Confianza científica: Alto

Desde el núcleo de sílice amorfa que te envuelve, el mundo se despliega como una catedral imposible: tres esferas concéntricas de ópalo biogénico se extienden hacia afuera, cada una perforada por poros hexagonales de apenas uno a quince micrómetros de diámetro, que actúan como lentes de precisión refractando la luz oceánica en halos prismáticos de violeta pálido y aguamarina cálida, multiplicados en centenares de ventanas curvas superpuestas. Detrás de ti pulsa el endoplasma del núcleo, una luminiscencia color ámbar dorado que irradia calor orgánico contra la geometría fría del azul profundo, mientras doce espinas trirrádeas de sílice atraviesan las tres mallas concéntricas y se pierden en el índigo de la columna de agua como agujas de fibra óptica cuyas puntas se disuelven en la bruma. La propia estructura —secretada por vesículas de depósito silíceo durante horas de lenta acreción mineral— es una obra de ingeniería biomineral que el organismo construyó átomo a átomo en el régimen viscoso donde la inercia no existe y el movimiento browniano sacude cada partícula de manera incesante. Más allá de la esfera exterior, el océano de medianoche se fragmenta en cientos de vistas refractadas, una por cada poro, como si habitaras el ojo compuesto de un organismo mineral que lleva quinientos cuarenta millones de años perfeccionando esta misma geometría en las profundidades del mar.

Suspendido a apenas doscientas micras de su superficie, el observador se enfrenta a una acantaria viva que ocupa el campo visual entero como una linterna joyada en plena oscuridad abisal: veinte espinas de sulfato de estroncio —celestita biológica cristalizada con simetría de Müller, un patrón exclusivo de este grupo entre todos los seres vivos— irradian desde una cápsula central de ámbar oscuro en orden geométrico perfecto, y bajo la luz polarizada cruzada cada varilla ha encendido su propio color de interferencia, cian eléctrico junto a magenta saturado junto a azul cobalto junto a oro, porque la birrefringencia del cristal uniaxial convierte el retardo óptico en longitud de onda visible y ninguna espina comparte el mismo espesor óptico a lo largo de su eje. La cápsula misma aparece casi opaca, calentada por el pigmento de las zooxantelas apretadas contra su pared interior, simbiontes fotosintéticos que ceden carbono al hospedador a cambio de nitrógeno y refugio, y desde esa penumbra tabaco emergen los cables de mionema como hilos tensos y orgánicos que anclan la geometría cristalina en tensión viva. En la periferia de las espinas, los axópodos se pierden casi invisibles en el agua —líneas plateadas de microtúbulos en haz hexagonal que vibran en la corriente browniana y están listos para colapsar en décimas de segundo al contacto de una presa—, recordando que toda esta arquitectura de piedra preciosa y geometría euclidiana no es un mineral sino un organismo que respira, caza y flotará en los sedimentos oceánicos durante millones de años cuando su tejido blando haya desaparecido.

En este instante somos casi nada, suspendidos a escasos centímetros sobre una llanura mineral curva que se extiende hasta un horizonte suave como el de un planeta en miniatura: la pared exterior de sílice de una célula viva de Spumellaria, cuya tésela de ventanas hexagonales y pentagonales cubre todo nuestro campo visual como el suelo de una catedral de vidrio opalino, con los bordes de cada poro levantados en labios luminosos de color ámbar dorado en los travesaños más gruesos y azul grisáceo donde el entramado se adelgaza hasta la casi transparencia. De cada nodo y de cada reborde de poro emerge verticalmente un axopodio rígido —un bosque de lanzas cristalinas de un micrómetro de diámetro que a esta escala dominan el horizonte como columnas de una nave industrial—, sostenidos por haces de microtúbulos internos dispuestos en geometría hexagonal que refractan la luz oblicua hasta encender un filo plateado en un flanco y una costura iridiscente de aguamarina, ámbar y magenta pálido en el opuesto. A lo largo de tres de los axopodios más cercanos, esferas ámbar topacio de tamaño variable —vacuolas alimentarias cargadas de presa digerida y envueltas en membrana— reptan con lentitud deliberada hacia la superficie de sílice, arrastradas por el flujo citoplasmático que recorre el riel invisible de microtúbulos en el interior de cada eje. El fondo absoluto de océano negro, la llanura mineral con su luz interna temblorosa de calymma vacuolado apenas visible a través de los poros, y esas lámparas ámbar desplazándose en silencio componen una arquitectura simultáneamente geológica y viva, congelada en un instante de medianoche que lleva repitiéndose sin variación desde el Cámbrico.

Te encuentras suspendido en el interior de una cámara silícea que curva sus paredes a escasos centímetros en todas las direcciones, construida en ópalo amorfo translúcido que ni es vidrio ni es mineral del todo, sino algo intermedio: lechoso, frágil, vivo en su geometría. La luz del océano penetra por centenares de poros elípticos dispuestos en filas espiraladas sobre la pared latticeada, y cada apertura proyecta hacia el interior un punto de luz aguamarina que tiembla, se superpone con los demás y se desplaza sobre las costillas de sílice como si el techo de una catedral sumergida ardiera con vitrales de agua fría. Sobre tu cabeza, cinco tabiques internos —los septa sucesivos del *Eucyrtidium calvertense*— se apilan en perspectiva forzada, cada uno perforado en su centro por un foramen circular que se estrecha en apariencia hasta convertirse en un destello turquesa puntual, pues cada constricción actúa como una apertura óptica natural que concentra y azulea la luz conforme asciende por la columna de cámaras. La estructura entera es el esqueleto mineralizado de un único protista planctónico cuya sílice fue depositada lentamente, hora tras hora, dentro de vesículas citoplasmáticas, y el filme mucilaginoso que aún recubre los ribetes internos captura la luz lateral con un suave destello ámbar que contrasta con el azul dominante. En el ápice del túnel, silhoueteada en negro perfecto contra el fulgor del océano exterior, la espina apical —un bastón de sílice densa que el organismo secretó como primer andamio estructural— se alza como una aguja de obsidiana hacia las aguas abiertas, último punto de fuga de un mundo arquitectónico construido célula a célula en el plancton del océano profundo.

