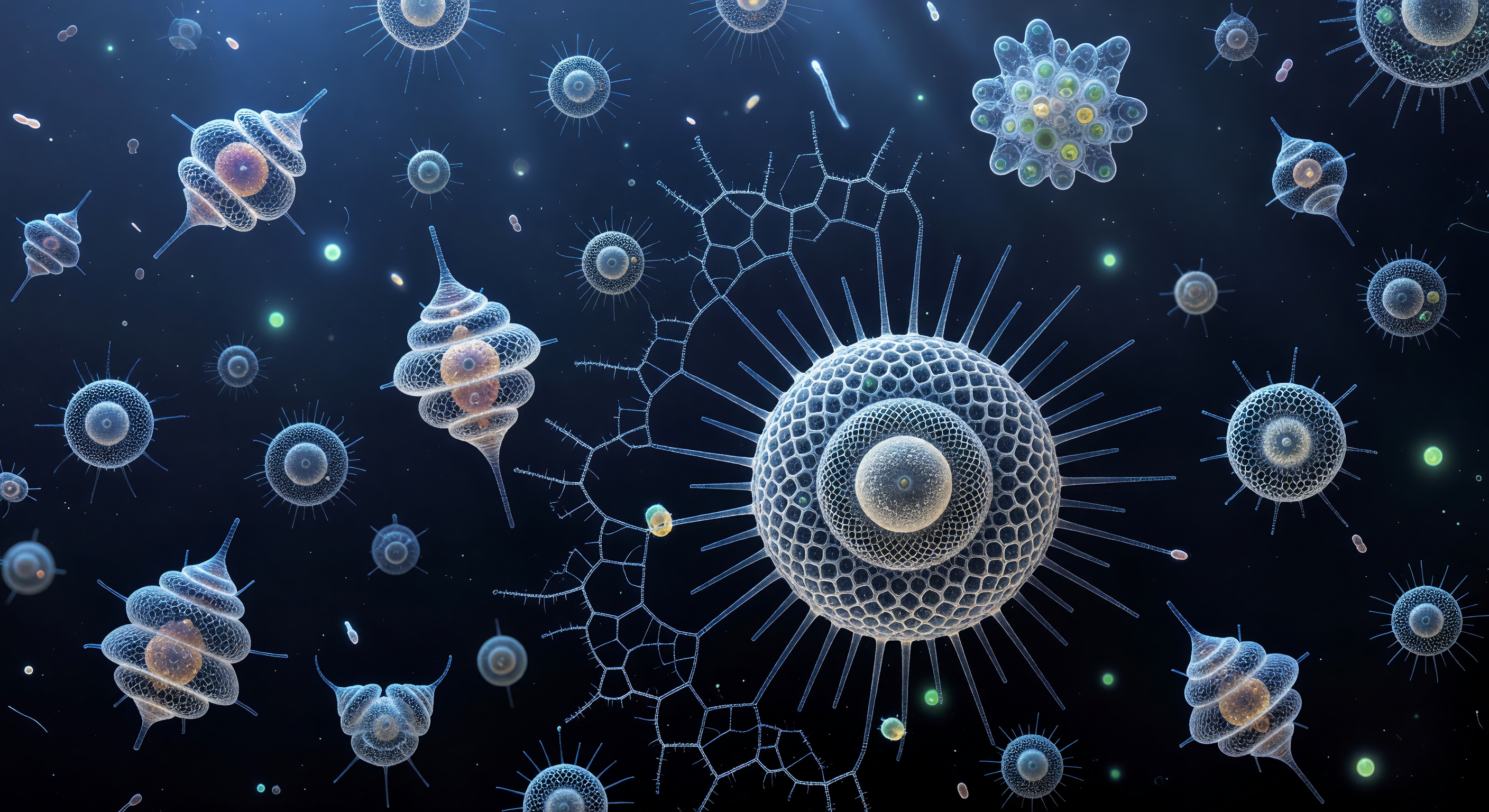
Affidabilità scientifica: Alto

Sospesi al cuore geometrico di *Actinomma asteracanthion*, lo sguardo si apre verso l'esterno attraverso tre sfere concentriche di silice opale, ciascuna traforata da pori esagonali larghi pochi micrometri che agiscono come lenti in miniatura, rifrangendo la luce blu-verde dell'oceano profondo in aloni prismatici — violetto pallido ai margini, acquamarina al centro — fino a trasformare l'intera struttura in una cattedrale di vetro smerigliato e luce viva. Dodici spine triradiate si irradiano dalla sfera più interna, attraversano le due lattice successive e si perdono nell'indaco del mare aperto come aghi di cristallo che trasmettono per tutta la loro lunghezza un barlume di luminescenza, simili a fibre ottiche forgiate dalla biomineralizzazione. Alle spalle, o meglio tutt'attorno al punto da cui si guarda, il nucleo irradia un calore ambrato — l'endoplasma denso e granulare che pulsa dolcemente, addolcendo con la sua luce mielata la freddezza blu della geometria minerale che lo circonda. Il test di silice, depositato dall'organismo stesso attraverso vescicole intracellulari in ore di lenta accrezione minerale, è al tempo stesso scheletro, filtro ottico e trappola per la luce: ogni poro inquadra una porzione diversa dell'abisso esterno, moltiplicando la prospettiva come l'occhio composto di un impossibile organismo minerale. Al di là dell'ultima sfera, l'oceano si risolve in un blu volumetrico attraversato da fotoni diffusi e da una foschia di particolato — batteri, frustuli di diatomee, organici disciolti — che galleggia nella quiete viscosa di un mondo dove la gravità conta meno del moto browniano e il tempo scorre alla velocità della mineralizzazione.

