Confiança científica: Alto

O observador flutua ao nível da superfície de um virião de influenza, rodeado por uma floresta densa de trímeros de hemaglutinina que se erguem como colunas de marfim com cerca de 13 nanómetros de altura, as suas coroas levemente alargadas cintilando com uma luz difusa e fria que parece emanar de todas as direções ao mesmo tempo. Dispersos entre esses pilares, os tetrâmeros de neuraminidase destacam-se como cogumelos de teal escuro e verdigris, mais baixos e compactos, quebrando a monotonia das colunas pálidas com a sua geometria e cor contrastantes. O meio envolvente não é espaço vazio, mas sim uma névoa densa e luminosa de macromoléculas em suspensão — glóbulos de albumina sérica derivam como lanternas âmbar translúcidas, enquanto longas cadeias de glicoproteínas e filamentos de mucina ondulam lentamente como algas em corrente submarina, os seus grupos laterais de açúcar refratando reflexos lilás e azul-gelo. A visibilidade extingue-se a poucos pilares de distância, onde a floresta molecular se dissolve numa neblina opalescente de branco e âmbar, e o plano da bicamada lipídica por baixo treme em ondulação térmica contínua, vivo com o agitar constante e inexorável do movimento browniano.

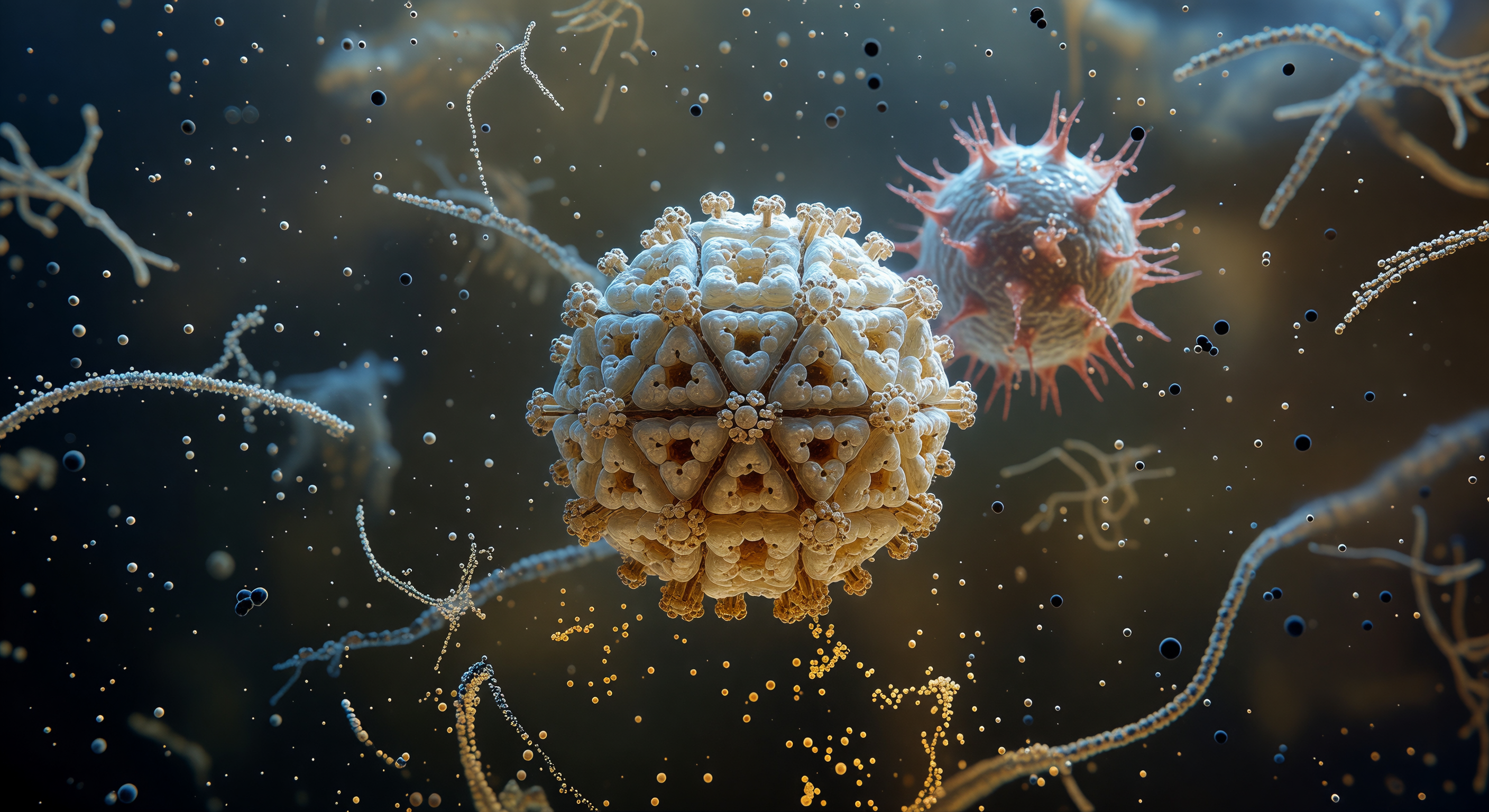
O observador encontra-se sobre a superfície exterior de um virião do SARS-CoV-2, num terreno âmbar e dourado de cabeças de fosfolípidos tão densamente compactadas que formam uma calçada fluida e ondulante, percorrida por vibrações térmicas lentas e por cunhas de colesterol cor de marfim que enrijecem localmente o pavimento viscoso. De todas as direções se erguem as proteínas spike triméricas — colunas monumentais de vermelho escuro e carmesim, enraizadas na bicamada lipídica e ascendendo vinte nanómetros acima como obeliscos retorcidos cujos sulcos longitudinais captam a luz difusa em cristas rosadas e sombras bordô profundas. No cume de cada coluna, os três domínios de ligação ao recetor abriram-se para cima em conformação ativa, como lâminas de uma turbina biológica à espera do contato com a célula hospedeira, enquanto os intervalos de quinze nanómetros entre trimeros formam avenidas entre estes pilares de arquitetura proteica, curvando-se suavemente em direção a um horizonte esférico que trai a geometria completa do virião. Lá em baixo, a membrana da célula hospedeira estende-se como uma planície cinzento-azulada coberta por uma floresta de glicanos brancos que se ramificam como cristais de gelo, e o espaço entre as duas membranas está saturado de iões e moléculas em suspensão coloidal — um meio aquoso que transforma a distância numa névoa suave e luminosa, lembrando que nada, a esta escala, existe em repouso verdadeiro.