Te encuentras suspendido a escasos milímetros sobre el fondo de un abismo que no tiene horizonte reconocible, contemplando una llanura gris construida enteramente de esqueletos silíceos: centenares de tests de radiolarios prensados contra arcilla pálida, sus celosías esféricas y cónicas dispuestas como los restos de una ciudad hundida hace millones de años. Un haz oblicuo de electrones rasga la superficie casi en paralelo, convirtiendo cada geometría elevada en un juego brutal de blanco metálico y negro absoluto, donde las perforaciones de cada test proyectan cuadrículas de pozos oscuros tan profundos como las ventanas de una catedral sumergida. Esta llanura es un cementerio de ópalo amorfo, sedimento litogénico y polvo de tests más antiguos aún, acumulado grano a grano durante épocas enteras mientras los organismos muertos descendían desde la zona fótica a razón de unos pocos metros por día hasta quedar sepultados bajo la presión de kilómetros de océano. Lo que la mirada confunde con ruinas arquitectónicas es en realidad el registro geológico más continuo que existe: una necrópolis de precisión matemática depositada sin interrupción desde el Cámbrico, donde cada forma perfecta es la prueba silenciosa de que la vida, en sus escalas más diminutas, construyó estructuras de una complejidad que ninguna erosión ha logrado borrar del todo.
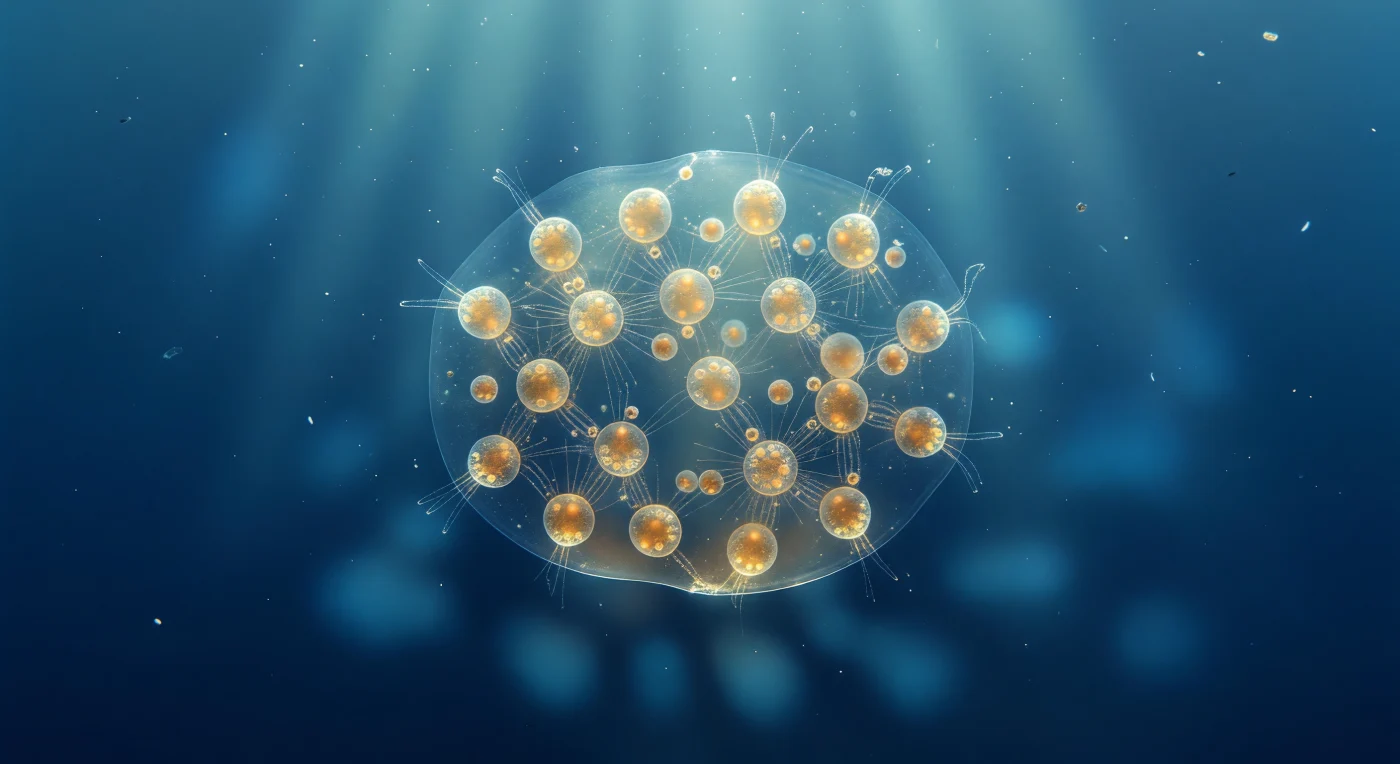
Ante ti se extiende una esfera de vida casi invisible, una lente biológica de ocho milímetros que apenas se distingue del océano que la rodea salvo por los cálidos faroles ámbar de sus células interiores, cada una irradiando su propio halo dorado donde las dinoflageladas simbiontes absorben la luz tropical que desciende desde arriba en anchas columnas acuamarinas. La colonia de Sphaerozoum flota en perfecta inmovilidad a diez metros de profundidad, sostenida por sus vacuolas lipídicas como una nave anclada en el azul, su mucílago gelatinoso tan transparente que la frontera entre organismo y océano no es más que un ligero estremecimiento óptico, un aumento imperceptible de viscosidad donde los filamentos reticulopodiales más externos atrapan bacterias y copos orgánicos que derivan desde el agua abierta. Desde cada célula parten axópodos como agujas de cristal —estructuras sustentadas por haces de microtúbulos en disposición geométrica precisa— que se vuelven invisibles hasta que una columna de luz los atrapa en el ángulo exacto y los transforma en agujas de plata que se entrecruzan por todo el interior gelatinoso, formando una red filamentosa que conecta blandamente a decenas de individuos en un único organismo colonial sin esqueleto de sílice. La colonia actúa como una lente débil: la luz que la atraviesa se refracta en su superficie curva y proyecta hacia abajo un patrón de cáusticas cambiantes, una danza de claroscuros azules que se desvanece hacia el añil y luego hacia el índigo sin fondo, como si este organismo transparente y autoluminoso fuera una vidriera de catedral a la deriva en una silencio azul e infinito.
