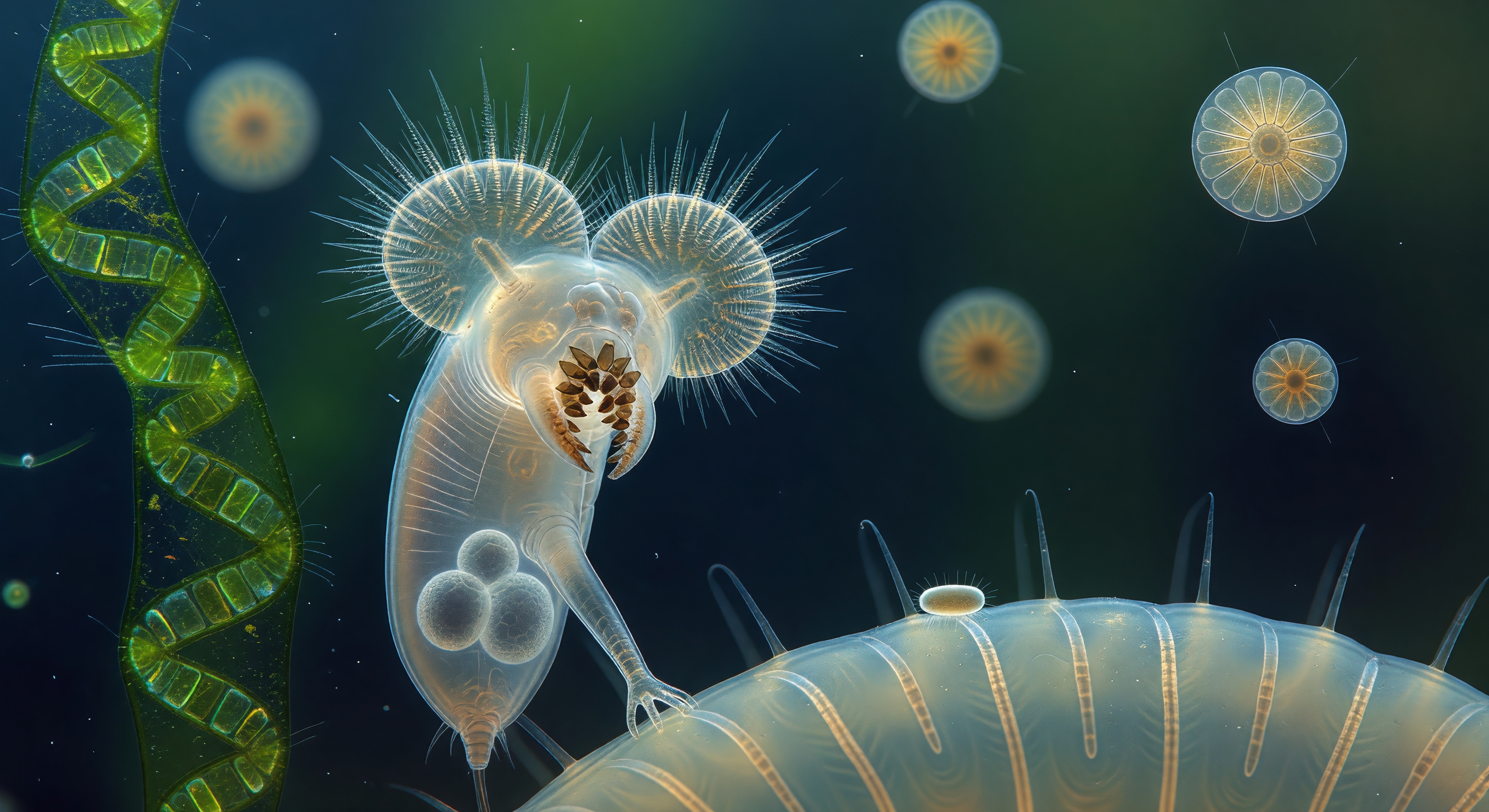
You drift at eye level with three *Keratella cochlearis*, each suspended in open water like an amber lantern no larger than a grain of fine sand, their rigid loricate shells of hexagonally faceted dorsal plates intercepting the downwelling silver light and bending it into sharp caustic stars that flicker and slide across a surrounding haze of pale turquoise. The loricas are extraordinary close up — box-like geometries of near-crystalline precision, their convex facets pooling photons at each center before surrendering them outward in soft aureoles, while six anterior spines project into the water column like drawn quartz needles and the barely visible corona shimmers in rapid metachronal pulses, each ciliary wave lasting a fraction of a millisecond as it sweeps food particles toward the hidden mastax within. Above, the water surface stretches as a ceiling of hammered mercury, its slow elastic distortions warping and magnifying the downwelling light into overlapping caustic nets written across pure liquid space, while deep in the blue-green distance a *Volvox* colony rolls with glacial momentum like a green chandelier, its daughter colonies glowing as small bright spheres within a hollow spinning sphere of emerald. Between foreground and background, the water column holds its own suspended architecture — diatom frustule fragments catching the light as slivers of opal glass, organic detritus drifting as silver threads, and thousands of *Chlorella* cells forming a living emerald smoke that gives the far water the color of shallow tropical sea glass.

