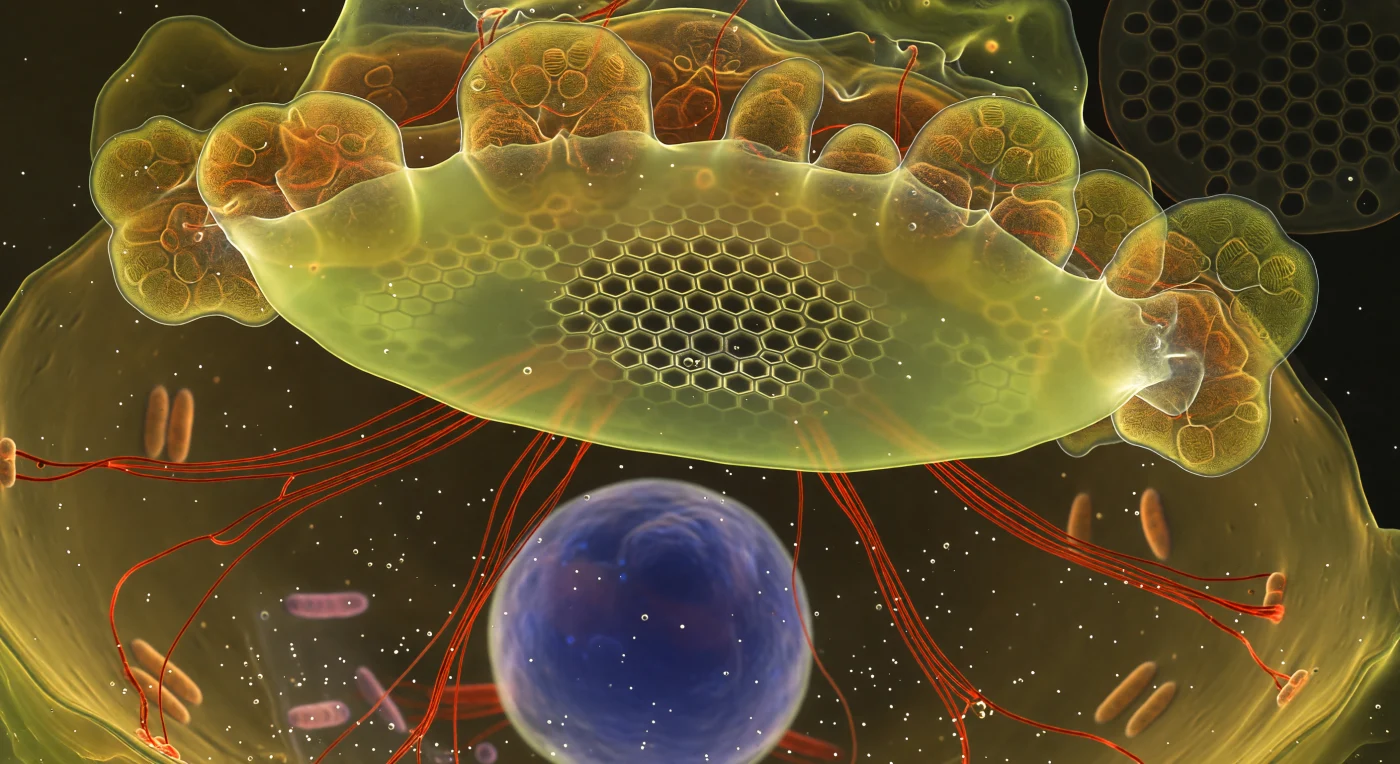
Sospeso nell'interno di una cellula di diatomea viva, l'osservatore si trova di fronte a una scena di geometria cristallina in costruzione: la Silica Deposition Vesicle si estende come un soffitto traslucido e luminoso, irradiando un verde-giallo caldo — la firma fluorescente del PDMPO incorporato nella silice appena depositata. Al centro di questa membrana, la struttura a nido d'ape è già risolta in una precisione quasi violenta, ogni areola esagonale un vuoto oscuro delimitato da pareti di silice che brillano d'oro bianco ai loro bordi; procedendo verso la periferia, la geometria si dissolve in una nebbia luminosa di materiale parzialmente mineralizzato, silice ancora indecisa sulla propria forma finale. Cavi cremisi di actina marcata con falloidina irradiano con tensione visibile dal bordo inferiore della vesicola, guidando con precisione molecolare la crescita della valva, mentre grandi lobi di cloroplasti ambrati — traslucidi come vetro di cattedrale — premono contro il soffitto di silice come formazioni geologiche animate. Più in basso, nel vuoto viscoso e caldo del citoplasma, il nucleo fluttua come una piccola luna di cobalto, e la vecchia valva parentale — un reticolo scuro e debolmente luminoso — echo in forma fantasma l'architettura che si cristallizza sopra, ricordando che questo mondo si costruisce dall'interno verso l'esterno, milioni di volte, da ere geologiche fino ad oggi.