Sospeso a questa distanza nell'oscurità dell'oceano aperto, l'osservatore si trova davanti a una delle architetture più straordinarie del mondo vivente: un'Acantaria, radiolario marino la cui struttura è governata dalla legge di Müller — venti spine di solfato di stronzio (celestite, SrSO₄) disposte secondo la simmetria perfetta di un icosaedro, ciascuna un monocristallo puro che si estende dal nucleo centrale come i raggi di una cattedrale organica. Sotto la luce polarizzata incrociata simulata, ogni spina trasforma la propria birifrangenza cristallina in colore puro: la differenza di percorso ottico tra i due raggi polarizzati che attraversano il reticolo cristallino produce interferenze distinte — ciano elettrico, magenta satura, blu cobalto, oro — colori che non sono pigmenti ma geometria della luce stessa, ciascuno dipendente dall'orientazione del cristallo rispetto ai polarizzatori. Al centro, la capsula centrale è un ovale caldo e opaco, affollata di zooxanthellae simbionti — alghe dinoflagellate da cinque a quindici micrometri ciascuna — che trasformano la luce in carbonio e ossigeno, tingendo la parete capsulare di ruggine e ambra. Tra le spine incandescenti, i cavi mionemici — strutture contrattili proteiche uniche agli Acantaria — corrono come fili tesi nell'ombra, capaci di muovere le spine coordinatamente per regolare la profondità di galleggiamento della cellula; all'estrema periferia, gli assopoidi si prolungano nell'acqua come fili d'argento appena percettibili, i loro assonemi di microtubuli vibranti nel moto browniano del mezzo, tesi e pronti a catturare qualunque particella di cibo che la corrente porti a contatto.

Ci troviamo sospesi a pochi centimetri sopra una pianura minerale curva che si estende in ogni direzione come il pavimento di un pianeta in miniatura — la parete reticolare esterna di una cellula di Spumellaria viva, costruita in silice opalina e percorsa da una tessellazione perfetta di finestre esagonali e pentagonali i cui bordi sporgenti catturano la luce obliqua DIC come argento fuso, mentre le loro cavità restano nere come abisso marino, lasciando intravedere appena il calymma vacuolato che palpita al di sotto. Da ogni orlo dei pori e da ogni nodo reticolare si erge verticalmente un assopodio rigido — un bosco fitto di lance cristalline del diametro di circa un micron, ma che a questa scala appaiono come colonne di cattedrale, le loro superfici vetrose striatamente illuminate su un fianco di bianco-argento e orlate sul lato opposto da una sottile frangia di interferenza DIC in turchese, ambra e magenta pallido, come cuciture al neon che percorrono l'intera lunghezza del fusto. Lungo tre degli assopodi più vicini scivolano lentamente verso la superficie sfere ambrate di vacuoli alimentari — involucri membranosi densi e rifrangenti che ardono come lanterne topazio nel buio circostante, trascinati verso l'interno dallo streaming citoplasmatico lungo il fascio di microtubuli interni a una velocità di uno o pochi micron al secondo, e la loro differenza di indice di rifrazione distorce appena il liquido intorno, producendo un lieve effetto di lente che tradisce la loro presenza nell'oscurità. Il freddo assoluto dell'oceano aperto, la geometria minerale sotto i piedi, la foresta di cristallo sopra la testa e quelle lente luci ambrate che migrano in silenzio compongono insieme un paesaggio che è al tempo stesso ecosistema, architettura e cosmo minerale.

Sospesi all'interno della camera più profonda di *Eucyrtidium calvertense*, ci troviamo avvolti da una volta di opale siliceo amorfo che si curva intorno a noi come la navata di una cattedrale scolpita nell'acqua marina solidificata — le pareti reticolate, a portata di mano in ogni direzione, punteggiate da pori ellittici disposti in file spiralate precise, attraverso i quali l'oceano aperto irradia verso l'interno come una luce acquamarina intensa e mobile, proiettando aureole blu-verdi tremolanti che si sovrappongono sulle nervature silicee e trasformano l'intera superficie in un mosaico vivente di luce rifratta. Il materiale stesso non è né vetro né cristallo ma qualcosa di intermedio — traslucido, leggermente lattescente, con un film mucillaginoso che riveste i listelli interni e dove il citoplasma ha lasciato residui lungo le travi, la silice brilla di un tono ambrato caldo che contrasta con il blu dominante, testimonianza della continua attività biologica di un eucariota unicellulare il cui scheletro è prodotto per precipitazione minerale guidata da vescicole di deposizione intracellulari. Alzando lo sguardo, cinque setti interni si sovrappongono in convergenza forzata — ciascuno un anello traslucido perforato al centro da un singolo forame circolare, le aperture che si riducono progressivamente fino a diventare un punto di luce turchese brillante, con ogni costrizione che agisce come un diaframma naturale concentrando e blueshiftando i fotoni oceanici in risalita. All'apice, sagomata in nero perfetto contro il blu abbacinante dell'esterno, la spina apicale si proietta come un ago d'ossidiana — un singolo bastoncino di silice densa il cui bordo cattura un sottile orletto bianco di rifrazione prima di dissolversi nella colonna d'acqua aperta sopra di noi, unico punto di connessione tra questo spazio minerale e il vasto oceano pelagico che lo contiene.

Ci si trova sospesi a pochi millimetri sopra un paesaggio lunare e privo di colore, dove centinaia di scheletri silicea di radiolari — organismi unicellulari planctonici vissuti e morti nella colonna d'acqua oceanica — giacciono compattati in un'argilla grigia sul fondo degli abissi, formando quello che i sedimentologi chiamano *radiolarian ooze*, un sedimento biogenico che si accumula a profondità superiori ai quattro chilometri dove il carbonato di calcio si dissolve ma la silice amorfa sopravvive. Il fascio di elettroni obliquo rivela con spietata chiarezza ogni geometria: test sferici di Spumellaria ancora intatti con le loro triplici sfere concentriche forate — una cattedrale geodetica in cui ogni poro misura appena qualche micron — accanto a forme coniche di Nassellaria rovesciate, le loro camere segmentate schiacciate come torri di raffreddamento collassate, mentre spine cave proiettano ombre nere assolute sul sedimento circostante. Tra questi monumenti geometrici, la matrice argillosa riempie gli spazi come neve silenziosa, seppellendo parzialmente forme integre e lasciando emergere altre su piedistalli di materiale compresso, testimonianza dei processi diagenetici e della pressione litostatica accumulata nel corso di milioni di anni. Non esiste colore in questo paesaggio — solo il contrasto fisico tra silice illuminata dagli elettroni e vuoto di ombra — eppure ogni superficie rivela la propria verità materiale: le microfratture nei gusci sotto compressione geologica, i granuli di silt incastrati nei pori come detriti nelle finestre di una città sommersa, un cimitero matematico di organismi che hanno costruito architetture perfette per vivere, e poi le hanno lasciate precipitare nell'oscurità dell'oceano profondo.