You are pressed flat against a surface that fills your entire field of vision with saturated green — the outer wall of a single Sphagnum cell, a polished expanse of cellulose stretching away in long, shallow undulations toward a curved aquatic horizon, its color shifting from deep jade to cooler teal wherever the wall thins and transmitted light bleeds through from within. Your own translucent foot extends into the foreground like a glycerol-clear finger, its pedal-gland adhesive bead catching the omnidirectional light in a tiny refractive star where you anchor yourself against the surface — because at this scale, gravity barely registers, and it is chemistry, not weight, that holds you to the world. Behind your foot, amber gastric glands pulse their slow gold warmth through your glass-walled trunk, twin topaz lanterns suspended in a body so transparent that cytoplasmic striations are visible within your own tissues. Your two trochal discs blaze white-blue at your anterior, their metachronal ciliary waves merging into continuous luminous coronas as they churn the viscous film of water above the cell surface — water that, at a Reynolds number near one, resists each stroke like glycerol and snaps you to a halt the instant you stop. To the mid-right, a Pinnularia diatom frustule rises four body-lengths from the moss wall, its silica striae catching the diffuse green glow in amber and pale gold, a monument of geometric precision standing in a universe where photons scatter through your own body before they reach the substrate beneath you.

You are suspended before what appears to be an ancient underwater cliff face — the epidermal surface of a *Potamogeton* stem stretching away in all directions, its pale jade cell-wall ridges glowing from within with warm transmitted chlorophyll light, and rising from it a colonnade of gelatinous tubes studded with hand-rolled detritus pellets in amber, ochre, and rust, each tube roughly three times your height and tapering toward an open crown. These are *Floscularia ringens*, sessile rotifers that spend their adult lives anchored within self-constructed tubes, laboriously rolling individual particles of organic matter into uniform pellets and cementing them into place — an architectural behavior unique among the rotifera and one that produces, at this scale, something indistinguishable from fine cobblestone masonry set in clear mucilage. The nearest animal has extended fully into the water column, and its corona floods your entire foreground: twelve broad translucent lobes radiating outward like the tracery of a Gothic rose window, each lobe edged with a shimmering fringe of cilia beating in long metachronal traveling waves at perhaps twenty cycles per second, their collective motion threading slow spiral vortices through the particulate water and drawing flagellates and fine detritus in wide arcs toward the dark buccal field at the corona's center. Light arrives from two temperatures simultaneously — warm green-gold transmitted upward through living plant tissue, and a cooler diffuse blue-white descending from the open water column above — and in the interplay between them the tube-shadows stripe the ridged stem surface in long cool bars that recede in perspective into green-lit aqueous infinity, where the crowns of further colonies glow as small luminous discs, patient and ancient, pulsing without pause.