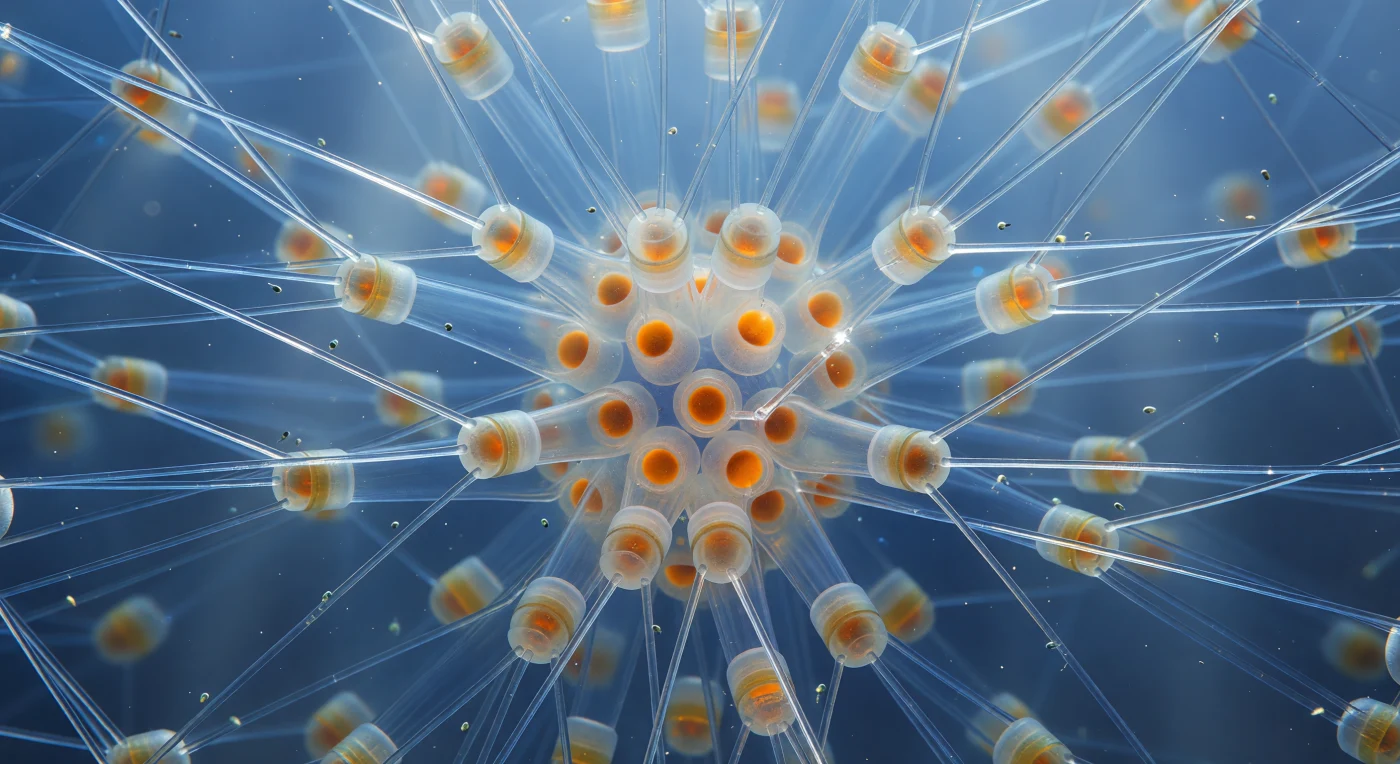
Sospesi nel cuore geometrico di una colonia di *Chaetoceros*, ci si trova immersi in una foresta di setae silicee cave che si irradiano in ogni direzione come nervature di una cattedrale di vetro: aghi quasi trasparenti, spessi quanto un filo di quarzo filato, le cui pareti sottilissime lasciano passare il blu oceanico interrotto solo da filamenti caustic bianchi e dorati là dove le superfici curve concentrano la luce in riflessi abbaglianti. Le setae si incrociano in reticoli a X tra i corpi cellulari cilindrici impilati, formando un'impalcatura tridimensionale aperta che si perde nella foschia nelle direzioni più lontane, mentre il blu sfuma dal turchese vicino alla superficie fino all'indaco freddo alla periferia della colonia. Al centro denso ardono le spore quiescenti, sfere intensamente silicizzate che diffondono la luce come braci ambrate — isole di calore organico dentro una geometria cristallina — e minuscoli flagellati grigio-verdi derivano tra le setae come pulviscolo in controluce, il loro passaggio tradito solo da guizzi infinitesimali quando ruotano nei fasci di luce filtrata dall'oceano. Ciò che all'esterno sarebbe invisibile a occhio nudo si rivela qui come un'architettura monumentale sospesa nella corrente: un lampadario vivente di silice biogenica, congelato in un istante cristallino nel freddo mare aperto.

Sospesi immobili sopra questa pianura di vetro vivente, i nostri occhi spaziano su una mandala circolare di oltre centocinquanta micrometri di diametro — un disco di silice amorfa idratata, traslucido come opale, che si estende fino a ogni orizzonte visibile come il pavimento di una cattedrale sommersa. Le areole, camere esagonali strettamente accostate in anelli concentrici, catturano la luce bianca trasmessa dal basso e la restituiscono in un calore ambrato al centro del disco, che sfuma attraverso il giallo zafferano fino a un'iridescenza violetta al margine dove le pareti di silice si assottigliano quasi nell'invisibile — un fenomeno di interferenza a film sottile, identico a quello che colora le bolle di sapone, ma fissato per sempre nella geometria molecolare di una parete biologica. I fultoportulae si ergono a intervalli regolari come ciminiere cave di vetro trasparente, strutture di supporto tubulari che in *Coscinodiscus* servono probabilmente alla secrezione di polisaccaridi verso l'esterno della cellula, e ciascuno intercetta la luce in modo diverso: alcuni ardono d'ambra pura, altri proiettano ombre ellittiche dolci sulle areole vicine, dando all'intera valva una topografia luminosa, quasi respirante. Il mezzo che ci circonda è acqua oceanica — una trasparenza cianea appena velata di materia organica disciolta — e sotto il disco non c'è fondo, solo il buio aperto del mare, mentre questa struttura di silice biologica, costruita dalla cellula in poche ore attraverso vescicole di deposizione intracitoplasmatica, galleggia in quiete sospesa come una vetrata gotica staccata dalla sua navata e calata nell'abisso azzurro.