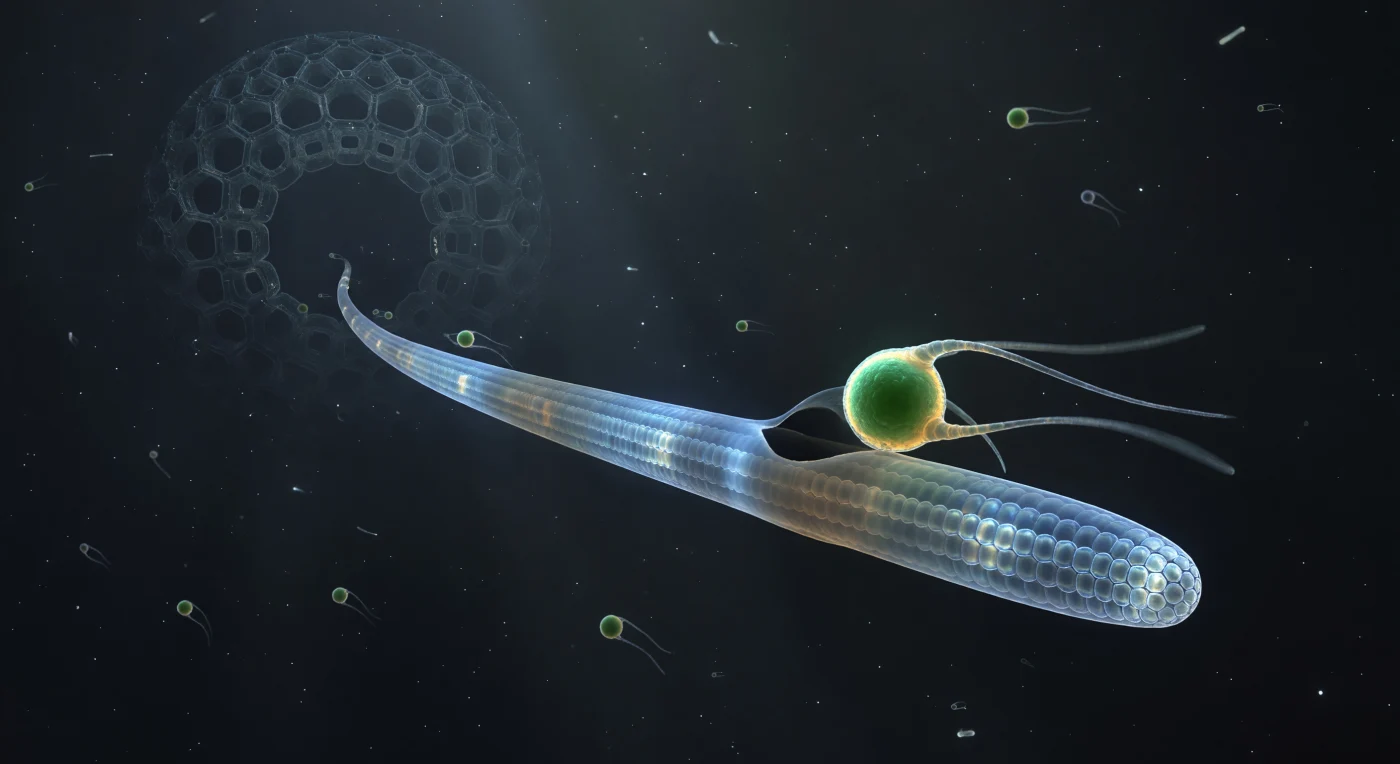
Suspendido en una oscuridad casi absoluta, el observador se encuentra a centímetros aparentes de un acontecimiento que en realidad ocurre a escala de micras: la superficie de un axopodio radiolario —una lanza traslúcida cuya arquitectura interna de microtúbulos paralelos proyecta halos prismáticos a lo largo de toda su extensión— acaba de recibir el contacto de un flagelado Micromonas de intenso verde esmeralda, sus dos flagelos todavía suspendidos en arcos luminosos como filamentos de luz detenida. En el punto exacto de encuentro, la membrana del axopodio muestra una leve deformación hacia adentro, y desde esa hendidura emerge un creciente oscuro —tan delgado que parece una sombra más que una estructura material— comenzando su primer grado de arco alrededor de la presa, mientras el citoplasma circundante vira de un azul vítreo y frío a un gris-ámbar turbio que delata la cascada bioquímica ya desencadenada. Más profundo a lo largo del axopodio, hacia la oscuridad catedralicia de la cápsula central, formas vesiculares a la deriva —comidas anteriores— han perdido sus pigmentos vivos y flotan ahora como linternas de color óxido en una corriente imperceptible, testimonio silencioso de que este momento de equilibrio entre presa libre y presa capturada es, en la escala del tiempo de la célula, fugacísimo. Al fondo, apenas resuelto, el entramado silíceo del caparazón del radiolario se alza como las ruinas de una catedral alienígena, sus aperturas hexagonales retroiluminadas por la dispersión bioluminiscente de la calymma, recordando que toda esta violencia microscópica ocurre dentro de una arquitectura mineral de una regularidad que la razón confunde con intención.