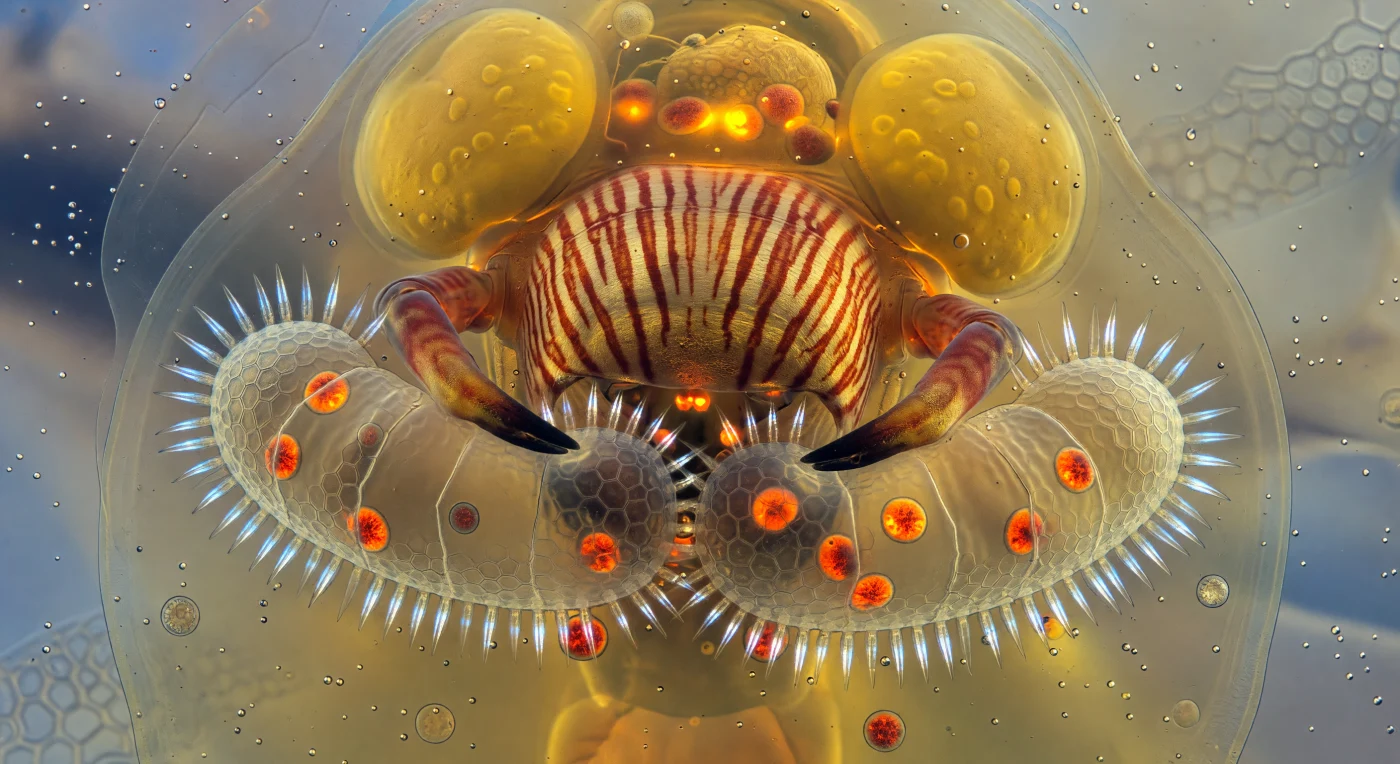
We are suspended inside the translucent body of a living predator no larger than a grain of fine sand, immersed in a viscous amber world dominated entirely by the mastax — a massive, striated muscular apparatus that fills our field of view like a cathedral organ carved from fossilized resin, its longitudinal muscle bundles shimmering in alternating bands of burnt sienna and pale gold as they contract under hydraulic pressure. Two chitinous forcipate rami arc outward at the frame's edges, their curved tips gripping the flanks of a captured *Brachionus* rotifer whose ornate lorica walls — still crisp with hexagonal surface detail in places — are visibly buckling inward under the mechanical force, while through the fracturing shell, liberated *Chlorella* cells tumble in slow viscous arcs, blazing orange-red against the surrounding amber architecture with an intensity that reads almost bioluminescent. Along the compressed anterior margin of the captive, its corona cilia continue beating in frantic metachronal sequence — each cilium catching incident light as a brief prismatic flare of iridescent white-blue, a stroboscopic halo of desperate, purposeless motion against the crushing darkness of the predator's jaws. Light here has no single source, diffusing omnidirectionally through layers of cytoplasm and lipid droplets so that every structure glows faintly from within, the entire interior of the *Asplanchna* fading from deep amber at the mastax to pale honey at the body wall to a ghostly aqueous blue-white where the outside world bleeds through as a blurred luminosity beyond.

At the edge of the retreating water film, you stand among catastrophe rendered in slow motion — the air-water interface curves overhead like a collapsing vault of flexible glass, bowing downward under its own surface tension while caustic gold lines drift across the mineral plain below where the meniscus has already pulled free and left its salt tide-marks behind. The quartz grains rise around you as rose-gray monoliths streaked with iron oxide, their surfaces pitted and warm in raking amber light that carves hard shadows into ravines between them, a terrain of pure mineral indifference. Three bdelloid rotifers occupy the middle distance in successive stages of anhydrobiotic tun formation: the first still a translucent cylinder with its germovitellarium glowing amber through the body wall, the second already folded into a matte, wrinkled opacity as its syncytium contracts and its body shortens, the third now a compact ellipsoid nested between grains — indistinguishable from mineral detritus, its metabolic machinery idled to near-zero, patient as stone. At this scale, where Reynolds numbers drop toward unity and surface tension governs everything inertia once ruled, desiccation is not death but a controlled retraction of the self into an architecture that can wait out geological time, encased in trehalose and protein glass, while the mirror above continues its slow, absolute descent.