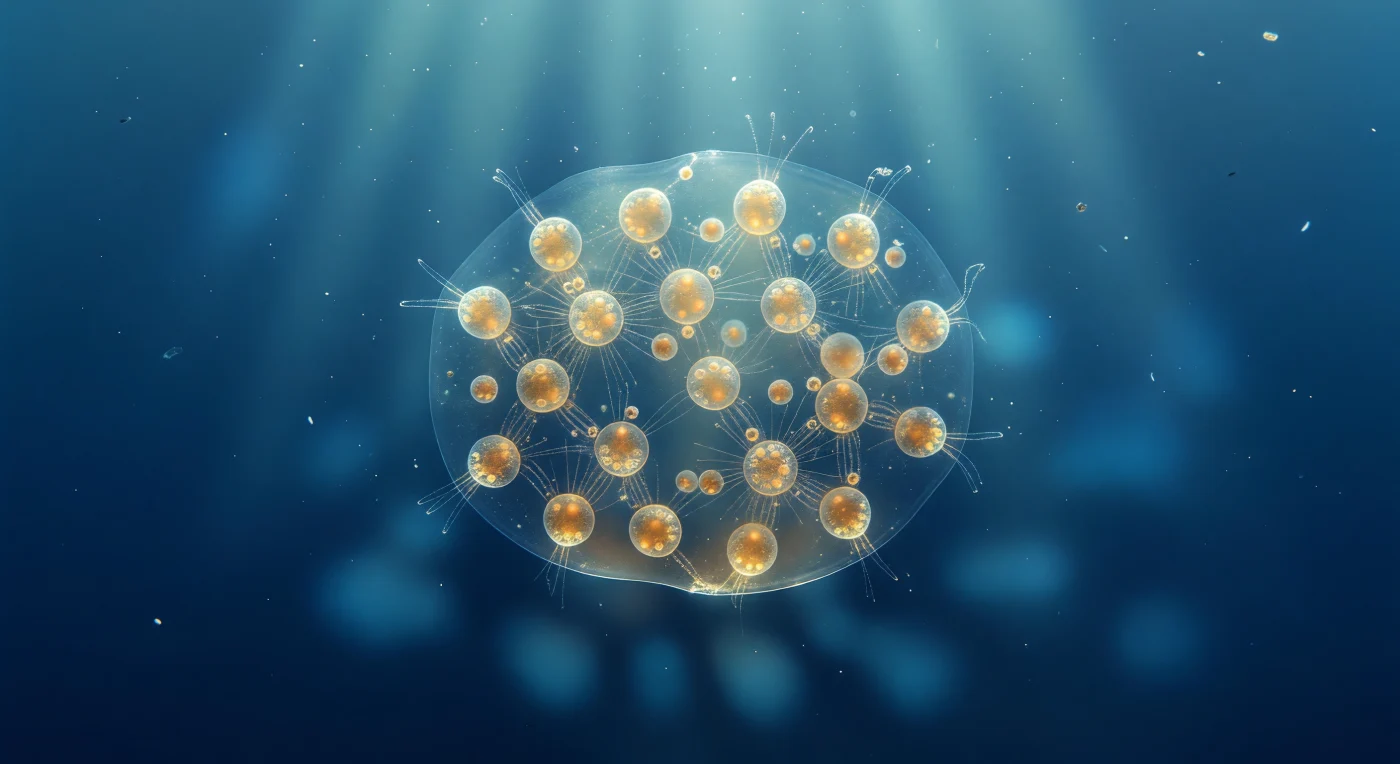
Sospesi a un millimetro dalla superficie di questa zattera coloniale di Sphaerozoum, l'intera colonia riempie il campo visivo come un pianeta gelatinoso che fluttua nel vuoto azzurro — una lente ellissoidale di otto millimetri di mucillagine quasi perfettamente trasparente, il cui confine con l'oceano circostante è segnalato soltanto da un impercettibile tremolìo ottico. All'interno di questa massa traslucida, decine di cellule individuali brillano come lanterne d'ambra distribuite nel gel, ciascuna colorata dal denso carico di dinoflagellati simbiotici — zooxantelle intrappolate nell'ectoplasmico calymma — che trasformano la luce tropicale discendente da acquamarina a miele caldo e ocra. Dai bordi di ogni cellula si irradiano assopodi sottilissimi, filamenti cristallini sostenuti da fasci di microtubuli in geometria esagonale, quasi invisibili finché un raggio radente non li illumina trasformandoli in aghi d'argento che si estendono fino a cento micron nel gel circostante, intrecciandosi con quelli dei vicini in una rete filamentosa che percorre l'intera colonia senza alcun supporto scheletrico di silice. L'assenza di un guscio mineralizzato — caratteristica distintiva dei Collodaria rispetto alle spumellarie e nassellarie con il loro elaborato reticolo di selce — lascia la colonia libera di oscillare come un'entità puramente morbida, la sua galleggiabilità mantenuta dai vacuoli lipidici che scintillano come perle opalescenti dentro ogni organismo, mentre sotto la zattera il sole proiettato attraverso la mucillagine curva disegna un arabesco di caustica blu in lenta deriva verso le profondità indaco.