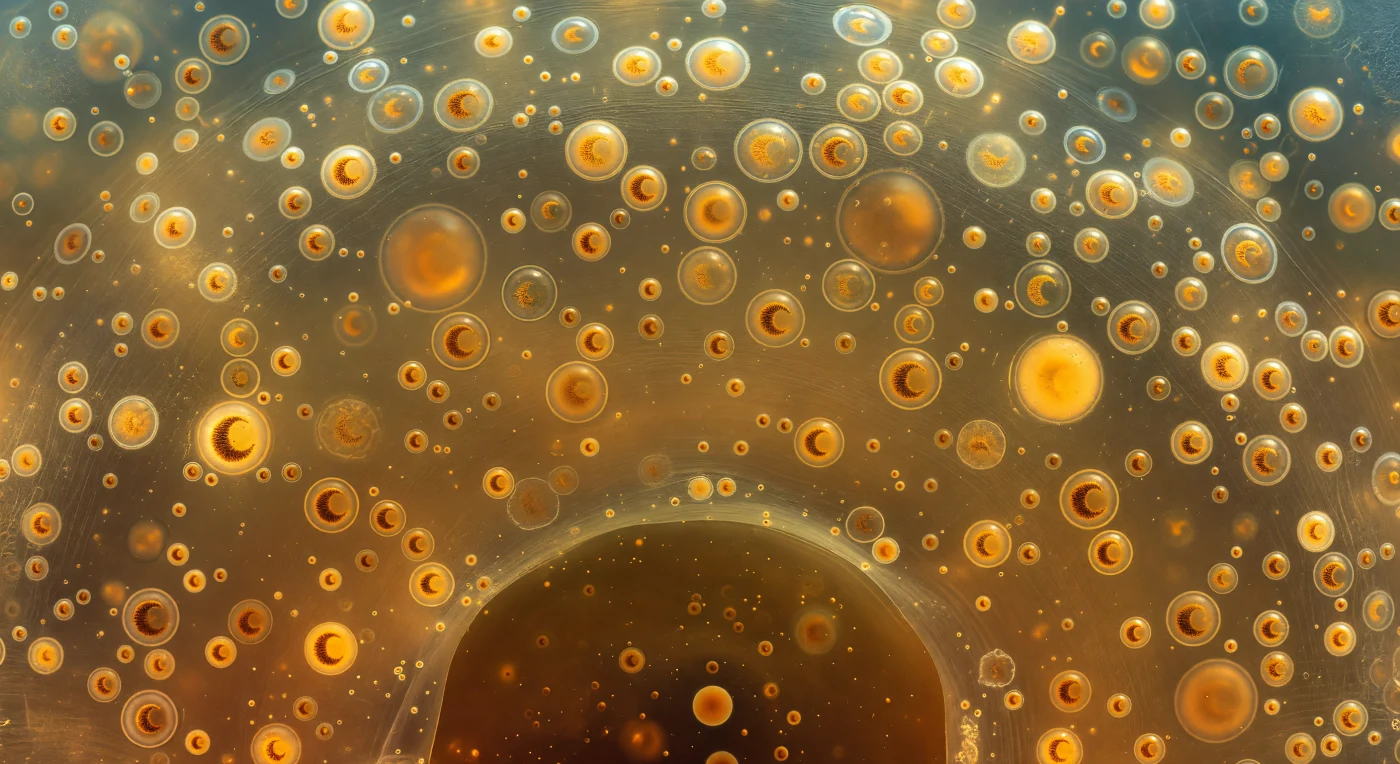
En todas las direcciones, a distancias que van desde unos pocos cuerpos hasta el límite de lo visible, flotan las zooxanthellae —esferas dorado-ambarinas de diez a quince micrómetros cada una, tan numerosas que llenan el campo visual sin llegar a tocarse, como faroles suspendidos en el aire quieto antes de una celebración. Dentro de las más cercanas, el núcleo en forma de C se curva como una luna creciente, un arco de ocre oscuro y granulado perfectamente legible a esta proximidad, recordando que cada una de estas esferas es una célula viva completa, una dinoflagelada simbiótica atrapada en el interior del calymma de un Colodario colonial, donde convierte la luz solar en nutrientes para su hospedador a cambio de refugio y minerales. El ectoplasma que todo lo sostiene —ese gel translúcido que tiembla entre lo líquido y lo sólido— no está vacío: una red apenas resuelta de filamentos de glicoproteínas lo atraviesa como escarcha sobre cristal, visible solo donde la luz ámbar la roza en ángulo rasante y la convierte en un encaje de plata. Las vacuolas lipídicas se elevan entre los simbiontes como esferas de cristal de treinta o cuarenta micrómetros, doblando la luz en cáusticas doradas que iluminan lo que flota detrás de ellas, actuando a la vez como depósitos de flotabilidad y como lentes que magnifican suavemente las células más distantes. Más allá de esta primera generación de zooxanthellae claramente resueltas, la colonia se espesa en una bruma cálida y dorada que absorbe la profundidad como el terciopelo absorbe la luz, hasta que el horizonte del calymma se disuelve en el contorno curvo y oscuro de la cápsula central —una membrana orgánica perforada que sella el endoplasma como la pared de un mundo interior, un santuario celular que late con quietud en la penumbra ámbar del océano abierto.