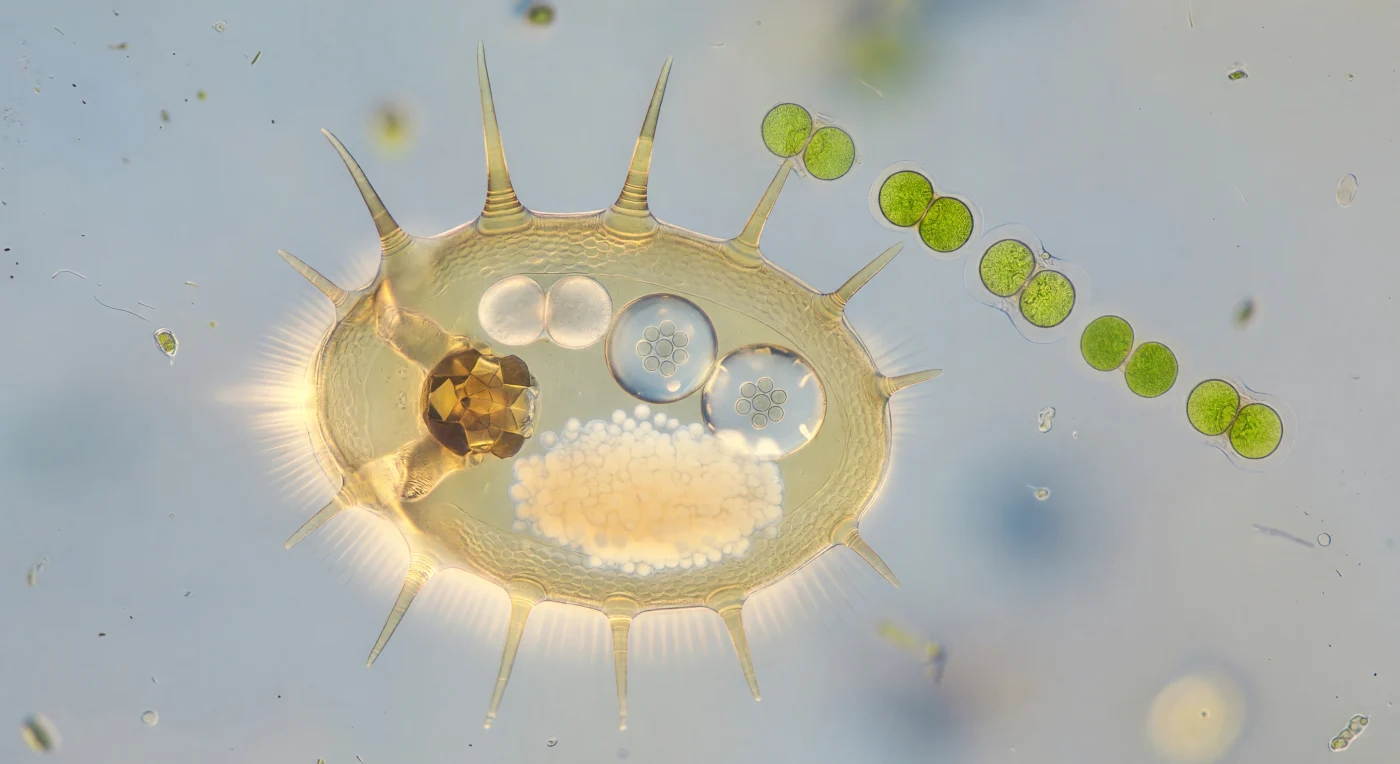
You hover motionless beside a creature nearly your own size, suspended in water so thick with viscous resistance that stillness feels earned — the female *Brachionus calyciflorus* fills your field of vision like an amber lantern, her gently ovoid lorica glowing with warm transmitted light, its surface pressed with shallow hexagonal micro-ornamentation like embossed parchment, and her six anterior spines arcing outward and upward from the lorica rim like the flying buttresses of a gothic cathedral, each one a tapered rod of translucent chitin lit from within. Through the nearly transparent lorica wall, the interior of a living organism is laid open to you as though through slightly fogged museum glass — the germovitellarium a dense cream-white cloud of compressed yolk granules glowing with milky warmth in the posterior trunk, the mastax pulsing slowly as a dark amber grinding apparatus whose rhythmic contraction reads as a slow mineral heartbeat, and at the posterior opening two eggs hang like blown-glass spheres of extraordinary clarity, each one refracting the ambient light into a soft equatorial lens-flare while inside, a tight eight-cell rosette casts pearl-grey shadows — life articulating itself in geometry before it has acquired any name. At the animal's anterior, the corona does not resolve into individual structures but blazes as a luminous atmospheric event, a wide blurred halo of white-gold incandescence where thousands of cilia beating in metachronal waves at nearly twenty cycles per second produce a continuous photonic shimmer, pushing soft pressure waves outward into fluid that registers every disturbance as a measurable displacement. Diagonally across the mid-depth of the scene, a chain of *Scenedesmus quadricauda* drifts in loose free-fall — four paired cells fused in a chartreuse-jade bracket, each one a convex lens of chloroplast-dense tissue — while the background dissolves into layered aqueous hazes and drifting particulate matter, a far diatom glinting like a distant lit window, the true optical depth of water at this scale softening even fifty micrometers of distance into a gentle atmospheric veil.

You hover just above the surface of a decomposing alder leaf, your entire field of view filled with a ruined landscape of cellulose — massive interlocking fiber bundles in deep chocolate and burnt umber, their fraying edges dissolving into translucent filaments where bacterial decomposition has softened the once-rigid wall structure, everything bathed in the warm sepia light of tannin-stained water filtering down from above like amber through stained glass. Rising before you from a cracked cellulose ridge is a single *Stephanoceros fimbriatus*, its gelatinous stalk glowing honey-amber with transmitted light, its five spirally-coiled ivory arms spanning roughly the width of a coarse sand grain — yet from within that span the cage they form reads as something architectural, a vaulted predatory cathedral whose curving members are already closing around a struggling ciliate whose surface still fires thousands of silver-bright cilia in frantic, futile motion. Inside the swollen body at the stalk's crown, the mastax — a compact geometric jaw apparatus capable of striking in milliseconds — is visibly tensed, its interlocking elements pressed together in the attitude of imminent action, while the ciliate's flexible pellicle already shows soft divots where two arms have made first contact. This is a world governed entirely by viscosity rather than inertia, where water moves like glycerol, momentum is meaningless, and every structure from the iridescent bacterial biofilms coating the fiber valleys to the amber glow of a predator's stalk exists at a scale where a human hair would loom as a massive cylindrical column overhead.