Deitado rente à superfície da membrana externa de uma bactéria, o olhar sobe em direção a uma estrutura que ocupa o horizonte inteiro: a placa basal hexagonal do bacteriófago T4, uma coroa geométrica de sessenta nanômetros de diâmetro, suas faces facetadas faiscando em prata fria com uma luminescência azul-violeta latente onde as folhas beta empacotadas refratam o brilho difuso do ambiente aquoso circundante. Seis fibras longas da cauda se abrem em padrão radial como as patas articuladas de uma aranha mecânica colossal, cada uma dobrada em seu ponto de articulação central, com as pontas distais já pressionando a planície de lipopolissacarídeos da membrana, travando-se em receptores de açúcar específicos a centenas de nanômetros de distância — enquanto seis espigões curtos se projetam para baixo a partir da borda inferior da placa, prontos para o contato irreversível. Acima, a bainha contrátil helicoidal sobe em perspectiva de ponto de fuga, seus anéis anuais estriados desaparecendo na névoa violeta molecular antes que a cabeça icosaédrica se torne visível, apenas seu vértice inferior emergindo como um fantasma do haze. A cena inteira treme com o ruído térmico contínuo da água — cada molécula funcionando como um martelo invisível, cada superfície proteica carregada eletricamente, e o momento de encaixe definitivo entre vírus e célula hospedeira suspenso num equilíbrio que dura apenas milissegundos antes do colapso inevitável da bainha e da injeção do genoma.

Você existe dentro de uma catedral geodésica com 36 nanômetros de diâmetro, encerrado por uma abóbada contínua de proteínas que se fecha sobre si mesma sem nenhuma superfície plana — a geometria icosaédrica do capsídeo T=7 do vírus da hepatite B constrói-se a partir de 420 subunidades proteicas organizadas em capsômeros hexaméricos e pentaméricos, cada um com dobras beta-barril que sobressaem 4 a 5 nanômetros da superfície interna como formações minerais em âmbar fossilizado. Os doze vértices pentaméricos brilham com uma tensão estrutural levemente mais intensa na fosforescência difusa que emana da própria arquitetura quaternária, pontos onde a lógica geodésica fecha a esfera e a curvatura se aperta de forma quase imperceptível. No chão e elevando-se em massas emaranhadas rumo ao centro do espaço confinado, o genoma de dsRNA empacotado serpenteia em laços comprimidos de laranja queimado e cobre oxidado, suas hélices tão densamente sobrepostas que os sulcos menor e maior mal se distinguem, toda a massa tremendo com agitação térmica — microoscilações que se propagam como estremecimentos lentos através de uma mola comprimida. O espaço entre o genoma e a parede proteica não é vazio: uma névoa molecular de moléculas de água, íons magnésio e poliaminas preenche a distância intermediária, velando levemente a parede oposta do capsídeo como se vista através do mais fino vidro âmbar, enquanto os choques aleatórios do meio aquoso a 37 graus impõem sobre qualquer estrutura presente uma violência térmica contínua e omnidirecional — não o flutuar suave de um líquido, mas o tremor constante de uma sala que nunca para de sacudir.