Sospesi a cinque micrometri dalla superficie, i nostri occhi incontrano una pianura di vetro così levigata da sembrare un mare immobile, percorsa da un'unica presenza dominante: una cellula di *Navicula* lunga trenta micrometri che scivola verso destra come un battello lento su acque ferme, il suo scafo di silice biconvesso che cattura la luce obliqua in una cresta argentea lungo il dorso, mentre le strie trasversali fittissime del frustulo diffrangono la luce in sottili interferenze di blu acciaio e bronzo caldo. All'interno dello scafo traslucido, due cloroplasti color ambra dorata occupano le metà opposte della cellula come lanterne accese dietro vetro smerigliato, riversando un bagliore ocra sul substrato in due lobi elongati di luce calda. Dietro la cellula si stende un nastro di mucillagine pallido-giallino, quasi invisibile ma refrattivo, che emerge dalla fessura del raphe come una scia di seta sul pavimento di vetro; aggrappato a quella scia, un singolo bastoncello batterico — scuro, denso, due micrometri appena — rimane immobile come una virgola su pergamena chiara, trattenuto dalla stessa chimica adesiva che guida la navigazione del diatomea. Il mondo intero respira in una luminosità oceanica diffusa, fredda dall'alto, calda dall'interno, dove persino la sottilissima pellicola d'acqua marina sospesa sul substrato disperde la luce in una nebbia molecolare che ammorbidisce ogni contorno già a qualche decina di micrometri di distanza.

Sospeso a mezz'acqua di fronte a una colonna vivente di cellule di *Thalassiosira weissflogii*, il visitatore si trova occhio a occhio con una struttura verticale di dischi silicei traslucidi, ciascuno largo circa cento micron, che si susseguono verso il basso come i vetri colorati di una cattedrale in miniatura, ognuno attraversato da raggi di luce marina che accendono i lobi dei cloroplasti in un ventaglio di ocra dorata e ambra profonda. A tenerli uniti è un singolo fibrillo di chitina, così sottile da essere quasi invisibile, che emerge come un filo d'argento solo quando un fascio di luce obliqua lo sfiora, teso con una tensione architettonica sorprendente tra le strutture tubulari specializzate di cellula in cellula. La catena non cade verticalmente ma descrive una lieve spirale, ogni disco ruotato di qualche grado rispetto a quello sottostante, cosicché il filo di chitina guadagna e perde il suo lampo in un ritmo lento e ipnotico, mentre batteri scuri punteggiano le superfici di silice come minuscole sentinelle e un aggregato di neve marina deriva nel campo visivo come un fantasma grigio-biancastro, retroilluminato fino a brillare ai margini. L'architettura del frustulo — la doppia valva a Petri dish con il suo cingolo di copule intrecciate visibile come striature parallele lungo la cintura equatoriale — rivela in quel contesto oceano blu la logica evolutiva di un organismo che costruisce case di vetro biologico per catturare la luce del sole, pompare ossigeno nell'atmosfera e ancorare le catene alimentari di buona parte dell'oceano. Più in basso, l'ultimo disco della catena svanisce nella diffusione volumetrica dell'acqua, diventando un'aureola ambrata sempre più tenue finché il blu aperto lo inghiotte del tutto, ricordando che questa struttura minuziosa abita un vuoto immenso e luminoso.

Sospeso nell'oscurità assoluta, ci si trova al centro di un'architettura cosmica di silice che non ha eguali nella natura visibile: i frustuli delle diatomee, scheletri vitrei di silice amorfa idrata (SiO₂·nH₂O), fluttuano nell'etere come reliquiari di vetro colorato disposti secondo la rigorosa estetica vittoriana dei diatomeisti del XIX secolo. Davanti a voi, un *Triceratium* triangolare — lungo forse ottanta micron da un braccio all'altro — brucia di cobalto elettrico e oro fuso, ogni singola areola un foro esagonale di trecento nanometri che agisce come un reticolo di diffrazione, proiettando corone cromatiche nel nulla circostante con la precisione di un prisma levigato. Più lontano, un *Coscinodiscus* grande come una cupola irradia ambra citrino attraverso i suoi anelli concentrici di torri esagonali, mentre un *Pleurosigma* sigmoide percorre il campo visivo come una pennellata in tormalina verde e ametista, la sua griglia di areole oblique che trasforma ogni minimo spostamento di prospettiva in un'ondata di teal-violetto sulla superficie di vetro immaccolato. Questi non sono organismi morti ma scheletri mineralizzati che sopravvivono per milioni di anni nei sedimenti oceanici — architetture biologiche che la selezione naturale ha ottimizzato simultaneamente per la resistenza strutturale, lo scambio gassoso e, in modo del tutto involontario, per una bellezza ottica che nessun artigiano umano ha mai eguagliato.