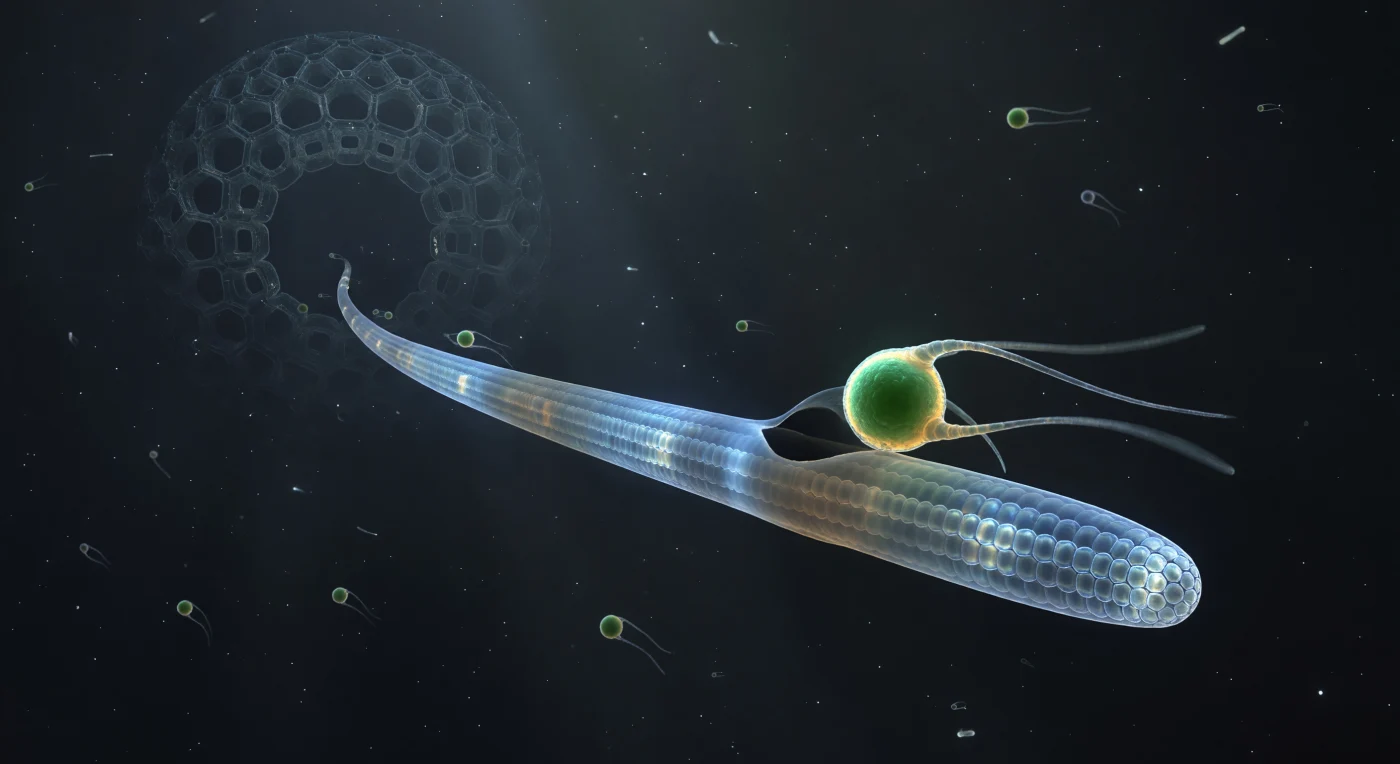
Ci si trova sospesi nell'oscurità dell'oceano aperto a una scala in cui i confini tra predatore e preda si rivelano come architettura molecolare: un assopodio di radiolario si estende come una lancia di vetro traslucido, il suo interno percorso da fasci di microtubuli disposti in sezioni trasversali esagonali che catturano e diffrangono i fotoni in aloni prismatici, trasformando l'intera struttura in una fonte di luce fredda e geometrica. Contro questa precisione cristallina, un flagellato di Micromonas — una sfera densa di verde smeraldo intenso, cloropasti che bruciano di autofluorescenza — emerge come un piccolo pianeta ardente nel momento esatto del contatto, i suoi due flagelli bloccati in archi di filamento luminoso, il moto browniano ai loro bordi l'unico segno del mondo cinetico che li circonda. Il dramma si concentra nel punto di giunzione: la parete vitrea dell'assopodio mostra una minima inflessione, e da quella fossetta una sottile falce di nuova membrana ha cominciato a estendersi attorno alla cellula verde come un'eclissi al suo primo grado d'arco, mentre il citoplasma circostante vira dal blu ghiaccio a un grigio ambrato torbido, segnale della cascata biochimica che converte un organismo libero in un vacuolo digestivo. Più in profondità lungo l'assopodio, verso l'oscurità della capsula centrale, forme vescicolari derivano lentamente come lanterne — pasti precedenti i cui pigmenti si sono spenti in ruggine e ocra — e sullo sfondo, appena risolto, il reticolo siliceo del test radiolario si erge come una cattedrale aliena con aperture esagonali retroilluminate dal tenue scatter bioluminescente, ogni poro una finestra buia incorniciata da travi di silice così regolari da sembrare architettura piuttosto che biologia.