You hover in a column of jade-green water so dense with suspended life that light arrives from every direction at once, scattered through layers of algal biomass until the medium itself seems to glow — Chlorella spheres drifting as olive pearls, Scenedesmus colonies hanging like translucent staples, all of it charged with a chemical tension you cannot see but the water can read, a kairomone gradient radiating from beyond the right edge of your world, the dissolved molecular signature of a predator too large to be visible but chemically inescapable. Dominating your field of perception, a newly hatched Brachionus calyciflorus female floats in architectural improbability: her amber lorica textured with sub-micron hexagonal ornamentation, her posterior throwing two glass-needle spines outward to half her body length, their shafts tapering to points so fine they dissolve into the medium, their tips catching the scattered light as pure specular flashes, the entire structure a phenotypic argument written in chitin and transparency, assembled since hatching in direct biochemical response to that invisible chemical threat. Beside her, the earlier-generation female presents her compact, unaugmented form — blunt posterior margin, minimal spines, a sturdy geometry encoding the biological memory of a world that had not yet warned her — and the contrast between the two bodies makes visible what evolution normally hides: that morphology is not fixed but negotiated, generation by generation, with the chemistry of danger dissolved in the surrounding water. Between them, the coronal vortices of both females pull spiraling clouds of algae inward as their ciliary crowns blaze white, twin feeding halos pulsing with metachronal waves, each one a small engine of abundance operating inside a medium that is simultaneously nutrient-rich and quietly, molecularly afraid.

You float in near-total darkness at the boundary between water and packed sediment, a twilight zone measured in micrometers where two rose-gray quartz boulders — each the size of a building from where you hover — press close together, their surfaces mantled in a thick honey-amber layer of extracellular polymeric substance studded with rod-shaped bacteria visible as granular inclusions pressed against the gel from within. Through the narrow gap between them, a single Nitzschia diatom frustule glows with warm gold radiance, its silica ribs and geometric striae precise as architecture against the surrounding organic chaos. The body you inhabit — a Philodina bdelloid rotifer — is stretched nearly to transparency across that gap, its posterior foot cemented fast to trailing quartz by pedal-gland secretions, its trunk drawn thin as blown glass while the fully extended corona fans forward in a trembling iridescent aureole, each trochal disc beating its cilia in metachronal waves that refract the diatom's gold light into spectral filaments across the amber biofilm. Across the sediment floor, scattered clusters of bacterial bioluminescence pulse cold blue-green in the gel matrix, casting the undersides of neighboring boulders in cerulean wash and deepening the amber EPS to rich sienna where the two color temperatures collide — a labyrinth of grain-boulders stretching away into impenetrable darkness, lit only by one silica relic and ten thousand quiet bacterial collaborators.

Suspended in the gray-blue haze of a late-summer water column, you drift weightless alongside three bodies that together narrate the full arc of rotifer reproduction compressed into a single instant: a female *Brachionus* nearly four times larger than the dwarf male gripping her posterior lorica, and behind them both, a fertilized resting egg already descending in slow gravitational surrender. The female glows from within like illuminated amber glass, her lorica — a rigid, finely ornamented vase of hardened syncytial protein — transmitting diffuse light through her cream-colored germovitellarium and paired gastric glands, her corona pulsing in a metachronal shimmer that creates the classic optical illusion of a spinning wheel against the turbid void. Clinging to her with purposeful desperation, the dwarf male is little more than a locomotory afterthought built around an enormous milky testis — his copulatory stylet, a needle of sclerotized chitin, already engaged in the precise mechanical act of sperm transfer that represents his entire biological mandate, his vestigial corona barely disturbing the viscous medium around him. Behind both, the resting egg — a dormancy capsule of layered, opaque chitin impenetrable to light and engineered to survive months of anoxic sediment — sinks with the slow patience of a world where gravity barely registers, carrying within its sealed walls a compressed future that will not unfold until conditions permit. The extreme asymmetry of this encounter — female luminous and open, male architecturally reduced, egg closed and inert — makes visible in three floating micrometers of geometry the entire reproductive logic of a lineage that has survived half a billion years by mastering the art of becoming dormant.
