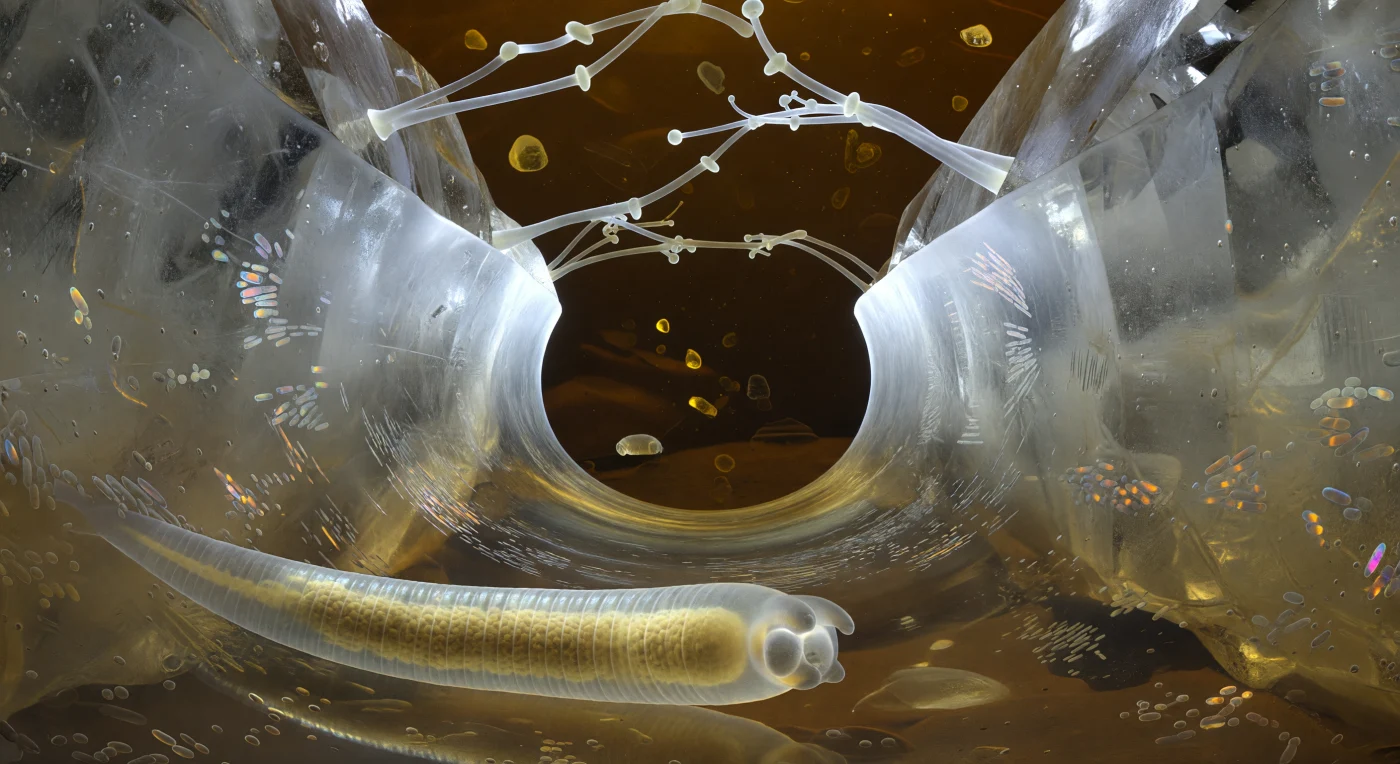
You are threading through a water-filled capillary gap between two quartz grains, their faces rising on either side like walls of pale glass — crystalline, inclusion-veined, refracting the ambient light into cold prismatic fragments wherever a surface meets water at an angle. The tunnel geometry is not carved but self-organizing: its lens-shaped cross-section is dictated entirely by surface tension, and at each end the meniscus curves outward into a blazing parabolic arch, so luminously white that it floods the corridor with a cool brightness that softens progressively into warm ochre-brown as it reaches the mid-corridor ceiling and floor, where bacterial biofilm sheathes every quartz face in a living lacquer shimmering structural iridescence — bronze, violet, aquamarine — its rod-shaped residents visible as faint parallel ridges under the meniscus-glow. Above, translucent fungal hyphae bridge quartz face to quartz face in long tensioned arcs, their glassy walls catching scattered light at each branching node like chandelier joints in an architectural rigging no engineer designed. Your own body presses gently against the water-film floor as a curved translucent cylinder, its cuticle annulations catching the lateral iridescence in short silver arcs, and within, your gut burns amber — a thick core of autofluorescent yellow-brown granules that cast a faint jaundiced glow upward through your body wall, scattering through the curved meniscus above and returning as diluted gold tinting the corridor floor, the entire scene self-illuminated by refractive geometry and biological light, the two meniscus caps blazing like white suns at each end of a world that is alive on every surface.

You are suspended at the geometric center of a crushing machine forty micrometers across, though every surface curving overhead reads to your scaled senses as the vaulted interior of a cathedral. Three wedge-shaped plates of electron-dense cuticle — laminated like polished horn, their beveled edges splitting the diffuse amber light into faint tortoiseshell interference fringes — sweep inward along radial seams, slamming together in a triradiate Y-lumen that concentrates pharyngeal fluid into violent viscous sheets, displacing shattered bacterial ghosts and spherical lipid droplets through a medium too thick and protein-laden for anything to settle cleanly. The chamber walls are living architecture: striated muscle bundles arc overhead in Romanesque ribs, each sarcomere visible as a pale-dark alternation in translucent salmon cytoplasm, with a blue-silver iridescence where actin and myosin lattices are in register and a warm rose relaxation where they are not — the whole driven by roughly 250 grind-strokes per minute, a rhythm maintained by the pharynx's own pacemaker neurons with no input from the brain. Light permeates rather than arrives, scattered inward through layers of translucent muscle and cuticle to produce a sourceless warm luminescence, while beyond those walls a faint amber pressure — the pseudocoelomic fluid pressing hydraulically inward — keeps the entire chamber sealed and taut against its own violence, every interior surface filmed with viscous fluid whose curved menisci magnify and distort the golden debris endlessly drifting past.