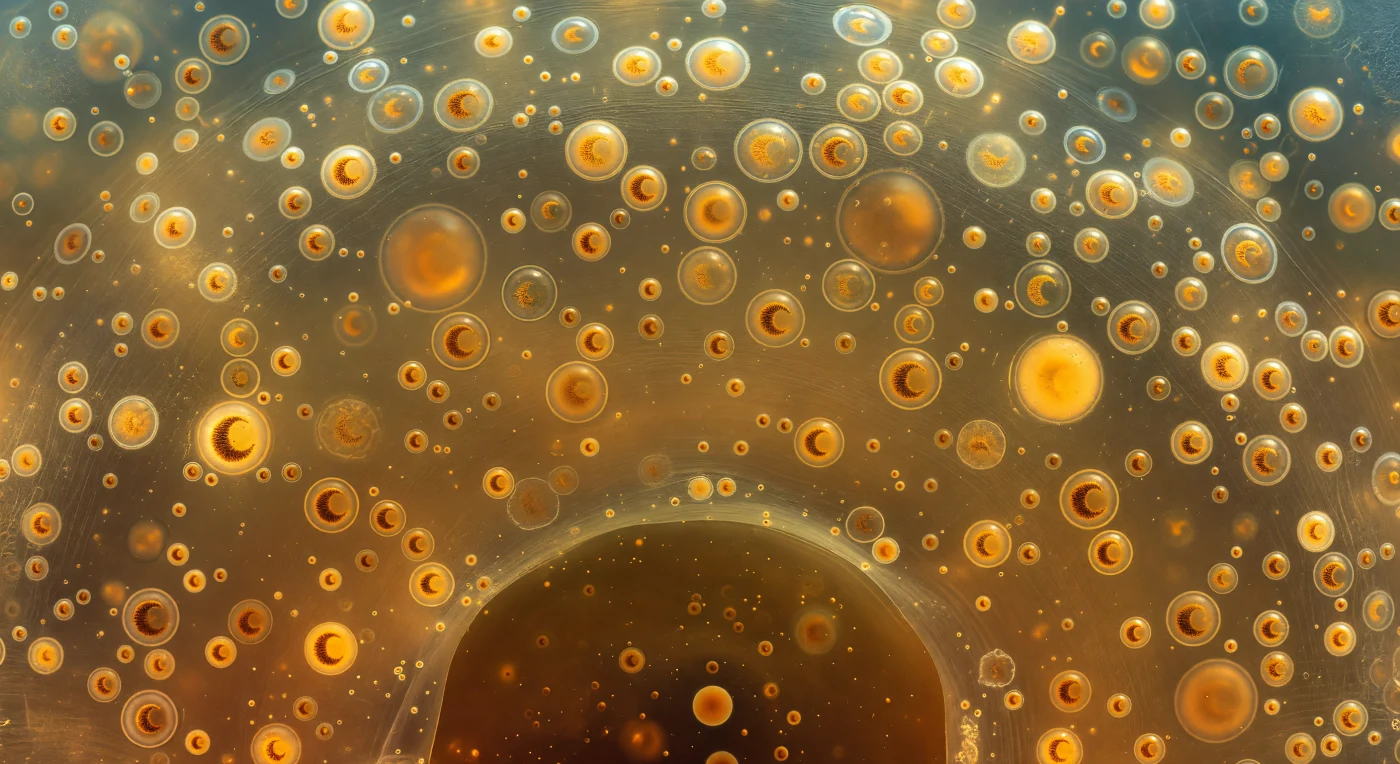
Sospesi all'interno della calymma di un Collodariano coloniale, il mondo intorno a voi non è né liquido né solido ma qualcosa di intermedio — un gel organico di straordinaria trasparenza, caldo come ambra riscaldata, che preme contro ogni senso di confine con un'intimità totale e silenziosa. In ogni direzione fluttuano le zooxantelle, sfere di dieci-quindici micrometri ciascuna, dorate e ambrate, così numerose da affollare il campo visivo senza mai toccarsi, come lanterne sospese nell'aria ferma prima di una festa; nelle più vicine si distingue con perfetta chiarezza il nucleo a forma di mezzaluna, un arco denso e granulare di ocra-bruna che curva all'interno del cortex dorato come una falce impressa nel caramello. I vacuoli lipidici si gonfiano a distanze ravvicinate come sfere di vetro trasparente del tutto prive di colore, trenta o quaranta micrometri di diametro, che curvano la luce ambientale in un brillante riflesso a mezzaluna sul lato esposto e proiettano un caustico dorato concentrato sulle zooxantelle che fluttuano dietro di loro; questi vacuoli non sono semplici strutture di riserva, ma organi di galleggiamento che regolano con precisione la posizione verticale dell'organismo nella colonna d'acqua, consentendo alla colonia di restare sospesa nella zona fotica dove la simbiotica algale può catturare luce. La luce stessa — filtrata dall'oceano aperto sopra, schermata da strati e strati di cellule e vacuoli — non arriva da una direzione precisa ma sembra emanare dal mezzo stesso, una luminescenza ambrata diffusa che trasforma ogni struttura in qualcosa che pare illuminato dall'interno, e il glicoproteico reticolo dell'ectoplasma diventa visibile solo ai margini, là dove la luce lo sfiora di taglio e lo trasforma per un istante in un lacework d'argento prima di dissolversi nuovamente nell'invisibile.