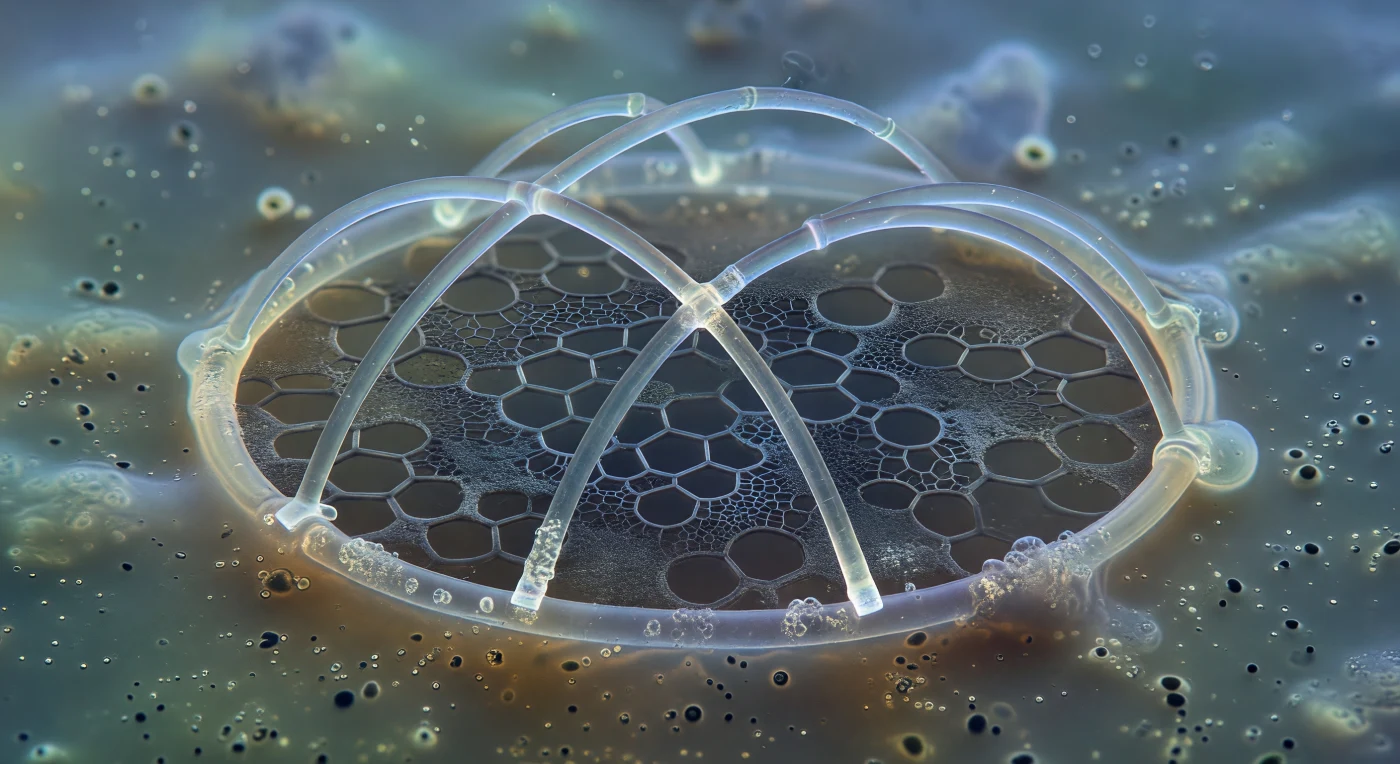
Flotas en suspensión absoluta dentro de un medio denso y gelatinoso, rodeado por la penumbra cálida del citoplasma de un radiolario vivo, cuando ante ti emerge la estructura más antigua del organismo: el anillo sagital de un *Pterocorys* en sus primeras horas de existencia, una arcada de sílice amorfa apenas depositada que brilla con reflejos blanco-dorados y halos iridiscentes donde la óptica de contraste de fase acaricia sus bordes curvos. Dos barras primarias cruzan el interior del anillo en ángulos precisos, sus terminaciones romas y recién formadas, rodeadas por el fulgor tenue de vesículas de deposición silícea — pequeñas fábricas moleculares que bombean ácido silícico desde el agua de mar hacia una arquitectura que tardará horas en completarse. Detrás de este andamiaje mineral, casi invisible, se despliega la plantilla proteica orgánica: una malla de glicoproteínas de color gris carbón que no emite luz sino que la absorbe, trazando ya con precisión asombrosa la geometría hexagonal de poros que aún no existen en mineral, un mapa dibujado en materia blanda para guiar lo que será duro. El medio circundante es un coloide ámbar y translúcido poblado por gotículas lipídicas esféricas y mitocondrias que tiemblan con el movimiento browniano, todo sumergido en un resplandor acuático de azul-verde difuso que se pierde progresivamente en la neblina vacuolada de la calymma, mientras la sílice que ahora se deposita, con paciencia mineral indiferente a la vida que la fabrica, sobrevivirá intacta en el registro fósil durante quinientos millones de años.

Te encuentras aplastado contra la cara interior de una vesícula de deposición de sílice, una pared que se curva sobre ti con la autoridad de un casco de fundición iluminado en ámbar, mientras la membrana silicalema se extiende en lo alto como una doble línea de oscuridad absoluta trazada con precisión quirúrgica sobre una luminosidad crema que difunde desde más allá. Ante ti avanza el frente de gel de sílice amorfa —una masa gris-blanca, semitransparente, casi pétrea en su lentitud— encaballando el andamiaje de proteínas ricas en silafinas como la marea que oscurece la arena bajo su peso, polimerizando el mineral alrededor de cada fibrilla terracota hasta convertirla en siena profundo, sepultándola con la paciencia geológica de una lava que enfría. El proceso que aquí ocurre es la biominealización en acto: las proteínas silafina actúan como plantillas electrostáticas que concentran el ácido ortosilícico disuelto y aceleran su condensación en redes de sílice hidratada, construyendo en horas las paredes del esqueleto que tardará millones de años en fosilizarse en el sedimento oceánico. Más allá de la vesícula, el citoplasma se hace presente como presencias de color —un glóbulo azul cobalto, una gota amarillo-ámbar, un gránulo verde esmeralda suspendido en la distancia traslúcida— recordándote que esta fábrica mineral latida y viva ocurre dentro de una célula única, invisible a simple vista, a la deriva en pleno océano abierto.