You are looking up from the fractured summit of a quartz grain the size of a small cliff, toward a dauer larva balanced on its tail tip like an amber minaret against a diffuse ochre glow — its sealed buccal plug the sole contact point between living tissue and ancient mineral, a posture called nictation that maximizes exposure for phoretic dispersal, the nematode broadcasting its presence to any passing arthropod large enough to carry it elsewhere. The quartz beneath fills the lower frame as a terrain of conchoidal fractures and glassy micro-plateaus, each facet deflecting multiply scattered light at a slightly different angle, the grain itself the product of millions of years of geological grinding now serving as a launching platform for an organism one millimeter tall. Through the dauer's thickened amber cuticle — its annular corrugations resolving as fine circumferential ridges under raking sidelight — large cream-white fat droplets cluster in the mid-body like pearls suspended in resin, reserves accumulated precisely because the buccal cavity is sealed and no feeding will occur until dispersal succeeds and favorable conditions trigger the dauer-exit signal. Below the grain, the world collapses into layered murk: broken soil aggregates, angular mineral shards, and looping pale fungal hyphae spanning between particle masses like frayed suspension cables in fog, the entire atmosphere thickened by bacterial exopolymers and dissolved organic matter suspended in interstitial moisture, a medium through which the nematode, were it moving, would sinuate at low Reynolds number with inertia entirely irrelevant — but here it simply stands, rigid and patient, waiting.
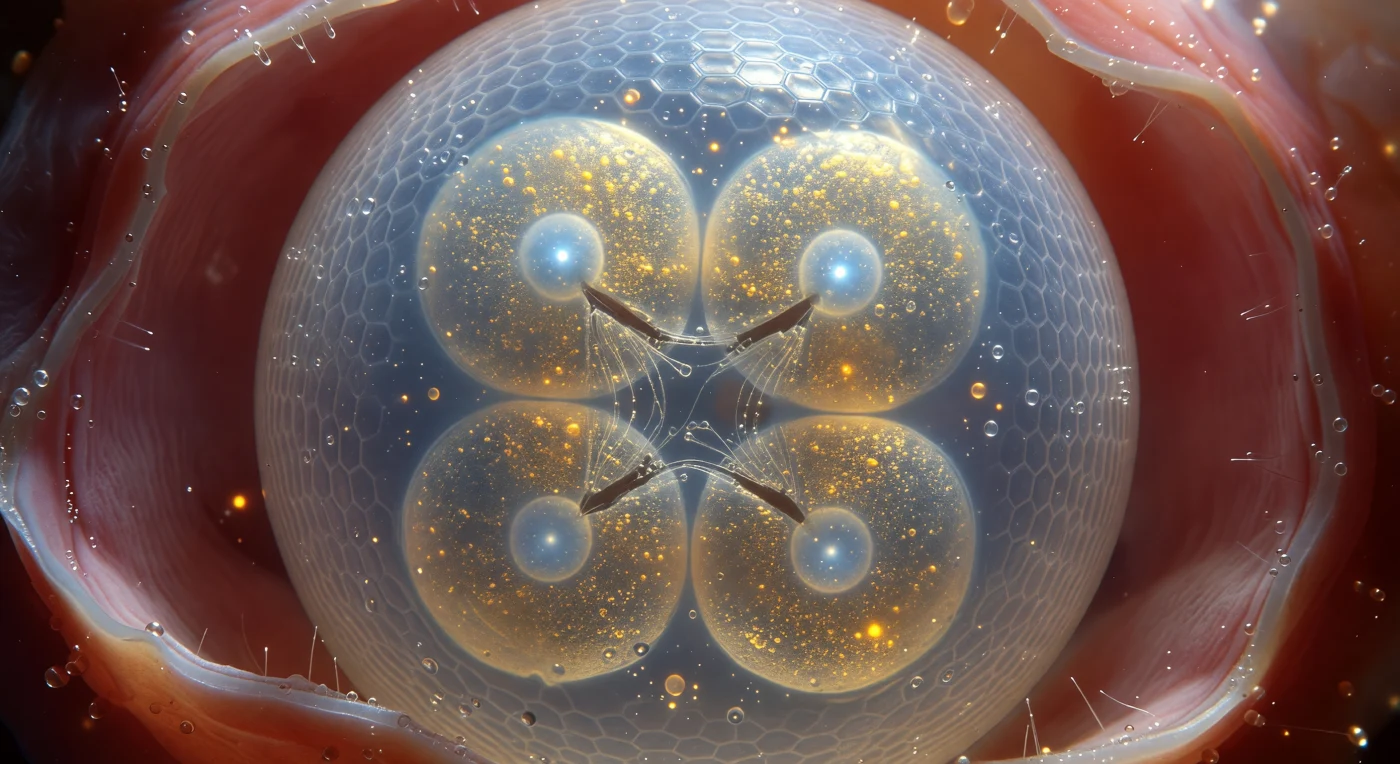
You hover so close to the eggshell that its curved face fills your entire field of view like the surface of a pale, frost-lit moon — a compressed chitinous lattice of interlocking polysaccharide fibers that scatters polarized light into shifting bands of indigo and silver, cool and luminous against the warm rose darkness of the surrounding uterine tissue. Behind this translucent barrier, four blastomeres press against one another in tight tetrahedral geometry, their flattened contact surfaces forming sharp, dark cleavage furrows while their interiors glow with dense golden yolk granules — autofluorescent masses of lipid and protein reserves laid down to fuel every division to come, from this first quartet of cells to the precisely invariant 959 somatic cells of the adult. At the center of each blastomere, a pale arctic-blue nucleus holds a single brilliant nucleolus, the ribosome-assembly engine running at maximum output for the demands of rapid embryogenesis, while between two cells the fading silver threads of a mitotic spindle — microtubules just completing chromosome segregation — catch the ambient light in brief metallic gleams before depolymerizing back into cytoplasmic tubulin pools. The perivitelline fluid sealing the space between embryo and shell is optically perfect, a pressurized aqueous medium that maintains the egg's shape while providing a chemically buffered, mechanically cushioned world in which this ancient, conserved developmental program — unchanged in its essential logic across hundreds of millions of years — executes with clockwork precision.

You stand on a surface that extends to every horizon as a vast corrugated plain of pale gold and burnished pewter, each circumferential annulation ridge rolling smoothly from crest to shadow-filled valley in relentless parallel sequence, the rounded summits catching a cold oblique light that arrives like starlight filtered through a metallic atmosphere, casting architecturally precise black shadows that reveal every micropore and surface texture with the precision of a geological survey instrument. This is the outer cuticle of a nematode — a pressurized hydrostatic exoskeleton of extraordinary biochemical complexity, built from a multilayered extracellular matrix of collagens, cuticlins, and lipid membranes secreted by the living hypodermis pressing against it from within, the whole structure held taut by internal turgor and functioning simultaneously as armor, sensory interface, and locomotor skeleton. To the left, the lateral alae rise as a true mountain range — a continuous longitudinal fin of biomechanical reinforcement running the length of the body, its summit plateau carrying finer striations that glow a fractionally warmer gold under raking illumination, casting a long hard shadow across the nearest valley and amplifying the otherwise horizontal world's only dramatic vertical relief. Near the anterior horizon, blunt dome-shaped cephalic sensilla papillae push upward from the corrugated plain like half-buried monoliths, each capped with a single specular micropore marking the external opening of an amphid or papillary neuron — sensory instruments measuring chemical gradients, osmolarity, and mechanical contact in the viscous water films where this animal spends its life. The entire landscape is absolutely still and sharp, the biological material reading as something between polished bone and oiled pewter, warm and organic at the ridge crests yet mineralic and cold in the valleys, reminding the eye that beneath this sculpted surface an entire animal breathes, feeds, and moves.