Sospesi senza peso direttamente sopra il polo della cellula, lo sguardo abbraccia un continente di silice amorfa che si estende fino all'orizzonte in ogni direzione: le costae trasversali della Pinnularia nobilis avanzano attraverso l'intera larghezza del paesaggio come creste di pietra levigata, ciascuna separata da baie rettangolari dove la luce polarizzata scivola dal bianco d'osso al grigio colomba, l'intera superficie letta come elettro martellato — oro e peltro fusi insieme — che la luce radente del contrasto di interferenza differenziale trasforma in bassorilievo scultoreo. Lo sterno centrale taglia il plateau con autorità architettonica: un'autostrada luminosa e quasi bianca, delimitata da doppie ombre fredde, che conduce agli noduli polari come pilastri terminali di un viale processionale, mentre la fenditura rafeale si apre come un canyon d'ingegneria molecolare, le sue pareti tagliate con una precisione che nessuno strumento umano potrebbe eguagliare, il suo interno di oscurità assoluta suggerendo profondità ben al di sotto della superficie vitrea. Attraverso la silice traslucida — come un pavimento di cattedrale visto attraverso vetro ambrato — i due enormi lobi cloroplastici a forma di H irradiano un calore interno d'oro-arancione saturo di fucoxantina, contrapponendo la biologia organica e luminosa di sotto alla scultura minerale fredda di sopra, mentre tra i lobi il nucleo galleggia come un globo opalescente illuminato da una radiosità interna diffusa e senza sorgente. Tutto è congelato in un istante di precisione fotonica, la luce DIC che rende ogni rilievo inequivocabile come un tardo pomeriggio metallico su un pianeta deserto: il tempo qui si misura in cicli di divisione e depositi di silice, ma questa frazione di secondo di luce polarizzata ha fermato l'architettura di centinaia di milioni di anni di evoluzione in una singola e mozzafiato istantanea.

Sospesi a un centimetro sopra la distesa fangosa intertidale nel momento in cui la bassa marea la lascia nuda sotto la luce radente del mattino, si contempla non del fango, ma una città vivente di bronzo e ambra: un tappeto biofilm di diatomee pennate stipate così fittamente da formare un foglio dorato continuo che si estende fino all'orizzonte, ogni frustulo di silice che agisce come uno specchio concavo o una lamina di oro battuto, così che l'intera superficie ondeggia di una luminosità metallica calda e profonda, increspata in creste di cellule migrate in massa verso l'alto. A scala micrometrica, le cellule di *Gyrosigma* percorrono sinuose il gel di polimeri extracellulari trasparente come canoe di mogano lucidato che scivolano in un fiume di glicerina opalescente, lasciando scie di fibrille mucilaginose nel substrato; i granelli di sabbia si ergono come megaliti rivestiti di valve di *Cocconeis* aderenti in adesione permanente, i cui strie agiscono come reticoli di diffrazione e disperdono la luce solare in ventagli spettrali pallidi. Perle di acqua marina in ritirata si aggrappano alla superficie tra le creste del mat, fungendo da lenti temporanee che ingrandiscono e distorcono il tappeto ambrato sottostante prima di evaporare o scivolare via sotto il respiro del vento marino. Sullo sfondo, dove una sottile pellicola d'acqua drenante ricopre ancora il sedimento, il colore vira dall'oro brillante verso l'olivina profonda e la sienna, il biofilm sommerso e velato da una foschia lattiginosa di particelle organiche in sospensione.

Sospeso nell'epilimniom color ambra-verde di un lago d'acqua dolce, ti trovi di fronte a una delle architetture più precise che la biologia abbia mai prodotto: una colonia di *Asterionella formosa*, un asterisco piatto a otto braccia largo circa 200 µm, le cui aste di silice amorfa si irradiano da un nodo centrale di mucillagine ambrata come i contrafforti di una cattedrale in miniatura. Ogni frustula è trasparente come vetro borosilicato levigato a fuoco, e la luce diffusa proveniente dalla superficie del lago si rifrange lungo le striature delle valve in aloni di interferenza dorati e argentati, rivelando nel loro interno due cloroplasti carichi di fucoxantina che percorrono ciascun braccio come lingotti d'ambra retroilluminati. Il cuscinetto centrale di biopolimeri trattiene l'intera rosetta con legami invisibili ma tenaci, mentre la colonia ruota lentamente nella viscosità dell'acqua — un mezzo che a questa scala si percepisce come una presenza resistente e intima, capace di immobilizzare ogni micron di movimento spontaneo. Sullo sfondo, desmidi *Staurastrum* trilobe aleggiano come lanterne di giada in slow-motion, e filamenti cianobatterici si dissolvono in fili azzurro-verdi nella penombra luminosa più profonda, popolare e discreta cornice di un ecosistema che costruisce e disfa strutture di silice in ritmi di ore, ma le conserva nei sedimenti per milioni di anni.