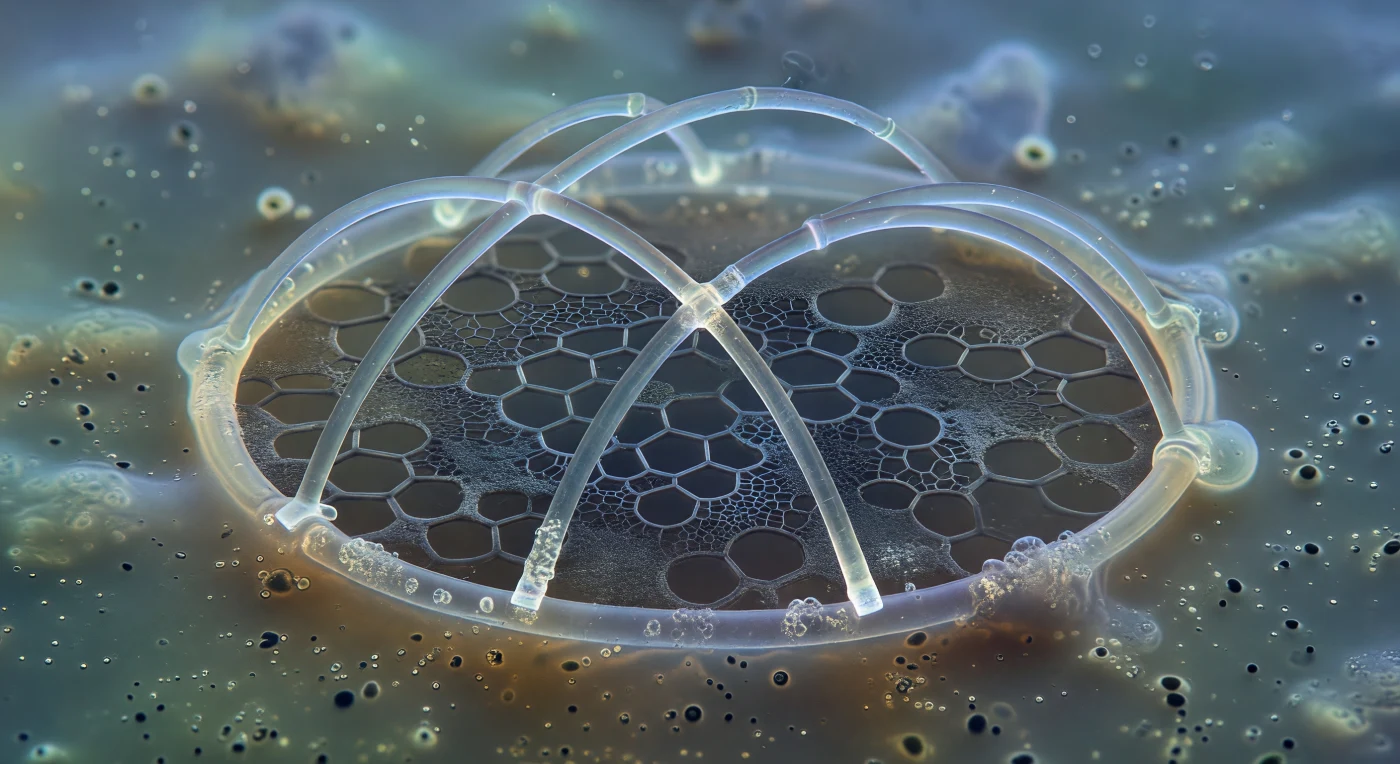
Nell'oscurità lattea di questo mare interiore, sospesa nel cuore liquido di una cellula viva, una struttura si erge davanti a te con la solennità silenziosa di una cattedrale incompiuta: l'anello sagittale di un *Pterocorys* nascente, appena deposto in silice trasparente come borosilicato filato, i suoi archi luminosi orlati da aloni iridescenti dove il contrasto di fase accarezza le superfici curve. Due barre primarie attraversano l'interno dell'anello ad angoli precisi, con le estremità ancora ottuse e fresche, come chiavi di volta non ancora inserite in un arco che non esiste ancora. Dietro al minerale, appena percettibile come brina su vetro scuro, si distende la trama proteica organica che precede e guida la mineralizzazione — un reticolo grigio-cenere di fibre glicoproteiche che già disegna con geometria infallibile il contorno di pori vuoti, esagoni inscritti nella materia organica morbida, in attesa che la silice li raggiunga. Il citoplasma tutt'intorno è un mezzo colloide ambrato e granuloso, percorso dal fremito browniano di goccioline lipidiche e mitocondri indistinti, tutto avvolto in un'acquamarina diffusa che dissolve ogni profondità in nebbia vacuolata. Ciò che guardi — questa struttura di pochi micron di spessore — legge come un'opera architettonica colossale dalla tua prospettiva, e il minerale già depositato brilla con la permanenza paziente di qualcosa destinato a sopravvivere al suo costruttore per cinquecentomilioni di anni.

Sei schiacciato contro la parete interna di una vescicola di deposizione silicea, una superficie curva che si estende come lo scafo interno di una fonderia sospesa nell'ambra, illuminata da una luce diffusa e lattea che filtra attraverso la silicalemma sopra di te — una doppia linea di assoluta oscurità che segue un arco lentissimo, come il bordo di un pianeta visto da vicino. Sul pavimento davanti a te avanza il fronte del gel di silice, una massa grigio-pallida e semitrasparente che progredisce con la pazienza geologica della lava che si raffredda, i suoi margini sfumati in un reticolo di fibrille proteiche color terracotta — le proteine silaffin-simili — che formano un'impalcatura biologica tra il tessuto e l'architettura minerale. Dove il gel è più spesso diventa quasi opaco, e alle sue frange traslucide si vedono le fibre sottostanti scurirsi dal rosso mattone al siena profondo man mano che la silice polimerizza attorno a loro, trasformando struttura organica in minerale eterno con la stessa lentezza con cui il mare avanza sulla sabbia bagnata. Oltre la parete della vescicola, nell'ambra del citoplasma circostante, globuli luminosi — un blu cobalto lontano, un verde smeraldo sospeso come una lampada dietro vetro smerigliato — ricordano che questo spazio chiuso e silenzioso, saturo di calore minerale, è il cuore pulsante di una cellula viva.