The world stretches away in every direction as an immense backlit plain, the nutrient agar terrain rolling in faint micro-ridges and glistening menisci to every horizon, its translucent amber depths set ablaze by cool transmitted light from below, and ahead of you the bacterial lawn rises as a dense, nearly impenetrable forest of dark *E. coli* silhouettes, each rod shimmering faintly with diffraction color where it stands clear at the nearest rank, the canopy behind absorbing the brilliant floor-light into deep charcoal shadow that extends without interruption to the limits of sight. Behind you, carved by your own sinusoidal passage, a cathedral corridor of cleared gel surface opens wide, its walls sheared crisp and vertical where the bacterial mat ends abruptly, blue-white edges gleaming on each exposed face, the sweeping crescent geometry of your locomotion written into the landscape in repeating arcs that curve away into middle distance where other worm trails weave their own subtly different sinusoidal signatures — older paths already hazed by bacterial recolonization creeping back from the margins. Immediately in front of you the pharynx drives its machinery in rapid rhythmic pulses, the terminal bulb compressing and releasing several times each second, each stroke drawing individual bacterial rods inward through the buccal cavity and into the amber grinder whose contents darken progressively into the warm orange-brown of a filling gut, gut granules catching the transmitted light as faint autofluorescent sparks within the intestinal cells, the entire alimentary canal visible as a brightening tube the length of a body that is itself luminous and architectural — its corrugated annulations tracing specular highlights across the cuticle wall like a rippling metallic skin stretched over a pressurized cylinder moving through a world defined entirely by surface tension, viscosity, and the slow logic of low Reynolds number existence.
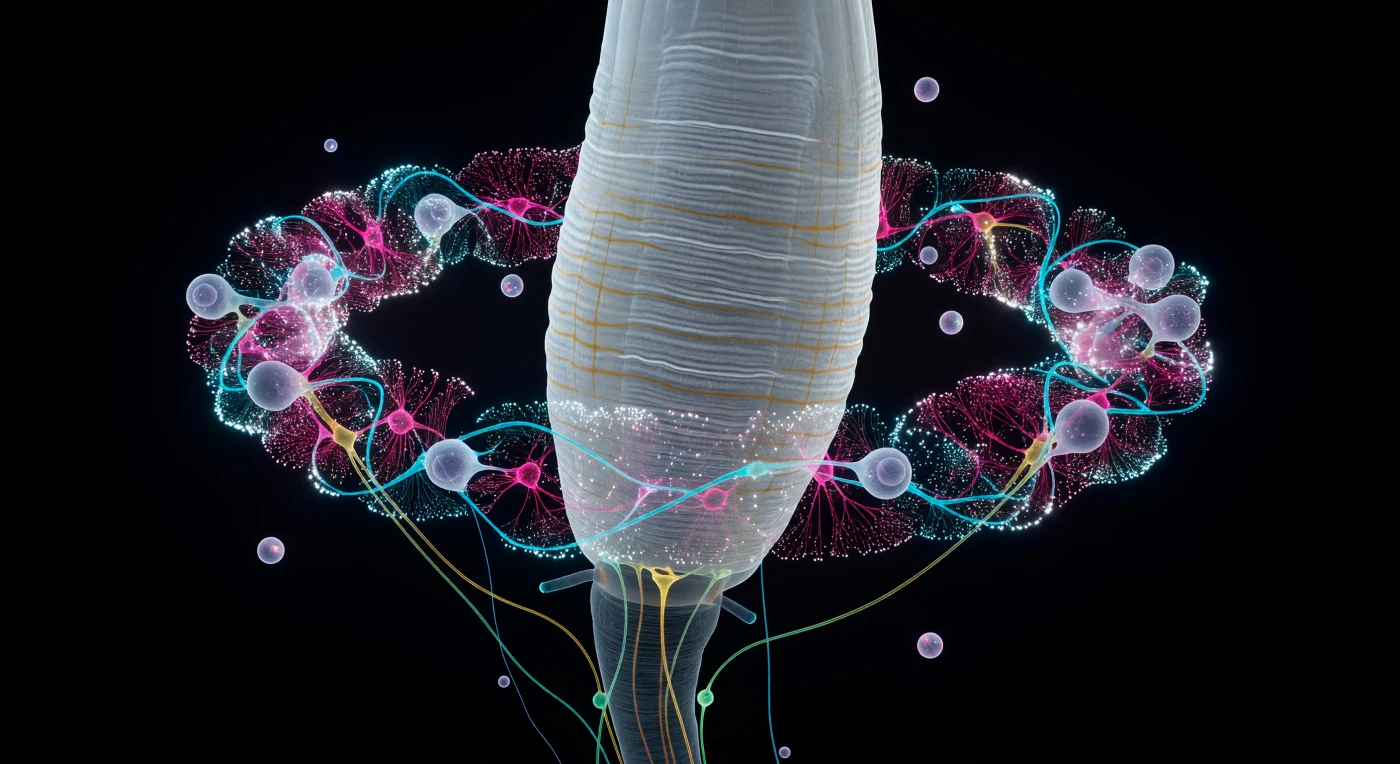
You hover less than ten micrometers from the pharyngeal wall, suspended in the optically clear pseudocoelomic void as a pale column of striated muscle tissue rises before you like the flank of a glacier — its surface carved into precise longitudinal ridges and transverse corrugations, the underlying collagen ghost-lattice just barely visible through translucent hypodermis. Around this central pillar, the circumesophageal nerve ring encircles everything: a blazing wreath of neuropil barely twenty micrometers across yet packed with a complexity that reads as continent-scale from your vantage point, each neuron class holding its own spectral signature the way minerals hold crystal color — cool cyan amphid sensory axons weaving inward from the periphery, hot magenta interneuron fibers pressing against them in fascicles separated by nanometer-scale gaps, yellow-green motor neuron commissures arcing downward toward the ventral nerve cord like suspension cables dissolving into darkness. Cell bodies of three to eight micrometers each orbit the ring like small planets, their nuclei floating inside as darker moons, while scattered through the neuropil, synaptic vesicle clusters detonate as brilliant white sparks — pinpoint supernovae of calcium-saturated membrane fusion — within a volume you could cross in a single slow drift. This living crown of electrochemical fire contains, in 302 neurons and roughly 7,000 chemical synapses, a complete and sufficient program for sensation, navigation, feeding, and reproduction, burning quietly in permanent darkness while the animal above navigates its bacterial continent one body-length per second.