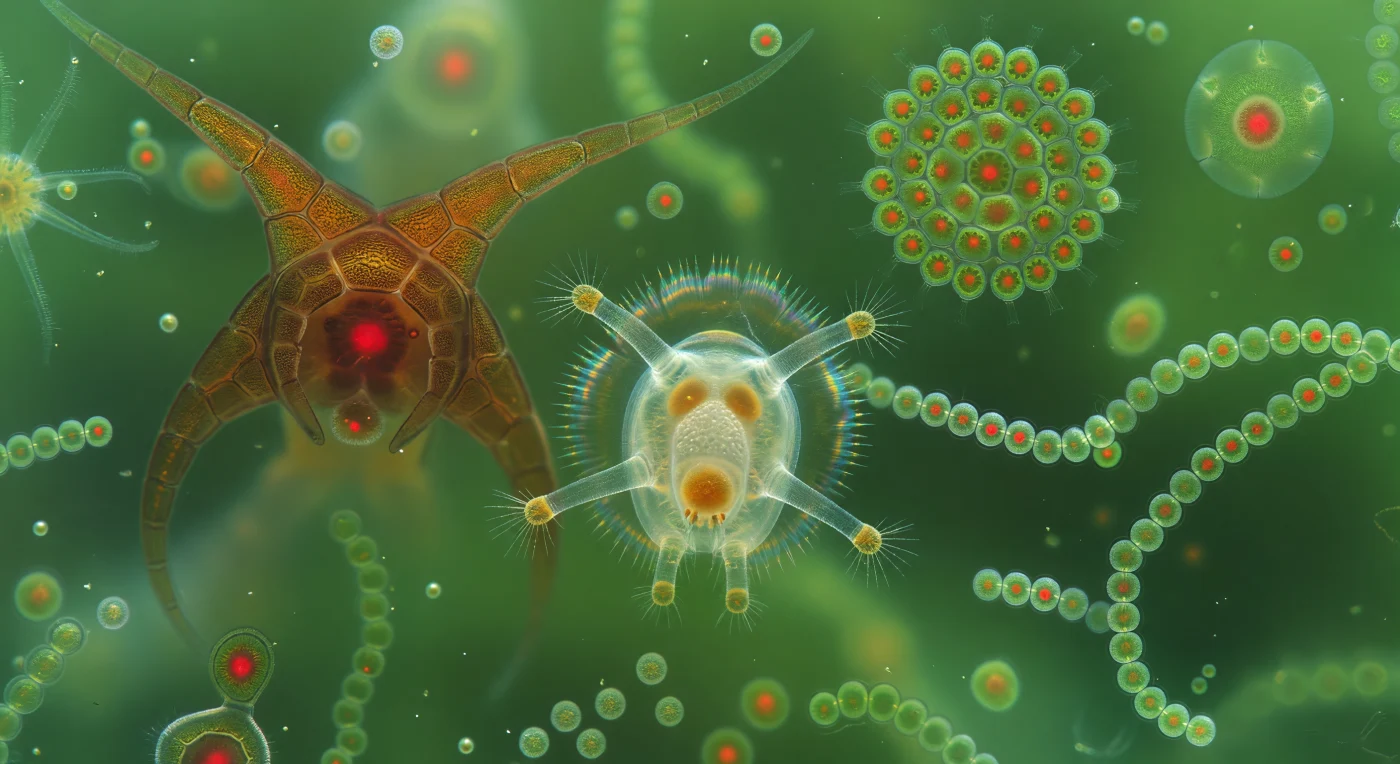
You drift forward through water that has become something other than water — a luminous suspension so saturated with photosynthetic bodies that light no longer travels through it so much as inhabits it, the jade glow arriving from everywhere at once as chlorophyll-dense cells scatter and re-emit every photon that descends from the surface twenty centimeters above. Directly ahead, a *Synchaeta* rotifer perhaps four hundred micrometers long fills the near field, its conical body a lens of living glass through which amber digestive organs and a pale germovitellarium are plainly visible, the mastax at its throat clenching rhythmically as four great auricle extensions reach forward into the medium, each tipped with trembling gold-white ciliary tufts that read the pressure field of this dense world like antennae tuned to frequencies we cannot sense. To the left, a *Ceratium* dinoflagellate presents its baroque triple-horned architecture — interlocking amber cellulose thecal plates etched with reticulate sculpture, its interior burning with a deep crimson autofluorescence where stacked chloroplast membranes conduct their photochemistry — while to the right a *Pediastrum* colony hovers like a hexagonal stained-glass window, each tessellated cell a small green lantern with its own crimson ember, the whole structure backlit by diffuse viridian light. Below it, *Anabaena* filaments drift in slow loops, each bead of jade-translucent cytoplasm carrying that same internal red glow, the chains moving through a medium so viscous at this scale that stopping is instantaneous and every stroke of every cilium must be earned against the resistance of water that feels nothing like water.

You are suspended just above the floor of an Arctic cryoconite hole, a near-perfect cylindrical chamber melted into glacier ice by the heat-absorbing dark sediment at its base, its walls rising around you in a cathedral vault of ancient compressed ice glowing cerulean-white from within — the light not reflected but diffused inward through millions of microscopic bubble inclusions sealed like silver pearls in the crystal lattice, each one scattering the column of soft polar illumination falling from the circular sky-disc far overhead. Beneath you, the cryoconite mat is a dense woven landscape of near-black cyanobacterial filaments interlocked with angular mineral grains the color of rust and iron, a compressed biogenic sediment that functions as the thermal and biological engine of this entire isolated community. Your bdelloid body moves slowly across this surface — elongated, pale, translucent — corona cilia beating in sluggish metachronal shimmer at the reduced frequency of cold-metabolic suppression, each stroke catching the omnidirectional blue-white radiance as a faint silver filament before folding back, the amber glow of your gastric glands warm against the overwhelming ice-cold palette of the chamber. Nearby, opaque cream-white tardigrades sit motionless as ceramic barrels against the sediment, and below everything the cyanobacterial mat holds the community together, fixing carbon and nitrogen in a sealed system that has been cycling through the same ancient ice-water for seasons, perhaps centuries — a time capsule of slow life enclosed in a vault of frozen light.