Ci si trova in piedi su una pianura curva di silice color cenere pallida, levigata quasi fino alla lucentezza del vetro borsiliicato smerigliato, la sua superficie appena granulare alla scala del nanometro poiché il materiale è opale amorfo, non cristallino — lo scheletro siliceo di un Hexacontium, un radiolario marino vissuto in acque pelagiche aperte. La tassellatura di pori esagonali si estende in ogni direzione fino a un orizzonte visibilmente curvo, ogni apertura un pozzo nero e assoluto di circa quindici micrometri di diametro, i suoi bordi rialzati in labbra di silice precise che si incontrano ai nodi della reticella in apofisi nodulari che catturano la luce radente come perle di vetro fuse — una geometria ottimizzata da centinaia di milioni di anni di evoluzione per filtrare l'acqua marina, intrappolare le prede e fluttuare nella colonna d'acqua oceanica grazie alla struttura ultraleggera e porosa. Sei spine triradiate si elevano dalla reticella come obelischi monumentali, ciascuna una struttura di silice estrusa che sorge dalla fusione con i nodi della lattice fino a una punta d'ago quaranta o cinquanta micrometri più in alto — al corpo del radiolario, la loro funzione è idrodinamica e difensiva, ma in questa finzione del microscopio elettronico a scansione, svuotata dell'acqua di mare che normalmente la circonda, la loro geometria appare puramente minerale e aliena, come monoliti su un satellite deserto. Sotto i pori, appena visibile come una tessitura scura attraverso il vuoto delle aperture, si intravede la sfera interna concenrica della stessa cellula — un'altra shell di silice con i propri pori esagonali sfasati rispetto a quelli esterni, promemoria della complessità architettonica di un organismo unicellulare costruito alla scala di una frazione di millimetro.

Ci si trova sospesi a cinquecento metri di profondità, immersi in una colonna d'acqua così fredda e densa da sembrare quasi solida, mentre intorno a noi scende una processione silenziosa di architetture minerali: i test nassellariani — forme coniche, campane sovrapposte, pagode allungate in opale amorfo — ciascuno tra i cento e i quattrocento micrometri nella realtà, eppure a questa scala cattedrali di vetro che scivolano lente davanti al viso come reliquiari in caduta libera attraverso un oceano senza fondo. Le reticelle di silice sono traslucide e debolmente iridescenti, e dove ancora abita una cellula viva la citoplasma ambrata e ocra preme contro la parete interna della gabbia geometrica, filtrando calore biologico attraverso ogni poro come luce colorata attraverso un rosone medievale, mentre i test vuoti — e sono molti — restituiscono soltanto un argento freddo, le loro camere colme di sola acqua marina, le matrici di pori aperte come orbite nel cranio di un essere fatto di vetro filato. Tra questi corpi in discesa fluttuano fiocchi di neve marina — aggregati di muco trasparente, materia organica morta, citoplasma perduto — che turbinano in spirali lente e a volte si avvolgono delicatamente intorno a uno sperone siliceo prima di liberarsi, ognuno che cattura la bioluminescenza diffusa nell'acqua e la restituisce come un barlume perlaceo nel registro cromatico dominante di indaco, ardesia e argento glaciale, con isole di ambra dove la vita ancora resiste alla deriva verso il fondo.

Dentro una sezione sottile di radiolarite giurassica, illuminate da una luce polarizzata che risale dal basso come un freddo bagliore lunare, le forme spettrali di milioni di scheletri planctonici riempiono ogni centimetro del campo visivo — cupole sferiche, coni nassellari compressi, ovali deformati dalla pressione litostatica di centocinquanta milioni di anni, tutti ricristallizzati in calcedonio e quarzo microcristallino nei toni dell'avorio spento, del grigio cenere e del bianco osseo. Ciò che un tempo erano test silicei traslucidi galleggianti negli abissi di un oceano scomparso — il Tetide o un suo predecessore — sono oggi fantasmi minerali saldati insieme in una massa compatta, le loro originali reticole esagonali di pori appena visibili come lievi rilievi superficiali, impressioni di un'architettura biologica quasi cancellata. Tra le forme si insinua la macchiatura a ossido di ferro: il rosso ruggine e il cremisi del sangue secco colano attraverso il cemento interstiziale, tracciando in negativo gli spazi porali che un tempo erano acqua di mare, ora mineralizzati in venature color ocra e ambra che sembrano scorrer ancora, lentissime, nella pietra. La luce trasmessa diffonde verso l'alto attraverso l'intera massa compressa, conferendo a ogni superficie un bagliore interiore fosforescente e discreto, come se il tempo geologico stesso emettesse luce piuttosto che assorbirla. Si ha la sensazione di stare immersi in una nebbia fatta di plancton fossilizzato, sospesi dentro l'archivio compresso di un fondale oceanico che non esiste più.

Nell'oscurità assoluta creata dai polarizzatori incrociati, ci si trova sospesi in quello che sembra uno spazio interstellare compresso su un vetrino da microscopio: intorno a te, in ogni direzione, le Acantaria esplodono come fuochi d'artificio rallentati, i loro venti spine di solfato di stronzio — un minerale birifrangente raro nel mondo biologico — che convertono la luce polarizzata in interferenze cromatiche pure, ogni cellula che brucia in una firma spettrale diversa, magenta e oro, blu elettrico e viola profondo, giallo zolfo e rosso cadmio. Queste spine si irradiano dalla capsula centrale con una simmetria icosaedrica imposta geneticamente, una geometria che la selezione evolutiva ha mantenuto invariata per centinaia di milioni di anni, e i filamenti di mioneme che le collegano al corpo cellulare appaiono come cavi luminosi tesi in un nodo di forze viventi. Tra queste esplosioni cromatiche, i gusci delle Spumellaria derivano come architetture fantasma color peltro — la loro silice opale amorfa otticamente inerte ai polarizzatori, eppure strutturalmente maestosa: sfere concentriche di reticolo esagonale, cattedrali gotiche in miniatura sospese nel buio, alcune così prossime che le loro axopodi impalpabili sfiorerebbero la tua pelle. L'intero campo di due millimetri si legge come una volta celeste abitata — cinquanta organismi distribuiti in un volume che occupi corporalmente, ciascuno un universo separato di mineralogia e citoplasma vivente.