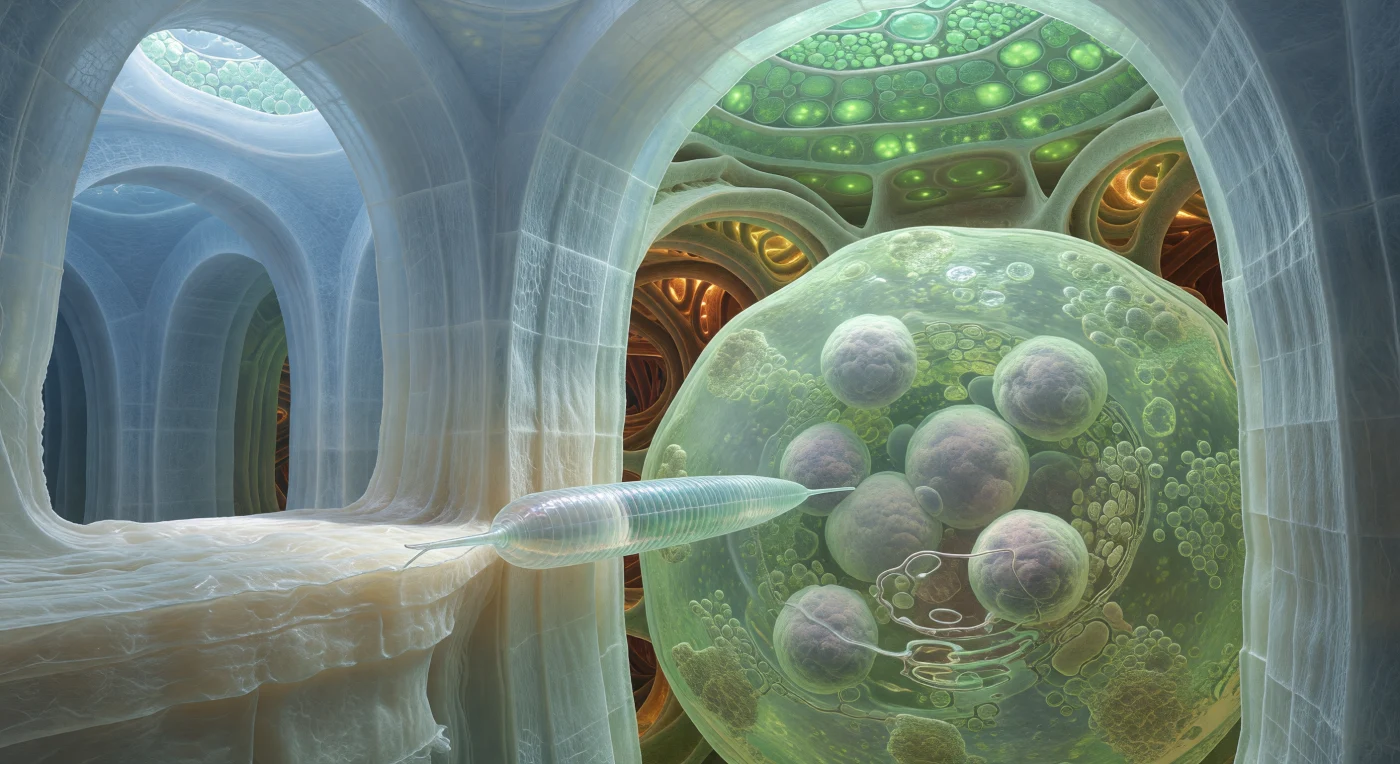
We drift suspended at the junction of two immense cellulose walls inside a hijacked root cell, pressed between vaulted panels of pale blue-white fibrous material that rise like the flooded interior of a cathedral carved from biological stone. These walls — extraordinarily thick laminated slabs of compressed cellulose microfibrils, crosshatched with a faint lattice texture — are the product of a plant cell under siege: root-knot nematodes (*Meloidogyne* spp.) inject effector proteins through their needle-like stylets that reprogram neighbouring cells to fuse, expand, and redifferentiate into giant feeding cells, metabolic factories for the parasite. The juvenile nematode pressing its stylet against the nearest wall junction is a taut pressurized tube of glassy annulated cuticle — a hydrostatic cylinder operating at negligible Reynolds number, its body wave stilled now into the focused mechanical act of puncture, every ring of cuticle catching the cool jade light filtering down through layer after layer of vacuolar water from the chloroplast-rich epidermis far above. Beyond the wall, the giant cell fills the field like a vast hall: pale lime-green cytoplasm clouded with ribosomes and metabolic machinery, multiple enormous lavender-grey nuclei floating like swollen moons, a clear central vacuole refracting the ambient light into warped caustic rings — all of it sustained by the amber-glowing xylem vessels pulsing in the deep background, their spiral-thickened lignified walls channelling the hydraulic flow that this single parasitic animal has rerouted entirely to its own nourishment.
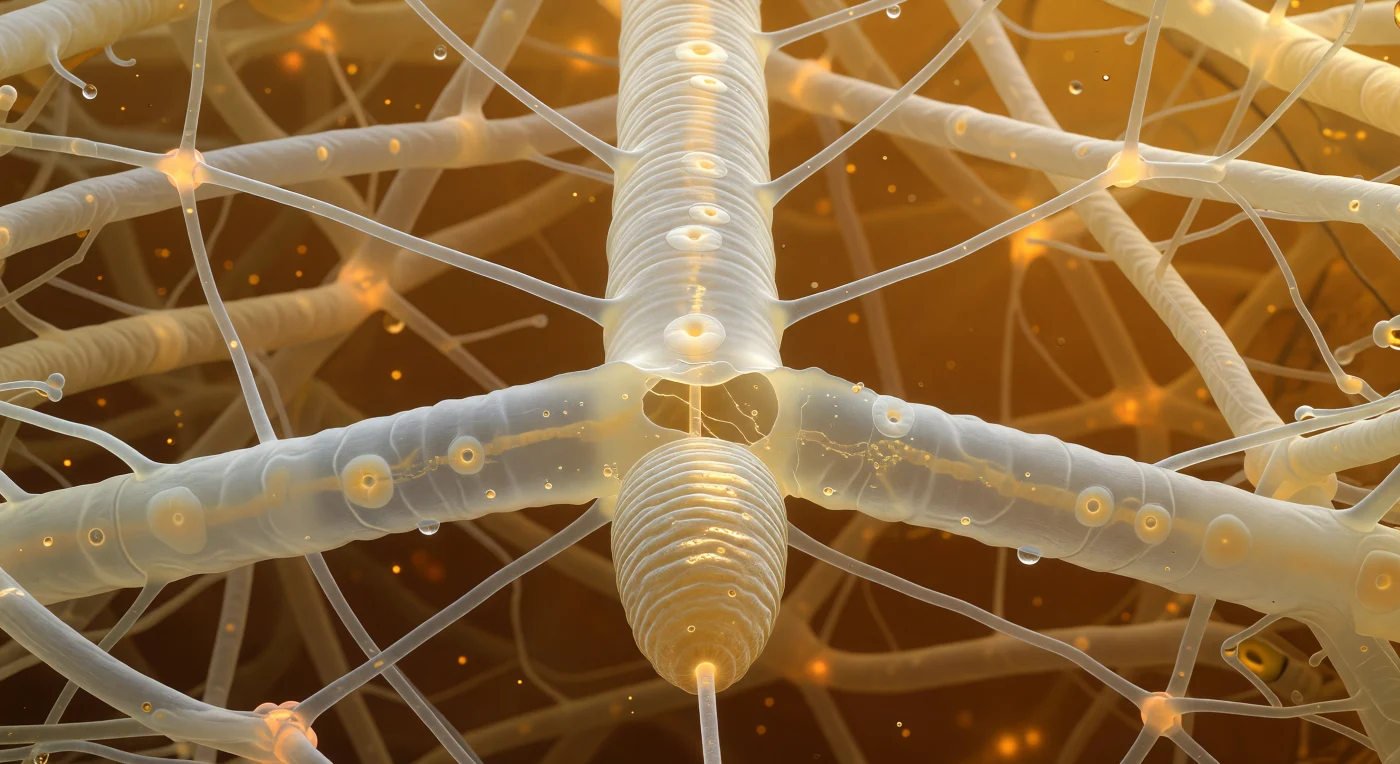
You are suspended inside a living architecture of translucent filaments, facing the broad flank of a fungal hypha that fills your entire forward view like an illuminated pressurized pipe, its wall faintly ringed with septal discs and its interior flowing with slow amber rivers of cytoplasmic granules streaming continuously through the central pore of each septum. The amber-brown light permeating this space descends through a ceiling of decomposing leaf litter overhead, the organic acids and tannins of decay filtering surface sunlight into something older and warmer, the color of whiskey held to flame, suffusing the water-saturated pore space between hyphae and cellulose fibers with an even, directionless glow. Pressed against the hypha directly before you is the anterior end of a fungivore nematode — a blunt corrugated cylinder of annulated cuticle whose ridges catch the amber light — its hollow stylet already breached through the hyphal wall, the pharyngeal pump cycling beyond in rhythmic suction that draws cytoplasmic contents inward while a pale retraction wave spreads outward from the wound site, streaming organelles deflecting around the lesion and leaving a faint clearing zone of lower optical density at the puncture margin. The background resolves into a structural lattice of swollen cellulose fibers and branching hyphae converging at anastomosis junctions that pulse fractionally warmer — almost orange — where cytoplasmic bridges between filaments exchange metabolic currency across the shared mycelial network, thin water films between every surface catching light as bright meniscal curves that define every navigable corridor at this scale.