You hover in pale turquoise open water as a crisis unfolds directly ahead: a copepod antenna — thick as a fallen cable, dark amber-brown chitinous surface banded with transverse rings and bristling with sensory setae like iron spikes — curves across your right visual field and grips a *Brachionus* lorica suspended in the water column, while below frame its mandibles press upward into the rotifer's sealed walls. The lorica is a small amber-gold rectangular box, its ridged hexagonal surface ornament catching the diffuse blue-shifted light in faint warm highlights, but the animal inside is invisible — anterior opening sealed flush, posterior spines jutting like pale ivory thorns, all organs compressed into a dark shadow barely readable through the translucent walls — and where the mandibles press, hairline fractures radiate from the bases of anterior spines, glinting faintly like craze-cracks in stressed resin. Rotifers at roughly 100–500 µm inhabit a viscous world where water resists like glycerol, momentum is irrelevant, and stopping is instantaneous — here, the sealed lorica represents the animal's only defense against a predator ten times its mass, withdrawing into its own box and closing every aperture while the chitinous walls absorb the crushing load. To the far left, a second *Brachionus* feeds in total indifference, its corona blazing into brilliant white luminescence, metachronal ciliary waves pulsing at 15–25 Hz in a shimmering living wheel, mastax grinding warmly visible through its translucent body — and the contrast between that radiant, open, pulsing animal and the sealed amber box cracking in the foreground is immediate and visceral, one organism in full biological expression while the other waits compressed inside its own fracturing walls, the copepod massive and implacable as a fallen tree.

You hover suspended in a tannin-amber water column enclosed by bromeliad leaf walls that curve away on all sides like the interior of a vast waxed cathedral, the cuticle surface so close on your left that its mosaic of convex epidermal cells — each one a low dome of jade and crimson, separated by micro-valleys furred with bacterial biofilm — fills your entire visual field like an ancient tiled architecture. The water is not transparent but a warm luminous broth, every photon stained weak-tea gold by dissolved humic acids and tannins leaching from the decomposing organic matter of this sealed phytotelm ecosystem, so that all shadows are amber and all distances dissolve into caramel fog. Against the leaf wall, a Lecane presses its glassy oval lorica flush to the cuticle, its corona tufts sweeping biofilm in urgent strokes while its mastax pulses amber flashes through the translucent dorsal shield — a creature for whom viscosity is architecture and stillness is grip; nearby in open water, a Philodina bdelloid extends both trochal discs into shimmering halos of metachronal cilia, their offset beat frequencies creating the optical illusion of counter-spinning wheels of pale light against the amber, while through its transparent body the germovitellarium glows like a dense pearl and flame cells flicker cold white along the lateral walls. To the right, a Cephalodella hangs motionless at its predatory angle, forcipate trophi half-extended and catching the amber light as geometric hooks, while above everything a fragment of decomposing leaf descends through the background with the gravitational presence of a collapsing building and a single mosquito larva seta enters the frame from the upper edge as a vast barbeled architectural cable — a suspension element for the entire amber world.