El suelo bajo tus pies es una llanura curva de sílice gris ceniza, pulida hasta un acabado casi vítreo, una armadura de ópalo amorfo tejida por el propio organismo molécula a molécula desde aguas abisales, y su superficie se extiende en una teselación hexagonal tan densa que la mayor parte de lo que pisas es vacío: cada poro es una abertura de quince micrómetros flanqueada por labios de sílice levantados que se encuentran en nodos nodulares, y el interior de cada uno se hunde en una negrura absoluta que ninguna luz alcanza. El horizonte cae con una curvatura suave pero inconfundible en todas las direcciones, recordándote que habitas la superficie de algo esférico y finito — una esfera de apenas doscientas micras de diámetro, una luna mineral cuya circunferencia completa no supera el grosor de un cabello humano. Por encima de ti, seis espinas triradiadas de sílice extruida se alzan como obeliscos a alturas equivalentes a rascacielos en esta escala, sus bases fusionadas con los nodos de la celosía y sus puntas perdiéndose cuarenta o cincuenta micrómetros arriba, construidas no por ningún proceso geológico sino por la maquinaria biológica de un protozoo marino que depositó este esqueleto en cuestión de horas dentro de vesículas citoplasmáticas especializadas. La iluminación raking del microscopio electrónico de barrido aplana todo color hasta un espectro de grises fríos — plata viva en las aristas iluminadas, carbón profundo en las sombras de los poros — y convierte lo que en el océano sería una estructura translúcida flotando entre bacterias y diatomeas en algo que parece una ruina geológica, silenciosa, perfecta e indiferente.

Suspendido en la columna mesopelágioca a quinientos metros de profundidad, el observador flota rodeado de una procesión silenciosa de testas nassellarias que descienden en todas direcciones como catedrales de cristal a la deriva: formas cónicas, pagodas escalonadas y cámaras apiladas de entre cien y cuatrocientos micrómetros de diámetro, cada una construida en sílice amorfa opalina cuya retícula translúcida fragmenta la última luz azul-índigo que se filtra desde la superficie en destellos fríos a lo largo de cada poro y cada barra radial. Las testas habitadas arden desde dentro con un resplandor ámbar y ocre, la célula viva presionando su citoplasma contra las paredes de la celosía como vitrales biológicos que emiten calor orgánico en medio de un entorno de frío mineral; las testas vacías, en cambio, capturan esa misma luz sin devolverle ninguna calidez, sus cámaras huecas rellenas solo de agua de mar y sus matrices de poros tan abiertas y oscuras como cuencas en un cráneo de vidrio hilado. Entre estas formas deriva la nieve marina, agregados mucosos y orgánicos semiluminosos que ruedan en espirales lentas y se enrollan ocasionalmente en una espina saliente antes de desprenderse y continuar su descenso. El conjunto compone un universo de complejidad geométrica extraordinaria, frágil y antiquísimo a la vez, suspendido en una oscuridad casi total atravesada únicamente por ámbar biológico y plata mineral.

En este universo mineral detenido en el tiempo, la mirada atraviesa un mosaico denso e interminable de formas esféricas y cónicas recrializadas en cuarzo microcristalino y calcedonia, los esqueletos silíceos de radiolarios planctónicos que habitaron un océano jurásico hace ciento cincuenta millones de años y que hoy yacen comprimidos en una roca de apenas treinta micrómetros de espesor. La luz transmitida sube fría y pareja desde abajo, difundiéndose a través de la masa compacta de tests deformados por el peso de kilómetros de sedimento, confiriéndoles ese resplandor interior sordo y fosfórico, como si la piedra misma recordara haber sido vida transparente en aguas abisales. Manchas de hematita en óxido de hierro —rojo sangre seca, naranja herrumbre— sangran por los intersticios que antes fueron porosidad rellena de agua marina y que la diagénesis transformó en cemento mineral teñido de tiempo. La geometría aplasta: domos aplastados sobre conos truncados sobre esferas soldadas entre sí por presión litostática, cada test un fantasma arquitectónico donde la trama hexagonal de poros originales apenas sobrevive como relieve espectral en la superficie recritalizada. Todo es simultáneamente íntimo y planetario, el archivo comprimido de un océano que no existe, legible solo a través de la luz polarizada y el silencio de la roca.