You drift through a dim, salt-laden corridor between immense pale boulders of silica — sand grains worn smooth by oceanic tumbling, their curved flanks glowing faintly with a sulfur-bacteria biofilm that shifts between magenta and deep violet as you pass, structural iridescence rippling across grain surfaces like slow chromatic breath. Wedged into every crevice between the grains, biogenic silica diatom frustules stand as elaborate glass architecture — honeycomb discs of *Coscinodiscus* wide as plazas, broken naviculoid fragments jutting like collapsed arches — their ornate surfaces bending the sourceless blue light into soft aureoles of amber and pale gold. Through this mineral city, a marine nematode sinuates with slow muscular inevitability, its corrugated ivory body encrusted with adhered sediment particles and diatom fragments at every cuticular ring, pushing through the interstitial fluid at near-zero Reynolds number where viscosity rules and every sinuous wave of its body is a negotiation with physics rather than a conquest of it. To the right, a foraminifera shell looms in cream-white calcite like a gothic cathedral, its coiled chambers stacked into vaulted archways that recede into warm interior shadow, while from above, golden polychaete chaetae descend like structural columns from the darkness between grains — reminders that this sediment is not a quiet archive but a layered, teeming metropolis compressed into the first few centimeters beneath the seafloor.
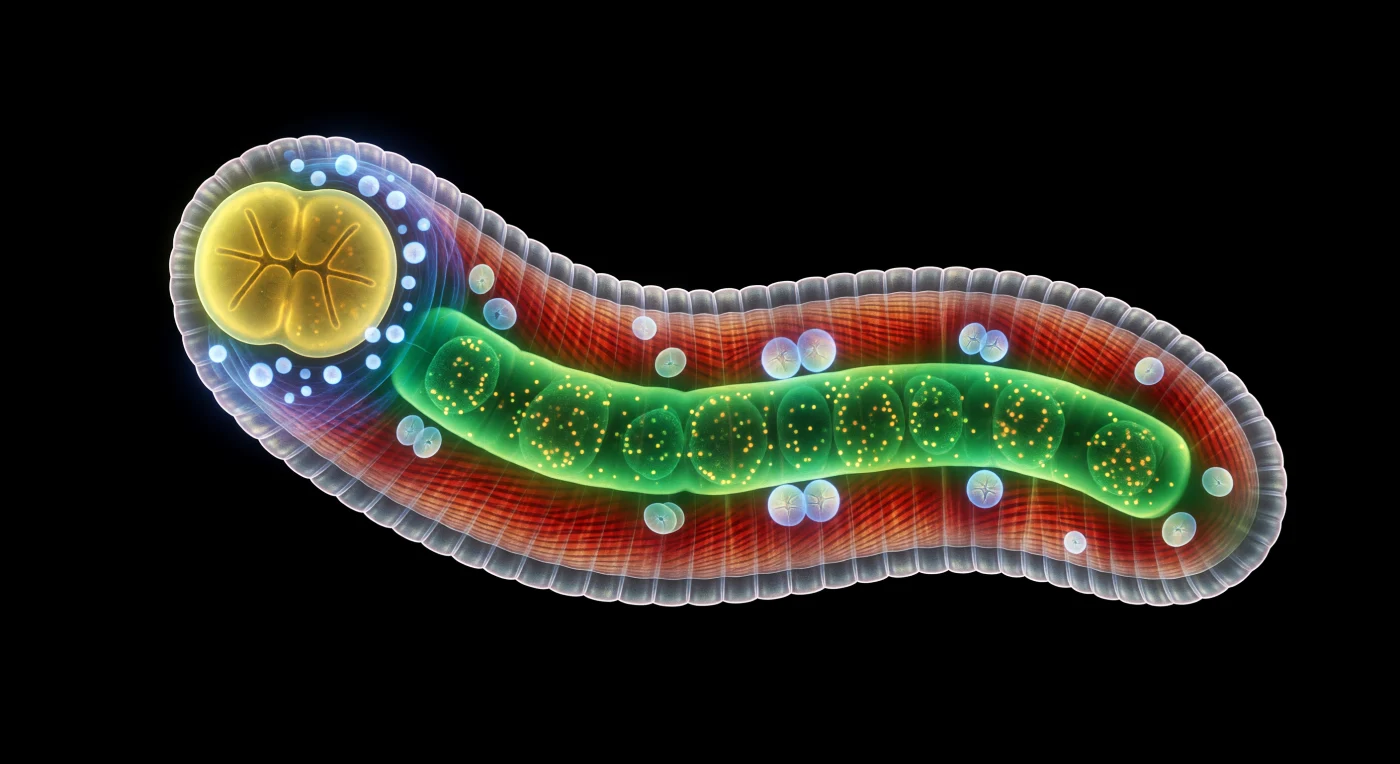
You hover at the full length of this creature, and what lies before you is not darkness but its opposite — a living architecture of self-generated light, a single adult hermaphrodite arching through absolute black in its characteristic S-curve, every photon in this void originating from within its own tissues. The body measures roughly one millimeter from its rounded anterior tower to its tapering tail, yet at this scale it stretches before you like an illuminated skyline, its translucent cuticle etched with fine annular ridges that refract escaping bioluminescence into a faint iridescent halo at the surface. Four great longitudinal bands of body-wall musculature run the full length beneath the cuticle like cathedral ribbing, each muscle cell glowing ember-red to deep crimson with diagonal sarcomere striations visible as a corrugated moiré of burning orange and mahogany — the molecular machinery of a hydrostatic pressure cylinder that oscillates at ten contractile cycles per second to drive sinusoidal locomotion through viscous films where inertia is irrelevant and surface tension governs everything. Down the central axis, the intestine blazes saturated emerald, its enormous cells packed with autofluorescent gut granules tumbling in slow cytoplasmic current, while at the anterior end the bilobed pharynx glows citrine-yellow, encircled by a crown of cool cyan neural cell bodies whose axons form the pale cerulean haze of the nerve ring — 302 neurons in total, a complete and fully mapped connectome. Cradled in the uterine space, embryos glow like opalescent pearls of blue-white, each one a miniature galaxy of precisely timed cell divisions — cleavages occurring every fifteen minutes, the entire developmental program from zygote to hatching larva encoded in a genome of just 100 million base pairs, playing out here in near-perfect darkness against a void from which no external light will ever come.