Sospeso a dieci micrometri di distanza, il osservatore si trova davanti a una delle trasformazioni più silenziose e straordinarie del mondo acquatico: due frustule di *Fragilariopsis*, ormai svuotate della vita che le animava, derivano ai lati del campo visivo come armature abbandonate, le loro superfici di silice striata che catturano la luce diffusa in bagliori azzurro-acciaio e verde fantasma, i bordi fratturati con la geometria concoidale di un minerale antico. Al centro, l'auxospora rigonfia domina la scena con la sua membrana madreperlacea tesa come una bolla organica, già cinque volte più grande delle cellule che l'hanno generata, e sulla sua superficie si intravedono le sottili fasce arcuate del perizonium in silice — anelli depositati uno dopo l'altro lungo meridiani diversi, ciascuno un lampo fugace di oro iridescente dove la luce li attraversa obliquamente. All'interno, una massa continua di cloroplasti color ambra-miele pressa contro la membrana in espansione, punteggiata da gocciole lipidiche che ardono come piccoli soli bianchi nel tessuto dorato, mentre al centro geometrico un vacuolo trasparente e lenticolare piega la luce come una lente viva, spingendo tutto il contenuto cellulare verso l'esterno con una pressione silenziosa e inesorabile. Questo istante — la cellula che supera i propri limiti fisici, gettando via il guscio minerale per ricominciare da capo — è la risposta biologica alla legge di Müller: ogni divisione cellulare produce una figlia più piccola, e solo la sessualità e la formazione dell'auxospora permettono alla stirpe di ritrovare le sue dimensioni originali, ripristinando il ciclo in un atto che unisce la fisica delle superfici, la chimica del silicio e la logica silenziosa dell'espansione.

Ci si trova sospesi all'interno di un canale di salamoia largo appena duecento micrometri che fende il ghiaccio marino antartico, circondati da pareti di ghiaccio policristallino che emanano una luce blu-cobalto profonda, frantumata in cunei di ciano, violetto pallido e argento freddo dalle diverse angolazioni cristallografiche di ciascuna faccia, come se ci si trovasse all'interno di una cattedrale costruita con prismi sovrapposti. La salamoia ipersalina che riempie il canale ha una viscosità ottica leggermente superiore all'acqua pura, conferendo ad ogni superficie un lieve tremolio rifrangente, mentre la temperatura si mantiene a meno due gradi Celsius — condizioni estreme in cui la vita ordinaria capitolerebbe, ma in cui *Fragilariopsis cylindrus* prospera grazie a cloroplasti ricchi di fucoxantina che ardono come braci ambrate all'interno di frustuli di silice quasi invisibili, trasformando ogni cellula in una piccola lanterna autoluminosa. Intorno a ciascuna catena di cilindri da otto micrometri, un sottile involucro di gel EPS anticongelante sfuma i contorni delle cellule in un alone prismatico appena percettibile, mentre filamenti di muco aderiscono alle facce cristalline delle pareti in fili gossamer. Sul fondo del canale, uno strato denso di biomassa diatomacea accumulata colora la salamoia di un ambra brunastra ricca di cloroplasti ancora attivi, mentre in alto le grandi facce cristalline del ghiaccio agiscono da specchi divisori di luce, dirigendo obliqui fasci di luce polare diagonalmente attraverso il canale e illuminando singole catene in rilievo nitido prima che esse svaniscano nell'ombra blu-nera delle profondità più remote.

Sospesi appena al di sopra della superficie inclinata di una valva di *Coscinodiscus wailesii*, ci troviamo di fronte a una pianura di vetro amorfo color platino che si estende fino a ogni orizzonte, tessellata da migliaia di pozzi esagonali — ciascuno largo quattrocento nanometri e profondo duecento — i cui fondali recano un reticolo di velum sottile come un rosone ridotto all'invisibilità, visibile solo perché la luce radente dell'illuminazione elettronica trasforma ogni cresta in un bagliore bianco e ogni interno in un nero assoluto. Questa è architettura in silice biogenica amorfa, non cristallina: la stessa sostanza del vetro d'opale, deposta dalla cellula viva in ore precise attraverso vescicole di deposizione della silice, con una perfezione strutturale irraggiungibile da qualsiasi manifattura umana. A intervalli irregolari sulla pianura si ergono i fultoportulae — colonne cave dalle pareti sottilissime che brillano ai bordi, ciascuna affiancata alla base da tre pori satellite come archi oscuri ai piedi di un faro, condotti attraverso cui la cellula secerneva fibrille di chitina verso l'esterno. Al margine della valva, dove la superficie piana curva bruscamente nel mantello, le rimoportulae appaiono come fessure allungate tagliate nette nella parete di silice, le loro labbra esterne catturate dalla luce come sottili linee bianche su un precipizio grigio; più in basso, le bande lisce del cingulum si succedono orizzontalmente, quasi luminosamente prive di ornamenti rispetto all'intricato campo areolato sovrastante, ricordando i corsi di pietra di una muraglia che si perde nella profondità.

