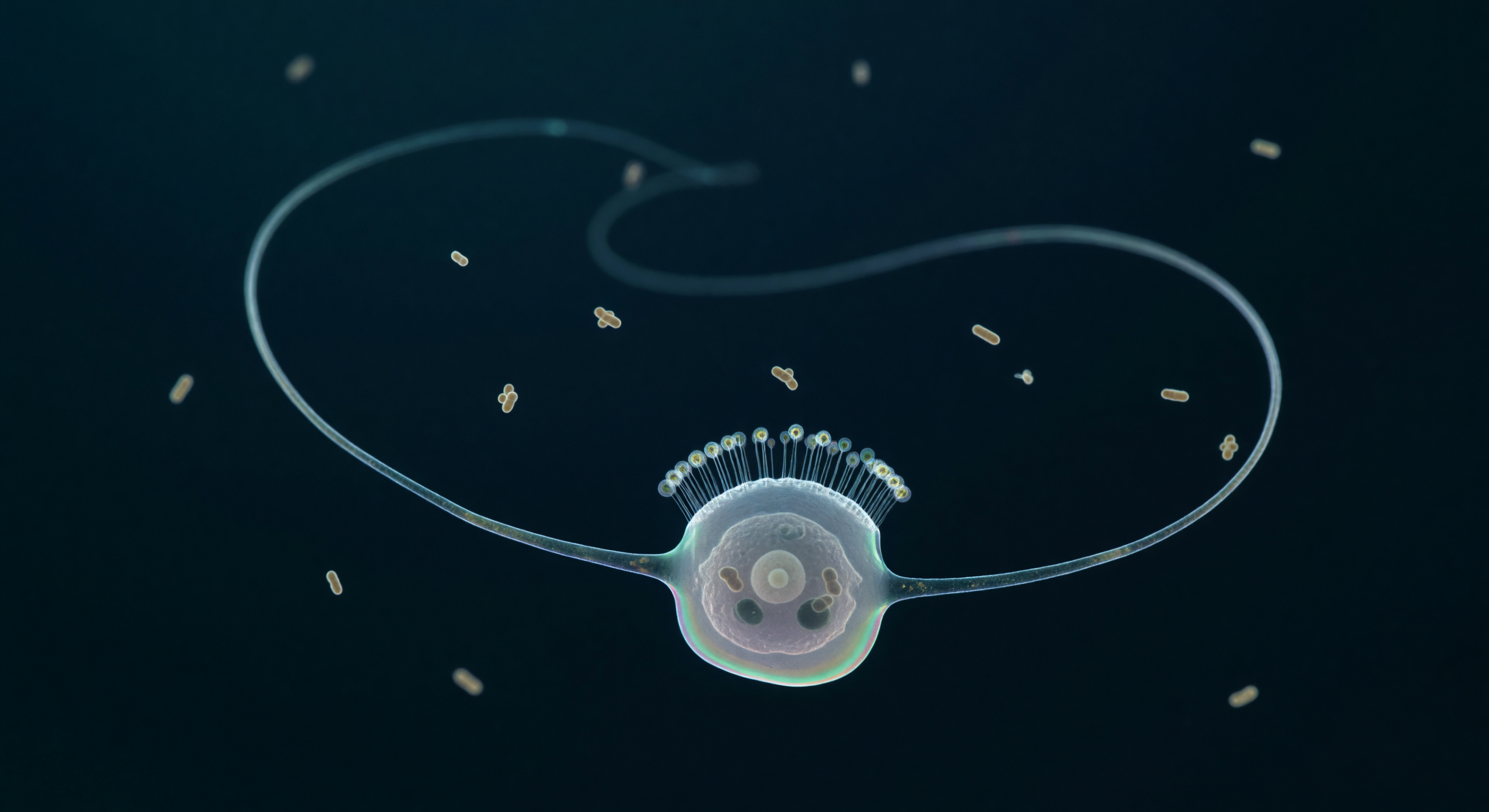
Suspendido en la oscuridad pelágica absoluta, te encuentras cara a cara con uno de los organismos más antiguos y decisivos de la historia evolutiva animal: un coanoflagelado solitario cuyo flagelo —un látigo helicoidal de veinte micrómetros— acaba de barrer el medio viscoso con la precisión de una ola sinusoidal congelada en el tiempo, dejando tras de sí una huella fantasmal de plata pálida que registra el instante exacto en que la física del mundo microscópico transformó el movimiento en alimentación. A esta escala, el agua no es líquida en ningún sentido intuitivo: es un medio dominado por la resistencia viscosa y el ruido térmico browniano, donde la gravedad carece de significado y un simple gradiente de presión generado por ese único axonema —nueve dobletes de microtúbulos moviéndose gracias a la dineína— es suficiente para arrastrar bacterias hacia el collar de treinta y cinco microvilli que rodea su base, una jaula casi invisible cuyos filamentos de actina filtran la materia orgànica disuelta como una red molecular. El cuerpo celular, una gota casi incolora de siete micrómetros atravesada por un núcleo vesicular que ocupa un tercio de su interior y por dos vacuolas alimentarias que aún conservan la silueta comprimida de bacterias en digestión, representa algo más que un organismo unicelular: es el pariente vivo más próximo a los animales, y su collar —estructuralmente idéntico a los coanocitos que tapizan las cámaras internas de las esponjas— es la prueba morfológica de que la multicelularidad animal surgió, hace más de seiscientos millones de años, de células que ya sabían atrapar, engullir y digerir. A tu alrededor, una docena de bacterias en forma de bastón tiemblan en la oscuridad teal como luciérnagas ámbar atrapadas en glicerina fría, ajenas a que la geometría luminosa que domina este mundo —un flagelo arqueado sobre el vacío como el trazo de un calígrafo— es el origen de toda la complejidad animal que alguna vez existirá.
Choanoflagellates & sponges