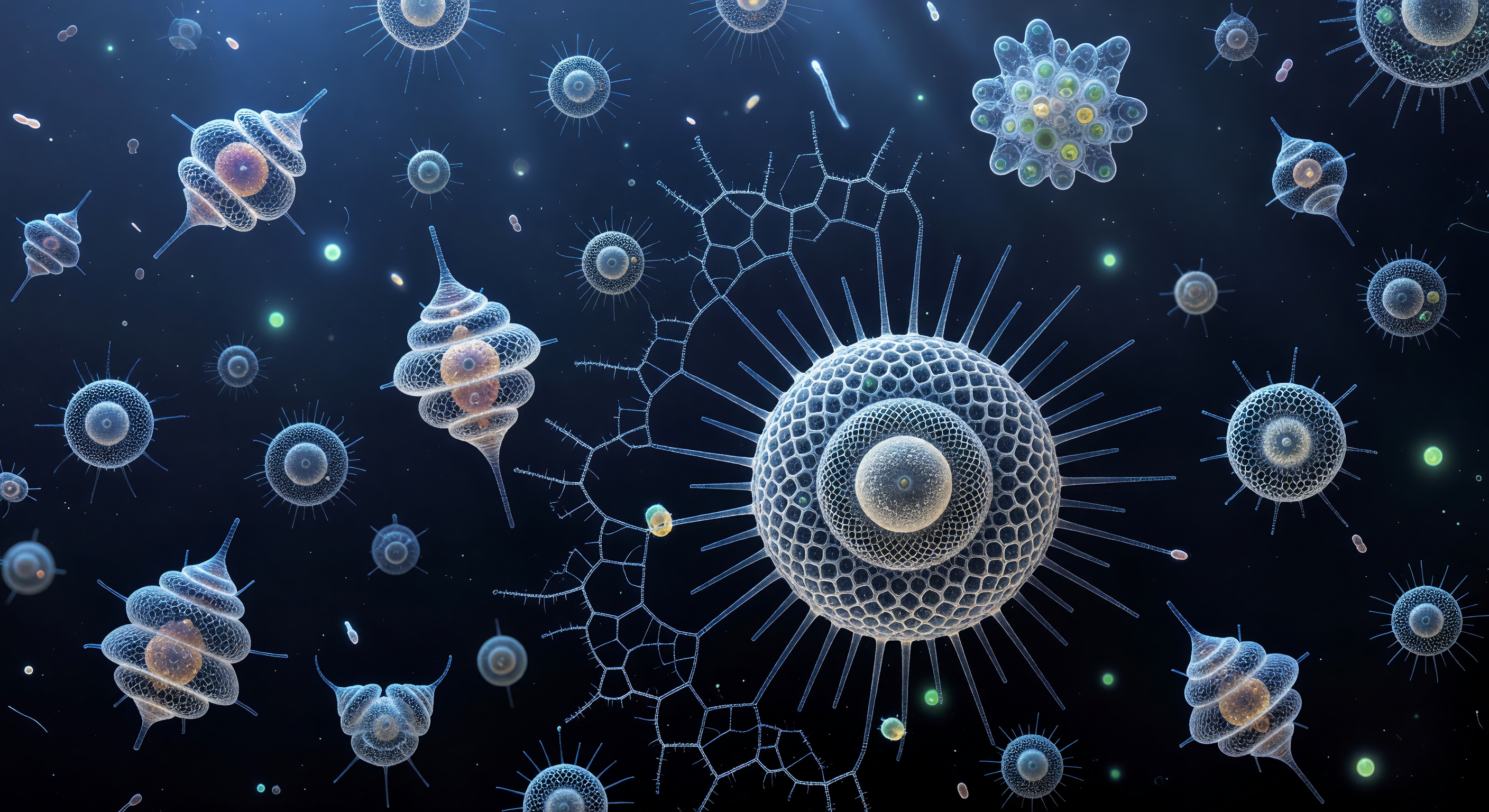
Flotando en una oscuridad absoluta que el cerebro confunde instintivamente con el espacio interestelar, el observador se encuentra rodeado de explosiones cromáticas suspendidas: las acantarias, cada una sostenida por veinte espinas de sulfato de estroncio dispuestas según una geometría icosaédrica, irradian colores de interferencia puros bajo la luz polarizada cruzada, convirtiendo la iluminación en destellos de magenta, cobalto, amarillo sulfuro y violeta eléctrico mediante birrefringencia, el mismo fenómeno óptico por el que los cristales transforman la luz polarizada en espectro visible. Entre estas explosiones de color, las pruebas de sílice amorfa de las esponjas esferoidales —las Spumellaria— flotan como arquitectura fantasmal en tonos de peltre frío, su sílice opalo incapaz de manipular la luz polarizada y por ello condenada a permanecer como jaulas de celosía hexagonal perfectamente articuladas pero espectralmente silenciosas, esferas concéntricas unidas por puntales radiales que evocan catedrales góticas conservadas en niebla metálica. El campo de dos milímetros que contiene este universo alberga decenas de organismos vivos distribuidos a profundidades variables, los más lejanos reducidos a puntos de joya pura hundiéndose en la distancia negra, mientras los más próximos extienden axópodos invisibles —filamentos citoplasmáticos sostenidos por haces de microtúbulos cristalinos— que rozarían al observador con la misma delicadeza con que una telaraña de luz podría capturar una bacteria a la deriva. Esta escena, comprimida sobre un portaobjetos de vidrio, condensa quinientos cuarenta millones de años de registro fósil continuo en un instante de fuego mineral y geometría silenciosa.

En la oscuridad absoluta de dos mil metros de profundidad, comprimir tu existencia a quinientas micras de agua de mar salada significa habitar un universo donde la presión es un sólido invisible y la luz no existe salvo la que tú mismo traes: el cono azul-blanco de tu foco submarino corta el agua sin color y allí, suspendida en el vacío que devora toda geometría, aparece *Aulacantha scolymantha*, sus quinientas espinas radiales de sílice hueco abiertos en todas las direcciones como la explosión detenida de una araña de cristal, cada tubo de apenas dos micras de diámetro conduciendo tu haz hacia dentro y devolviendo en sus extremos distales un destello frío de fibra óptica, una constelación esférica de coordenadas azul-verdosas dispersas en la nada. En el centro del organismo el faeodio ocupa el núcleo como una herida oscura, una masa opaca de material parcialmente digerido de tono marrón oscuro que bloquea por completo la luz transmitida y confiere al ser ese peso interior inquietante, mientras la malla silícea de la cápsula central apenas se vuelve legible como hilos de plata fugaces en el haz, presencia deducida más que vista. Más allá del halo iluminado, la oscuridad no es ausencia sino material, una sustancia que el luz tantea y no logra penetrar, y el organismo permanece en su universo privado de geometría fría, temblando levemente en las puntas de sus espinas por el roce browniano del agua, respirando el frío de las profundidades como si la vibración misma fuera su única señal de vida.