Sospesi appena sopra la lama di kelp, lo sguardo abbraccia un paesaggio di cellule vegetali color oliva e ambra — una distesa ondulante e traslucida che si estende come un continente sommerso, rivestita da un sottile film di mucillagine e polisaccaridi extracellulari che iridescono di oro e bronzo ogni volta che le causttiche di luce filtrata dall'acqua la percorrono in onde lente. Da questa superficie si ergono le colonie di *Licmophora*, ventagli flabellati di venti o quaranta cellule disposte in archi dorati sulle loro sottili radici mucillaginose, le frustule di silice amorfa — opale-A biogenica — traslucide e finemente striate, accendendo riflessi color ambra, testa e rame al passaggio della luce. Il biofilm è in realtà una civiltà stratificata: valve di *Cocconeis* piatte e quasi invisibili, i cui contorni ellittici tradiscono a stento la propria presenza contro il substrato, e spirali di *Rhabdonema* che si torcono tra le colonie come nastri a geometria zigzagante, i loro bordi accesi da lampi argentei. Nella scena di fondo, una cicatrice chiara attraversa la matrice di EPS — la scia lasciata da un anfipode che ha raschiato via il biofilm esponendo la superficie nuda del kelp, con i margini del muco strappato arricciati e iridescenti; la creatura stessa, grande quanto una montagna a questa scala, lavora ancora con le mandibole al bordo del varco, e le onde di pressione che genera fanno tremare in unisono ogni ventaglio dorato di *Licmophora*, come antenne sintonizzate su un'unica frequenza lenta e oceanica.

Sospesi all'interno di questa camera calcarea, le pareti ad arco si innalzano tutto intorno come la navata di una cattedrale romanica scolpita in calcare crema, la luce che filtra attraverso la calcite semitrasparente conferendo all'ambiente un chiarore ambrato e diffuso, privo di ombre nette, come se le pareti stesse fossero la sorgente luminosa. La camera appartiene al test di un foraminifero Amphistegina, un organismo unicellulare che costruisce questa architettura minerale loculo per loculo, precipitando calcite biogenica in una struttura che ospita e protegge sia il proprio citoplasma sia i simbionti fotosintetici che ne dipendono. Immersi nel citoplasma granulare color beige che riempie il pavimento della camera, i simbionti Nitzschia riposano come passeggeri in quiete: ciascuno è un frustulo di silice pennato lungo venti-quaranta micrometri, le cui valve catturano la luce indiretta restituendola come un sottile iridescente guizzo argenteo, mentre all'interno ardono i cloroplasti ricchi di fucoxantina — lobi di ambra calda e rame brunito, puntuati da goccioline lipidiche rifrangenti come perle di luce bianca sospese nell'oro. Fili di pseudopodi si snodano attraverso la massa citoplasmatica e si spingono verso le aperture nei pori della parete calcarea, tenui come filamenti di vetro stirato, unico legame tra questo mondo raccolto e filtrato e l'aperto fondale di sabbia carbonatica che scintilla appena al di là.
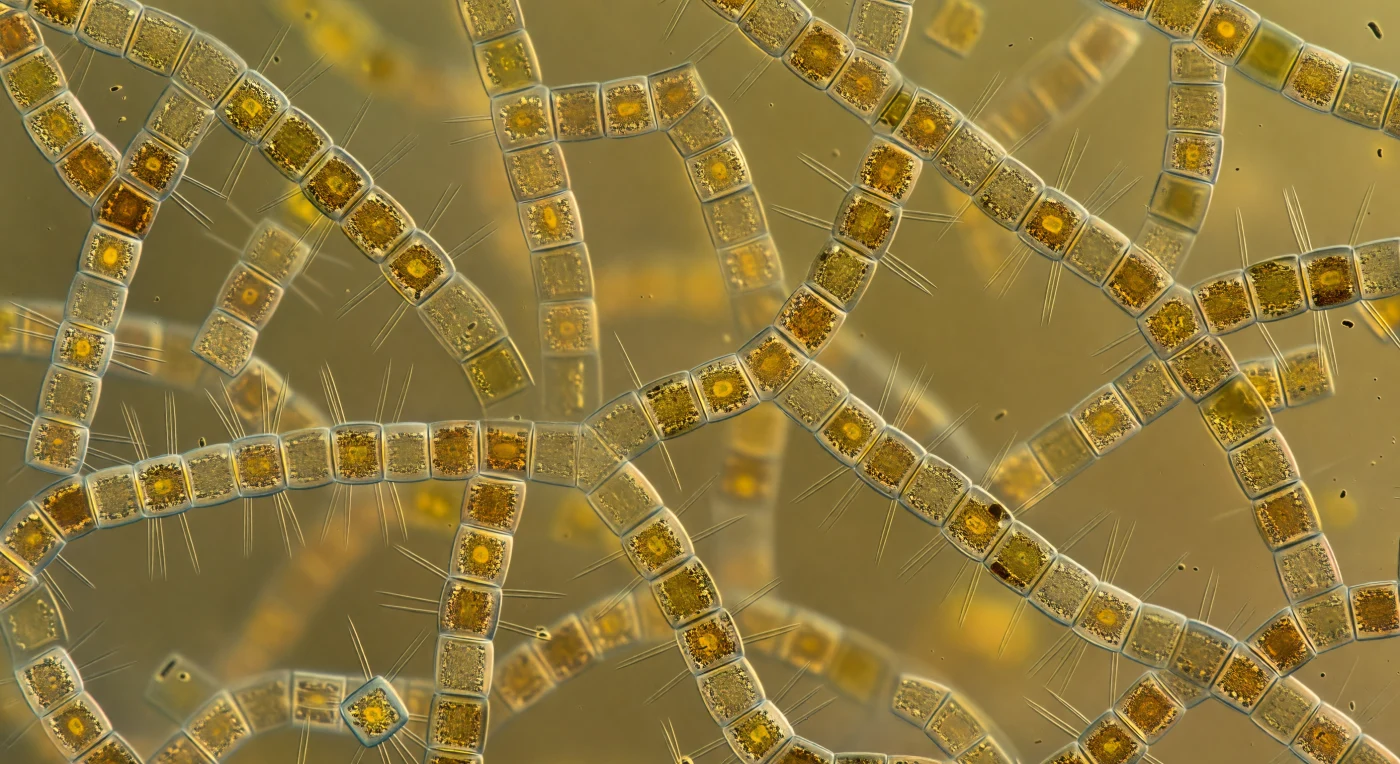
