian hacia el agua costera como agujas de cristal que dispersan la luz en halos pálidos. En el interior de este farol desvanecido, la espora de reposo domina el espacio como una piedra dentro de una burbuja de jabón: sus paredes de dos micrómetros de sílice amorfo granular son opacas, de un blanco marfil mate que absorbe los fotones en lugar de doblarlos, en contraste total con la transparencia espectral que la envuelve. Sellada dentro de esa bóveda de cerámica sin vidriar, la masa de cloroplastos colapsada irradia un ámbar profundo y siena quemada —pigmento fucoxantina concentrado al límite— mientras gotas lipídicas esféricas e intensamente refractivas brillan como brasas en amarillo cadmio y naranja oscuro, reservas energéticas que aguardarán meses o años en las frías aguas oceánicas hasta que las condiciones nutricionales permitan a la célula germinar y reanudar su ciclo.

Suspendido en el interior de una pared de diatomita del Mioceno, el observador existe como parte de un archivo mineral que lleva diez millones de años inmóvil: una luz fría y difusa se filtra a través de la propia matriz de sílice, irradiando hacia afuera desde las paredes vítreas de miles de frústulos comprimidos en cada dirección, tiñendo el espacio de un blanco marfil interrumpido únicamente por las sombras gris ceniza que se acumulan en los poros vacíos entre células. Directamente al frente, una columna de valvas de *Melosira* se apila como una torre de monedas pálidas, cada disco de cuarenta micrómetros con su arquitectura de estrías radiales aún resuelta en vidrio fosilizado; a la izquierda, una valva de *Stephanopyxis* ocupa el campo visual como una ventana de catedral, su geometría de lóculos hexagonales visible en sección transversal como una pared de panal de sílice blanca perforada por oscuridades perfectas. Frustulos pennados de *Nitzschia* yacen incrustados en ángulos oblicuos, sus estrías paralelas aún legibles a pesar de la alteración diagenética que ha nublado ligeramente el vidrio sin borrarlo, mientras que cerca del borde inferior un fragmento de escama de pez fósil curva su superficie translúcida como una lente de ámbar, el único destello cálido y orgánico en un mundo por lo demás enteramente mineral. Todo el espacio visible se extiende sin límite como un mosaico geológico denso y pálido, miles de especies identificables congeladas en resolución microscópica, un testimonio silencioso de que la biología y la geología son, a esta escala y en este tiempo, la misma cosa.