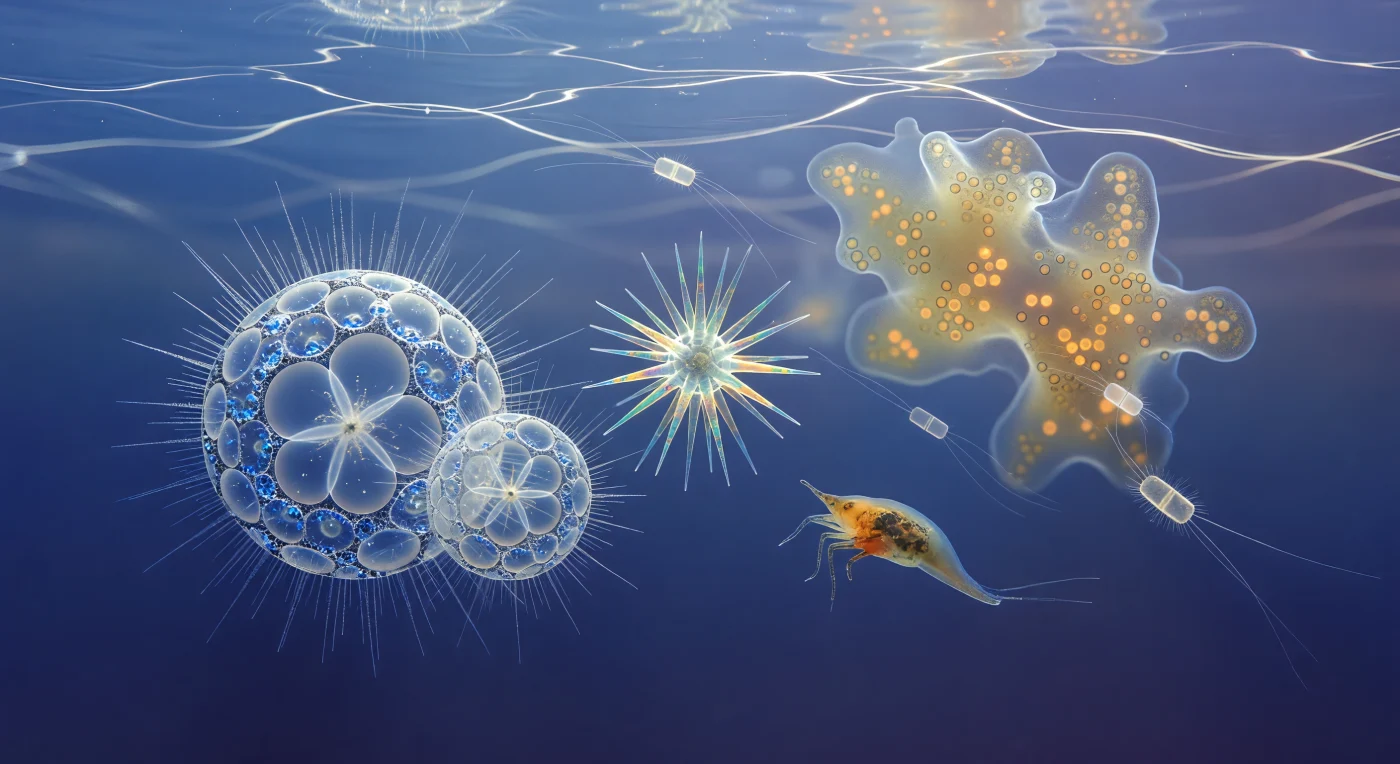
Suspendido a treinta metros bajo la superficie del Pacífico tropical, el mundo a esta escala no tiene horizonte reconocible: solo un vacío azul-índigo luminoso que se extiende en todas direcciones, atravesado desde arriba por redes de luz cáustica que oscilan como caligrafía blanca en movimiento perpetuo, cada filamento refractado por las olas de la superficie antes de descender suavemente hasta acariciar las estructuras colosales que te rodean. A tu izquierda, dos individuos de *Sphaerozoum* se elevan como catedrales de vidrio hilado, sus retículas de sílice amorfa —ópalo biogénico— anidadas en esferas concéntricas conectadas por barras radiales que brillan con una blancura lechosa al rozar la luz, mientras su corona de axópodos, filamentos tan finos como agujas de cristal sostenidos internamente por haces de microtúbulos en geometría hexagonal, destella brevemente en plata viva cada vez que una red de luz cáustica los atraviesa. Más atrás, una estrella de *Acantharia* gira con lentitud glacial, sus veinte espinas de sulfato de estroncio —mineral que no precipita en ninguna otra célula conocida, ensamblado en simetría icosaédrica codificada genéticamente— cambiando entre aguamarina eléctrico, ámbar dorado y rosa interferencial a medida que la luz las atraviesa con birrefringencia. La colonia de Collodaria que ocupa tu periferia superior es ya tan vasta que semeja una nebulosa ámbar: una matriz gelatinosa compartida por centenares de células, encendida desde dentro por miles de dinoflagelados simbióticos —sus cloroplastos transformando fotones en carbono orgánico a contraluz del sol superficial— mientras las diatomeas *Chaetoceros* derivan entre todo esto como lanzaderas de vidrio esmerilado arrastrando setas de sílice tan delgadas que solo existen cuando la luz las elige. En este régimen de Stokes donde la viscosidad domina y la inercia carece de significado, cada organismo flota suspendido en una sopa azul translúcida cargada de bacterias, orgánicos disueltos y el pulso invisible de la cadena trófica microbiana que sostiene los océanos desde el Cámbrico.