Sospeso al centro geometrico di questa cattedrale ambrata, lo sguardo incontra in ogni direzione torri cilindriche di silice — cellule di *Skeletonema costatum* impilate l'una sull'altra, larghe appena cinque-otto micrometri, collegate da raggi di fultoportule che si irradiano come costole di una lanterna esplosa, intrecciandosi con quelle della cellula adiacente fino a formare un reticolo di vetro biologico che si estende senza interruzione in ogni asse dello spazio. La luce solare che penetra dalla superficie è stata completamente metabolizzata dal bloom: ciò che scende come luce blu-bianca emerge qui come una radiosità ambrata e diffusa senza sorgente precisa, poiché i fotoni rimbalzano sulle pareti di silice, vengono parzialmente assorbiti e ri-emessi dalla fucoxantina dei cloroplasti dorati, finché l'intero mezzo liquido diventa esso stesso luminoso. Ogni cilindro è ricoperto da uno strato denso di batteri — forme a bastoncino e a virgola di appena un micrometro, così fitti da trasformare le pareti otticamente lisce della frustula in una pelliccia scura che assorbe la luce dove il vetro l'avrebbe riflessa. L'acqua tra le catene non è vuota ma una zuppa colloidale di polisaccaridi extracellulari, pigmenti disciolti e particelle virali che addensano il mezzo fino a renderlo quasi gelatinoso. Non esiste uno sfondo libero: ogni linea visiva termina su un'altra catena, un altro gruppo di cellule ambrate, un altro ventaglio di struts interlacciati, e la scena intera collassa in una nebbia claustrofobica e meravigliosa di vetro, oro e buio batterico, incandescente dall'interno con la luce metabolica di miliardi di cellule che bruciano il sole primaverile in silice e zucchero.

Ci si trova sospesi a pochi centimetri al di sopra del fondale dell'Oceano Antartico, a tremila metri di profondità, in un'oscurità quasi assoluta dove la pressione dell'acqua gelida si avverte come un peso fisico su ogni lato del corpo. Davanti si estende una pianura grigio-beige di ooze diatomacea, costruita nel corso di cento milioni di anni dall'accumulo di frustuli silicei precipitati dalle acque di superficie: dischi di *Coscinodiscus* emergono inclinati dal sedimento come finestre di vetro colorato semiinterrate nella cenere, i loro anelli concentrici di areole esagonali ancora intatti, conservati in opale biogenico con una fedeltà che sfida il tempo geologico. Tra questi, frammenti di catene di *Eucampia* e valve pennate di *Fragilariopsis* — i cui margini si sono fatti lacunosi e sfrangiati dall'azione corrosiva delle acque abissali appena al di sopra della lisoclina — si mescolano a sfere bianche di gusci di foraminiferi, alcuni ancora architettonicamente integri, altri già dissolti in una tenue macchia calcarea nel fango. Da più punti sepolti nell'ooze, brevi pulsazioni di bioluminescenza blu fredda filtrano verso l'alto attraverso la silice traslucida dei frustuli sovrastanti, accendendo per un istante le areole come array di fibre ottiche; più lontano, verso il limite dove la visibilità si perde nella foschia di particolato, un lento pennacchio pallido si alza silenzioso dove un polichete scava nel sedimento sottostante, sollevando decenni di accumulo delicato in una nube di frammenti di valve e polvere carbonatica che sale e cattura quei bagliori batterici mentre si disperde nel buio.