You stand at the mouth of what reads, at this scale, as a vast cathedral hewn from living amber — the buccal cavity of a *Mononchus* predatory nematode, perhaps 1.5 mm in total body length, whose gaping oral aperture at close range dwarfs your entire field of vision. The dominant structure is the dorsal tooth: a single massive arch of sclerotized cuticle, biochemically hardened into a stiff, dark reddish-brown material analogous to arthropod chitin in its mechanical role, its translucent surface refracting diffuse soil-light into caramel and burnt-ochre gradients along its razor-curved edge, while radially arranged denticles line the chamber walls in ivory-pale concentric rings, each one tapered to a piercing point, receding into the pulsing triradiate lumen of the pharynx beyond. Within this annihilating space, a smaller bacterivore nematode is already failing — its normally pressurized hydrostatic cuticle, which relies on internal pseudocoelomic fluid pressure for structural integrity, is visibly wrinkling and folding under the predator's suction force, diagonal compression creases propagating backward along its body as its hydraulic skeleton collapses, its transparent cuticle still revealing the pharynx, looping gonad strand, and gut granules glowing with a faint golden autofluorescence from within. The surrounding soil matrix — ochre-brown compressed mineral grains coated in opalescent bacterial biofilm — frames the scene like rough cliff-faces, light arriving not from any coherent source but as a warm diffuse fog, multiply scattered through a world where surface tension and viscosity govern all motion, and where this act of predation, lasting only seconds in human time, unfolds as a geological catastrophe at the scale of its participants.
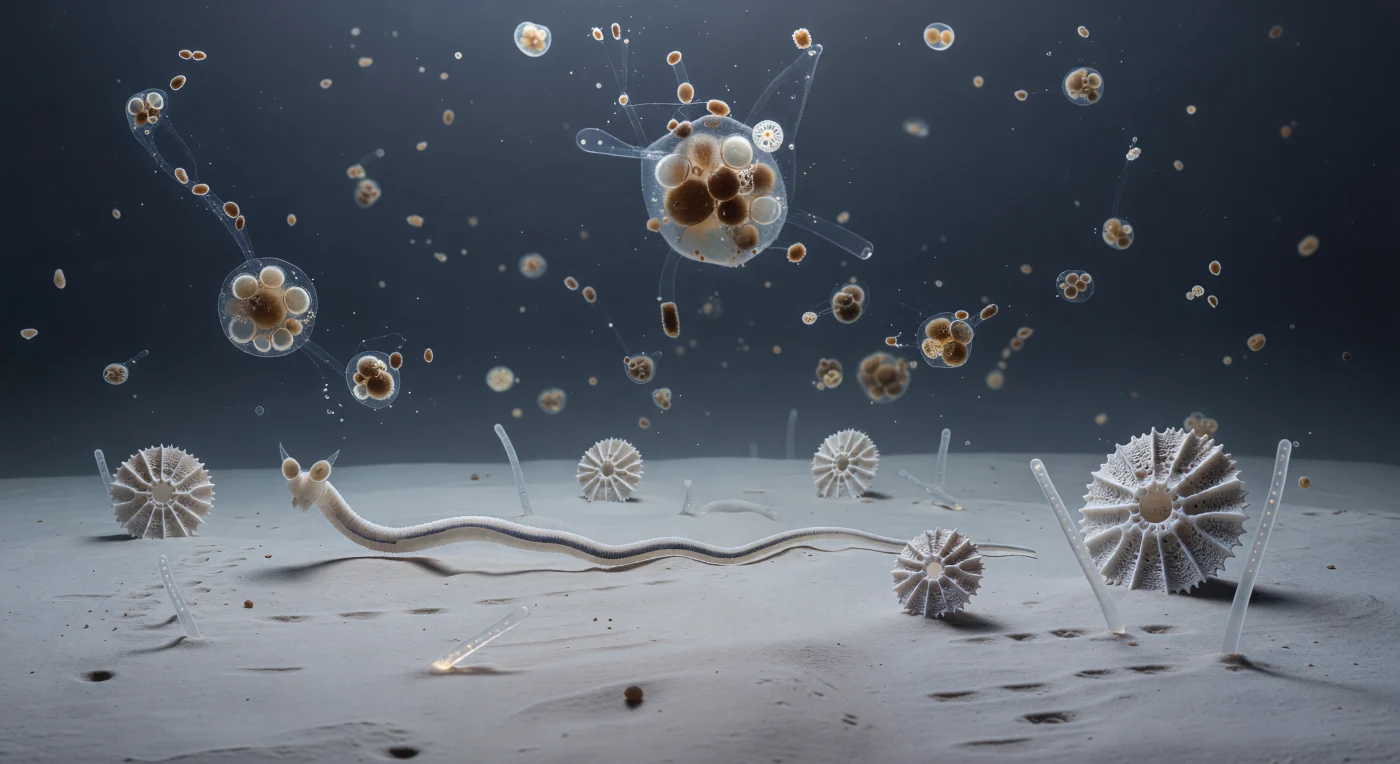
The seafloor here is not a floor in any ordinary sense — it is a nearly textureless plain of ultra-fine deep-sea clay, so cohesive and fine-grained that it behaves as a pseudoplastic surface at this scale, yielding almost imperceptibly under the slow pressure of an elongate body moving across it in long sinusoidal waves, each undulation completing so gradually that the motion registers more as geological deformation than animal locomotion. Foraminifera shells rise from the sediment like isolated cathedrals of calcite, their radial chambers and pore-studded walls the size of substantial architecture relative to the nematode body pressing through the near-bottom water alongside them, while silica sponge spicules lie scattered across the clay as enormous glassy beams, refracting the cold ambient blue into faint prismatic traces along their length. From far above, marine snow aggregates descend with the absolute patience of objects falling through a medium barely less dense than themselves — each one a loosely bound cluster of diatom frustules, compressed fecal material, mucus strands, and mineral dust, trailing transparent threads of mucopolysaccharide as they rotate in slow oscillation through the viscous bottom water, their dissolved organic compounds diffusing outward and downward into the sediment where chemosensory amphid organs, proportionally vast on the nematode's head, sample the gradient with the same unhurried precision that governs everything here. This is life at the tempo of the abyss: a world where two kilometers of ocean overhead compress into a soft brown blizzard of descending particles, where bacterial films coat every mineral surface as invisible cities, and where a nematode's 302 neurons are sufficient to navigate a cold desert of bone-pale clay and fallen silica towers in total, permanent, structural darkness.