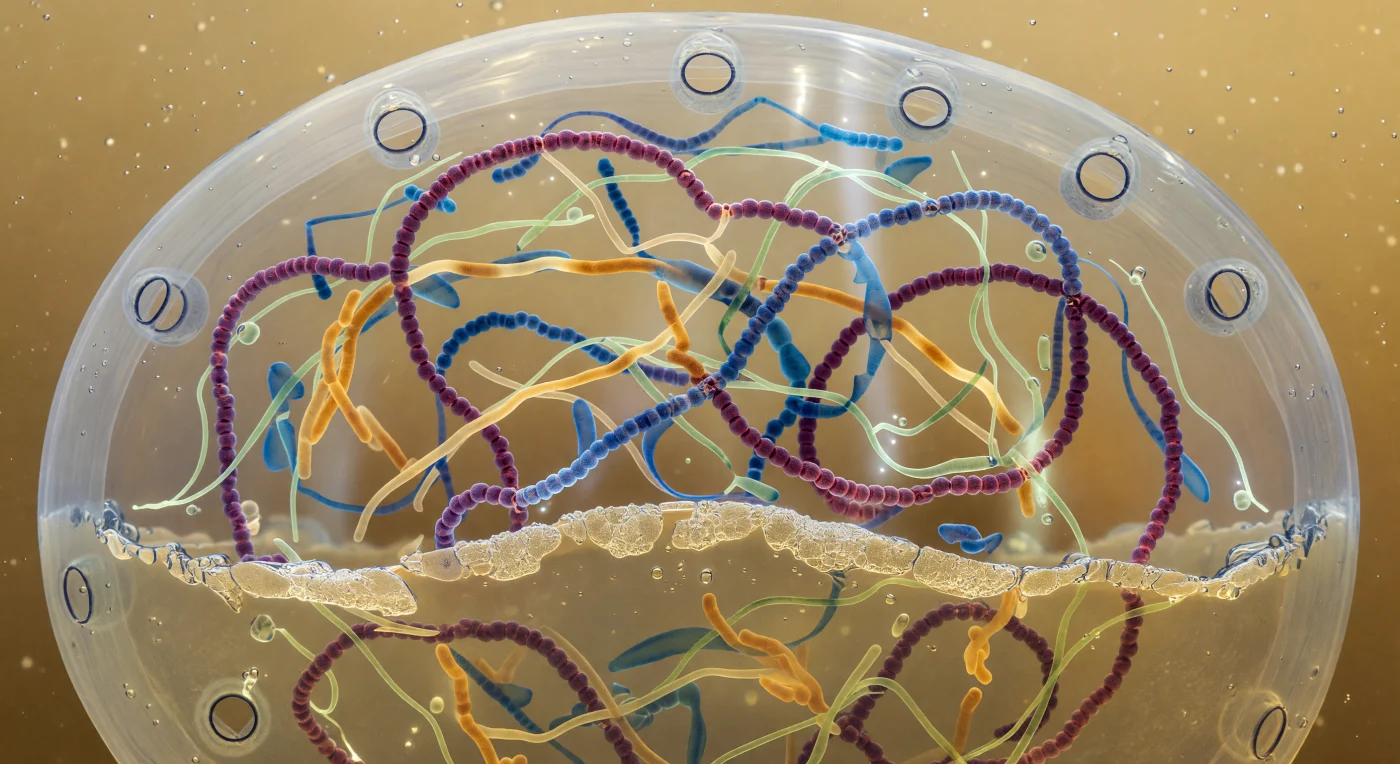
You are suspended inside a living nucleus at the moment it remembers how to be alive. Everywhere around you, the double-layered nuclear envelope curves away like the inner surface of an immense translucent dome, its waxy pearl membranes punctuated by barrel-shaped pore complexes whose protein-rimmed channels gape open at their maximum dilation, admitting the first incoming molecules as the cell emerges from cryptobiotic arrest — a state in which bdelloid rotifers can survive complete desiccation for years, their cellular architecture preserved in an amorphous glassy matrix until water returns. Rising from below is that return: a slowly climbing frontier of hydrogen-bonded order, not quite liquid and not quite solid, a geometric transparency that bends the warm amber light of the surrounding cytoplasm into prismatic planes wherever it advances, converting desiccated rigidity back into viscous, functional space nucleation point by nucleation point. Suspended within this awakening interior, four distinct chromatin lineages interweave in a canopy that is itself a record of extraordinary biological history — the thick violet-red coils of the rotifer's own chromosomes entwined with thinner bacterial, fungal, and algal DNA sequences incorporated through horizontal gene transfer, a mechanism so pervasive in bdelloids that roughly one in ten of their functional genes carries a non-metazoan origin, a genetic cosmopolitanism invisible at any larger scale but written in molecular detail across every strand visible here. As the water-lattice rises and osmotic pressure is restored, these chromatin threads begin their slow unfurling, hydrogen bonds reforming along the double helix, the nucleus rebuilding its architecture from the bottom up in a resurrection that has been rehearsed, in this lineage, for perhaps eighty million years.

You are suspended within the sediment column itself, pressed between clay platelets the size of flagstones and dark fibrous ropes of decomposed organic matter, while the sediment-water interface blazes above you as a single cold-white luminous seam — the entire sky of this world compressed into one diffuse horizon. Below that threshold, the matrix grades through straw-gold and pale ochre into deep tobacco brown and then into zones of near-absolute darkness where photons arrive only as grey scatter, carrying within it a stratigraphic record of rotifer reproductive history: dormant resting eggs distributed throughout the column, each a sphere wrapped in hexagonally interlocking sculptural relief that catches the distant ceiling light in brief flares of gold. These are diapause eggs — ephippia and subitaneous resting stages produced when populations faced seasonal stress, drought, or predation pressure, their outer walls hardened into structures capable of surviving decades or centuries of anoxic burial, each one housing an arrested embryo in metabolic suspension, visible through a faint equatorial translucency as a pale gold ghost of deferred life. The uppermost eggs, closest to the luminous interface, remain warmest amber and most translucent; deeper eggs have oxidized to mahogany and near-opaque umber, their geometry architecturally intact but their gold relief softened to tarnished bronze; and among them cluster the hollow split shells of hatched eggs — collapsed amber lanterns, their hexagonal tessellation preserved but their interiors empty, ghost architectures of animals that woke from diapause and swam upward into open water seasons or centuries ago, leaving behind a sedimentary archive of every population collapse and resurrection this lake has ever witnessed.