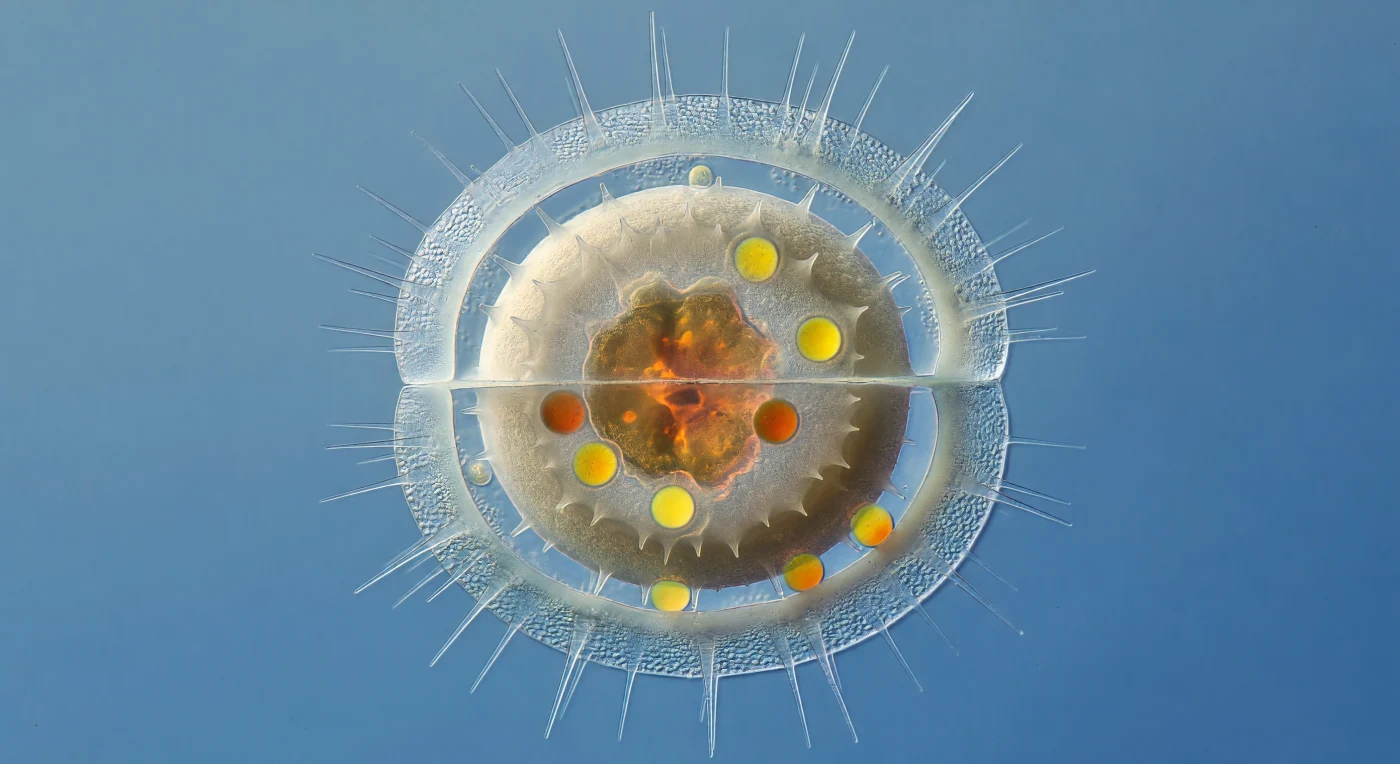
Sospesi a dieci micrometri dalla parete esterna di una frustula di *Chaetoceros* in agonia, ci troviamo immersi in una cattedrale di vetro che si dissolve e di fuoco vivo: le pareti di silice del genitore si sono assottigliate fino a diventare quasi impalpabili, fogli di opale idratato larghi appena un centinaio di nanometri, la loro iridescenza ormai spenta in un lattiginoso pallore lunare, mentre lunghi setae cavi si irradiano in ogni direzione come aghi traslucidi che catturano la luce diffusa e la disperdono in aloni spettrali. All'interno, la spora a riposo domina lo spazio con la prepotenza di un masso intrappolato in una bolla di sapone: le sue pareti spesse due interi micrometri di silice amorfa granulare — opache, avorio caldo che sfuma verso il bianco osso non smaltato — creano un contrasto tonale straniante con la fantasmatica trasparenza della cellula madre che la contiene. Questo processo di incistamento rappresenta una delle strategie di sopravvivenza più raffinate del fitoplancton: sotto stress da carenza di nutrienti, *Chaetoceros* produce una valva interna rinforzata, sigillando al proprio interno una massa cloroplastica condensata di ambra bruciata e gocce lipidiche sferiche di un giallo cadmio intenso — riserve di energia concentrate che brillano come brace sotto la luce trasmessa, visibili solo perché la frustula madre è ormai così sottile da funzionare come paralume di una lanterna che si spegne. L'intera struttura deriva nella colonna d'acqua costiera, un gioiello grezzo e pesante che cela all'esterno ogni indizio del tesoro liquefatto e incandescente che custodisce dentro di sé.

Sospeso all'interno della sezione lucida di una parete di diatomite miocenica, il visitatore è immerso in un archivio minerale bianco-avorio che si estende senza confini in ogni direzione, composto da migliaia di frustuli silicei compressi e identificabili specie per specie, congelati da dieci milioni di anni nella roccia. Le colonne di valve di *Melosira* si innalzano come torri di monete pallide, ciascuna con la sua architettura concentrica ancora leggibile nei minimi dettagli, mentre la monumentale valve di *Stephanopyxis* si apre come una rosone di cattedrale — una parete di loculi esagonali scandita da vuoti assoluti di poro che fanno risplendere per contrasto la silice circostante. I frustuli pennati di *Nitzschia* giacciono obliqui, le loro striae della rafea ancora risolte come solchi incisi nel vetro diageneticamente velato, testimoni silenziosi di un processo diagenetico che ha trasformato la silice opalina originaria senza cancellarne la geometria biologica nanometrica. In basso, un frammento di scaglia fossile di pesce curva attraverso il mosaico come una lente d'ambra traslucida, unico calore organico in un mondo altrimenti minerale e bianco, la sua superficie levigata che reindirizza la luce diffusa in un tenue bagliore ocra. Questa è la diatomite: una roccia sedimentaria composta quasi interamente da scheletri silicei di organismi unicellulari fotosintetici, depositati sul fondo di antichi bacini marini e lacustri in strati così puri da conservare la tassonomia a risoluzione di microscopio elettronico per ere geologiche.