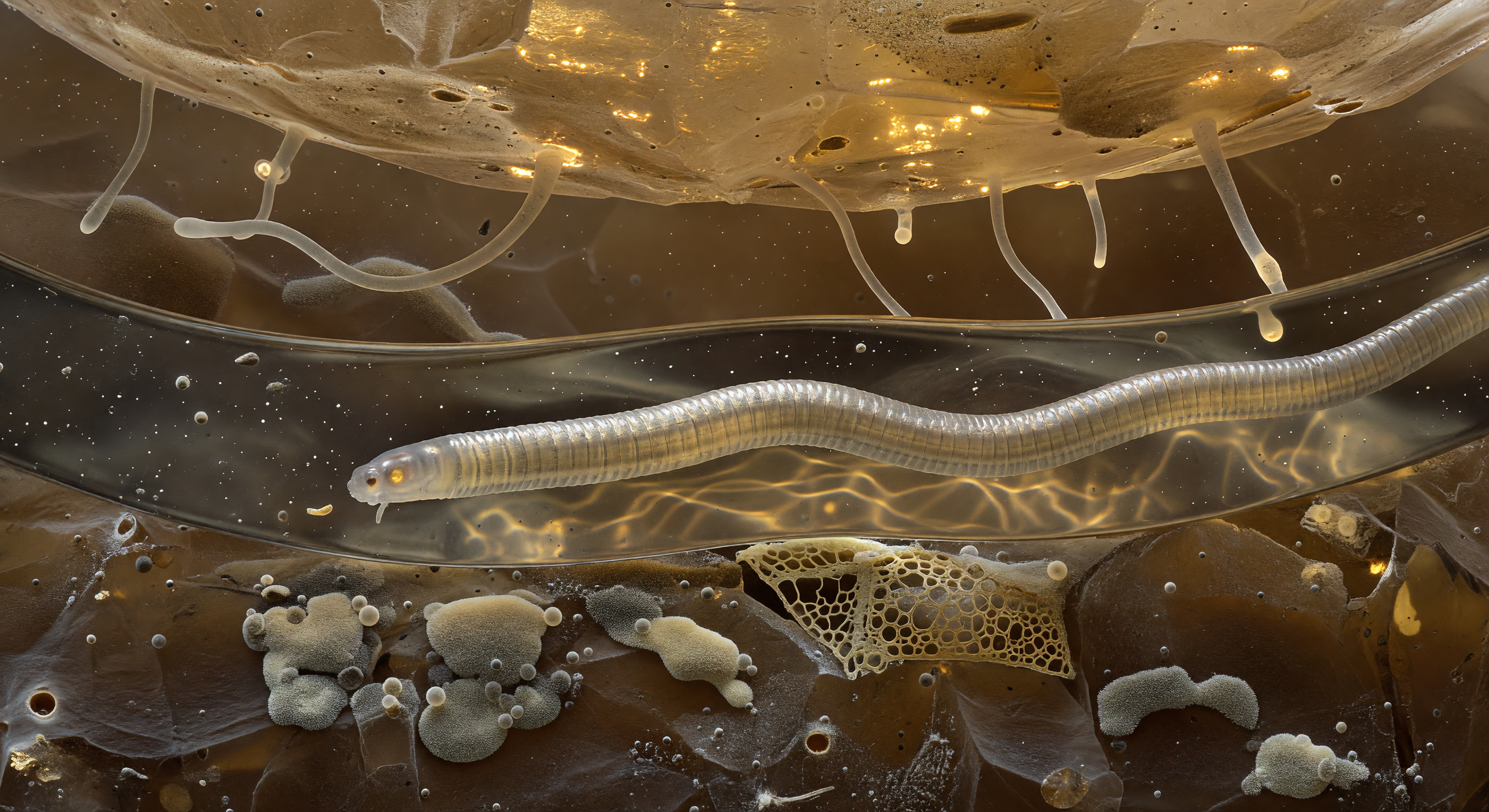
You hover suspended inside a lens of amber-tinted water no wider than a human hair, looking up through a curved meniscus dome that bows overhead like the inner surface of a soap bubble caught between two mineral boulders — a capillary bridge held in tension by the very physics of water molecules pulling against air. The meniscus acts as a convex fisheye mirror of extraordinary precision, its geometry drawn tight by surface tension, and in its curved silvered surface the entire surrounding soil landscape is radially compressed into a single shimmering panorama: frosted quartz faces, rust-stained feldspar, dark clots of decomposing organic matter, and pale corridors of gel-thick interstitial fluid, all folded together and trembling with every Brownian vibration transmitted through the medium. The water itself is not transparent but a deep luminous amber — humic acids leached from decomposing leaf litter staining the fluid warm as old tea held to light — and within it submicron platelets of kaolinite and illite drift in slow erratic arcs, each one flickering silver then rust-gold as it tumbles and scatters the multiply-diffused light arriving from all directions at once through overlying grain layers. At the edge of frame, the nematode's own cuticle intrudes as a pale corrugated wall of circumferential annuli — ridges spaced micrometers apart catching amber glints while their grooves drop into cooler shadow — and where it contacts the meniscus, the three-phase boundary where water, air, and biological surface converge burns as a continuous iridescent rim of concentrated refraction, white-gold at its crest fading to copper and rose, a luminous seam marking one of the most consequential interfaces in soil ecology: the place where life meets physics at its most unforgiving and intimate.