Nell'oscurità assoluta di duemila metri di profondità, il fascio del proiettore taglia l'acqua come una lama di vetro freddo, e lì, al margine del nulla, *Aulacantha scolymantha* emerge dalla tenebra come un'esplosione congelata: cinquecento spicole cave di silice si irradiano in ogni direzione, captando la luce e restituendola alle punte distali come fredde fibre ottiche, ciascuna un fantasma azzurro-verde sospeso nel vuoto, l'insieme una costellazione sferica di meno di un millimetro che galleggia in uno spazio che rifiuta di essere misurato. Al centro, il feodio incombe come una contusione viva — una massa opaca di materiale digerito e organelli che blocca ogni luce trasmessa, mentre attorno ad esso la sottile rete della capsula silicea si rivela solo per lampi argentati, un'architettura deducibile più dall'inferenza che dalla visione diretta. Il calimma gelatinoso, costellato di vacuoli microscopici, diffonde il fascio in un'aureola di luminescenza diffusa tra il nucleo scuro e il campo di spine radianti, e l'acqua stessa, a questa scala, ha una texture di irrequietezza termica che fa tremare le punte più esterne delle spicole come se l'organismo stesse respirando il freddo del mondo in cui vive.
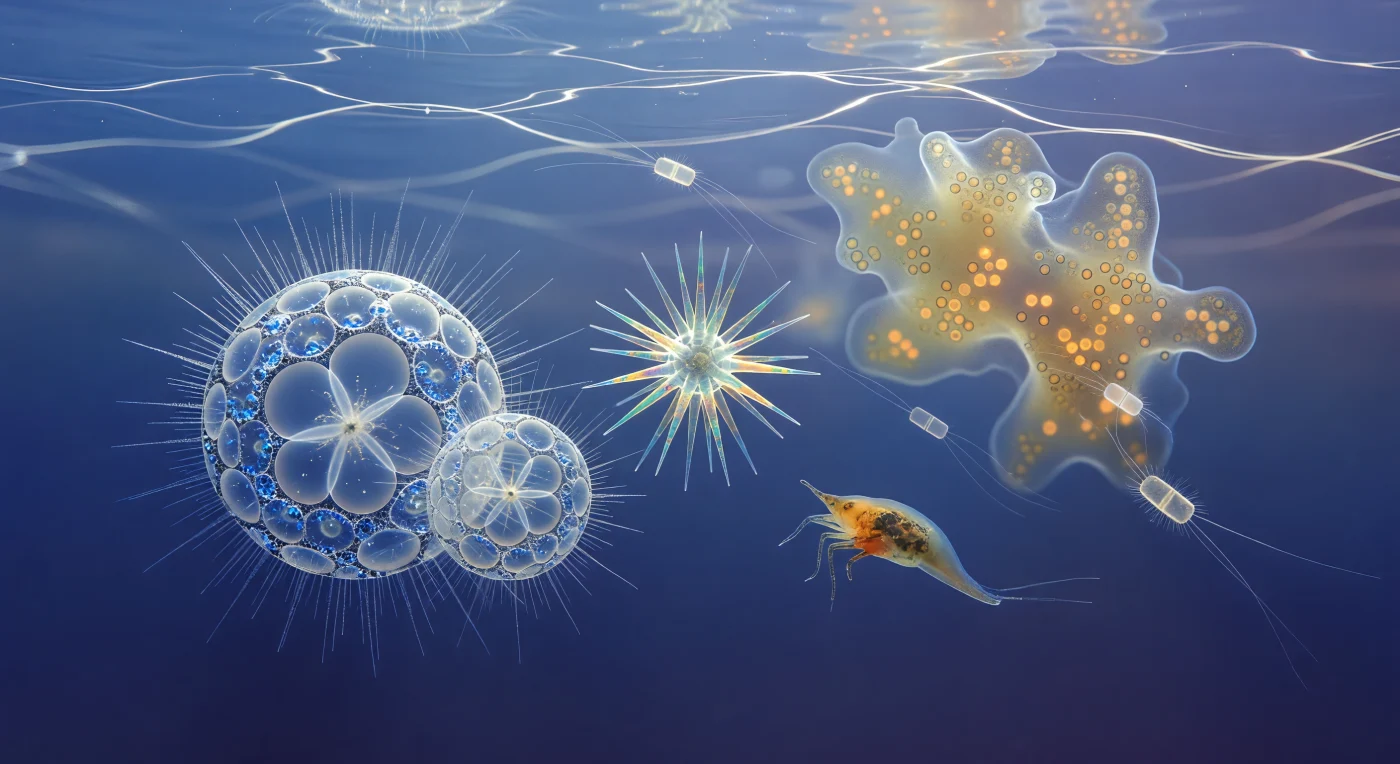
Sospesi a trenta metri di profondità nell'oceano Pacifico tropicale, ci troviamo immersi in una delle scene più straordinarie del mondo vivente: una comunità planctonica illuminata dalla luce solare che filtra dall'alto come una calligrafia luminosa in continuo movimento, tessendo reti di caustiche bianche attraverso un vuoto di indaco cristallino. Intorno a noi, le due sfere di Sphaerozoum si ergono come cattedrali gotiche di silice amorfa opale, le loro reticelle di pori esagonali — ciascuno largo pochi micrometri — disperse in prismi di luce fredda, mentre i filamenti assopodiali irradiano in ogni direzione come aloni d'argento quasi invisibili finché una caustica li sfiora e li accende in un istante. Più lontano, gli spilli di strontio solfato dell'Acantaria ruotano lentamente nella loro geometria icosaedrica, rivelando birifrangenza che scorre lungo ogni spina come un'iridescenza di acqua e fuoco — teal elettrico, ambra dorata, rosa antico — colori che esistono solo perché la struttura cristallina del minerale decompone la luce del sole in interferenza pura. La colonia di Collodaria alle nostre spalle è una nebulosa ambrata densa di migliaia di dinoflagellati simbionti, ogni cellula algale grande cinque-quindici micrometri che brucia di clorofilla come una lanterna contro il blu freddo dell'oceano aperto, mentre nauplii di copepodi — enormi a questa scala, come autobus semitrasparenti — battono i loro tre paia di arti in archi ritmici lenti attraverso un'acqua che non è mai davvero vuota, intrisa di organici disciolti e nebbiolina batterica che avvolge ogni struttura distante in un'aureola morbida e luminosa.