You hover centimeters from the anterior face of a nematode — a creature the size of a grain of sand — close enough that its three blunt, translucent lips fill your entire visual field like the mouth of a sea cave carved from iridescent alabaster, their corrugated annular ridges scattering light into pale gold and mother-of-pearl sheens across the pressurized cuticle. Dome-shaped papillae stud the lip margins like frozen water droplets, their mechanosensory endings implied by the taut stretch of cuticle over each convexity, while at the center a triradiate oral slit pulses rhythmically — the external face of a pharynx that grinds at roughly 250 strokes per minute, a muscular pump operating at a frequency closer to a heartbeat than a jaw. On the lateral flank, recessed into a shallow epidermal groove, the amphid pore opens as a crescent-shaped cleft barely a few microns wide, its glassy interior lined by sheath cells that cup a meniscus of secreted fluid like water held in polished obsidian, and within that channel twelve ciliated dendritic endings stand in a tight parallel fascicle, each filament tipped with elaborately folded receptor membranes that absorb accumulated fluorescent dye as a pale lime-white ghost-light — direct evidence of chemical signal being transduced into neural activity. The aqueous film coating the entire anterior surface is itself a map of the invisible: a saturated cyan pools near an attractant source just off-frame, bleeding slowly through teal and sage into warm amber-bronze at the periphery, a chromatic ombré of dissolved molecular gradients that, at this scale, is no longer metaphor but a physically navigable landscape of information the animal reads with its body.
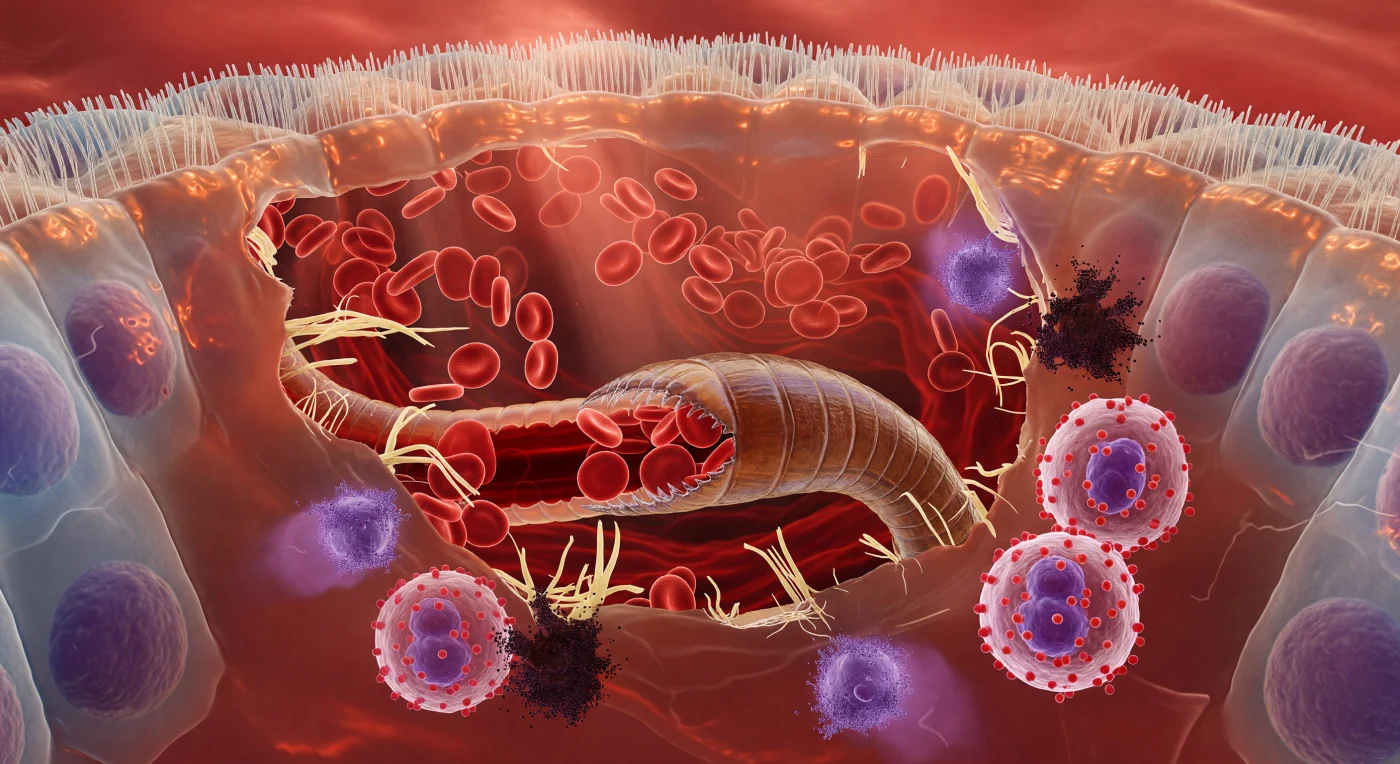
You are suspended inside a living wound — a cathedral of torn tissue suffused with warm, rust-red light filtering through the translucent walls of an intestinal villus, every surface curved, slick, and radiating biological heat. Above you, the brush border of columnar epithelial cells forms an undulating cobblestone vault: each microvillus a tightly packed translucent bristle, the cells beneath them towering cylinders of pale cytoplasm with violet nuclei hovering in their basal thirds like lanterns behind frosted glass, the whole canopy refracting the ambient glow into honeyed, wet striations. At the center of the scene, a hookworm's hardened buccal capsule — amber-brown, ridged, its cutting plates glassy-edged and pressed deep into the submucosa — has opened the tissue like a siege engine breaching soft architecture, shredded collagen fibers splaying outward in pale cream-yellow frays while biconvex red blood cells pour from ruptured capillaries in slow, laminar streams, each disc translucent and stained-glass scarlet, tumbling in currents toward the worm's open mouth and drawn inward by a steady peristaltic pulse visible through the iridescent, annulated cuticle as a dark arterial column moving through the worm's own gut. At the wound margin, the immune response erupts as a weather event: eosinophils crowd the torn edge, their salmon-red granules vibrating with contained chemistry against bilobed violet nuclei, while mast cells detonate in slow purple-black billows — granule clouds dispersing through the extracellular fluid in feathering plumes that scatter transmitted light into violet halos, the whole fluid-filled cavity pressurized and warm, a body responding in every chemical register to a parasite feeding quietly in its interior.