Você existe dentro de um corredor com pouco mais de 2 nanómetros de diâmetro, uma calha proteica esculpida em anéis de proteína de placa basal cristalizada, cujas paredes se erguem a angstroms de distância em cada direção, revestidas de resíduos carregados que emitem uma luminescência violeta fria — argininas e lisinas cujas cargas positivas guiam e confinam o que está prestes a acontecer. O que acontece é isto: uma hélice de DNA de dupla cadeia irrompe por ti a velocidade catastrófica, impulsionada por seis atmosferas de pressão acumuladas na cabeça do fago acima, uma corda de vidro molecular prateado cujos sulcos major e minor passam num borrão de geometria coerente, o espaço entre a hélice e a parede reduzido a nanómetros únicos onde a água organizada treme como uma camada de Debye iridescente e instável. À tua frente, o poro de saída abre-se como uma ferida de luz azul elétrica, onde o canal perfurou a membrana interna bacteriana e os lípidos se afastaram para expor as suas caudas hidrofóbicas como uma franja dourada em torno da brecha. Para além desse limiar começa o citoplasma — uma escuridão densa e fervilhante de colisões moleculares, aquecida pelo caos térmico de um mundo um bilião de vezes mais movimentado do que qualquer intuição humana poderia antecipar — e o genoma atravessa-o agora, irreversivelmente, de uma vida para outra.

O observador encontra-se no interior de uma estrutura em formação — uma cúpula de proteínas Gag arqueia-se sobre a cabeça como a abóbada de uma catedral viva, tecida inteiramente em âmbar orgânico, cada anel hexamérico de oito nanómetros compondo um padrão de favo de mel que se curva suavemente até ao pescoço do broto onde filamentos de ESCRT-III enrolam numa trança helicoidal cobreada, como uma sutura metálica a apertar o tecido membranoso. Para além da face exterior da malha proteica, a membrana plasmática luz como um véu de âmbar quente — dois folhetos mal distinguíveis, salpicados pelos silhuetes escuros das primeiras espículas Env do vírus emergente, retroiluminados pela incandescência laranja da própria bicamada lipídica. Todo o espaço interior emana uma luz difusa e esverdeada, gerada não por qualquer fonte exterior, mas pelo brilho químico da multidão molecular que o preenche até à saturação — ribossomas escuros agrupados em cachos, filamentos de ARN que derivam como fios de seda, e uma agitação térmica perpétua que sacode cada elemento da cena num tremor baixo e incessante. A escala da experiência é inteiramente íntima e ao mesmo tempo vertiginosa: a cúpula ocupa todo o campo visual, a selva citoplasmática afunda-se abaixo em profundidades impenetráveis, e o próprio ato de brotamento — a célula hospedeira a ser esculpida de dentro para fora por uma geometria proteica que se auto-organiza com precisão termodinâmica — decorre em silêncio, como uma arquitetura que se constrói a si mesma.

Você está de pé sobre a superfície exterior de um virião de HIV-1, e o que se estende à sua volta em todas as direções é uma planície imensa e ondulante — uma savana lipídica de tons ocre e âmbar profundo, composta por fosfolipídios e ilhas de colesterol que formam um pavimento fluido e levemente luminoso, tremendo em micro-sismos incessantes gerados pelo bombardeamento browniano das moléculas de água circundantes. A membrana não é uma superfície inerte: é uma bicamada lipídica viva, com 4 a 5 nm de espessura, enriquecida em colesterol de forma deliberada pelo vírus para aumentar a rigidez e a resistência à fusão prematura, e cada cabeça de fosfolipídio aos seus pés é um objecto real com carga elétrica, rodeado por uma camada de Debye de apenas 1 nm de espessura em fluido fisiológico. A cerca de quarenta comprimentos de corpo, uma das apenas doze espículas triméricas gp120/gp41 ergue-se como um monólito basáltico — 14 nm de altura, a coroa trilobada densamente revestida de glicanos que formam uma neblina azul-esverdeada protetora, uma estratégia de camuflagem molecular que esconde os epítopos vulneráveis do reconhecimento imunitário. A extraordinária raridade destas estruturas — doze monumentos dispersos por toda a superfície esférica de aproximadamente 120 nm de diâmetro — não é um acidente evolutivo, mas uma solução de evasão imunitária: quanto menos alvos antigénicos expuser, menos anticorpos neutralizantes consegue o hospedeiro recrutar. O céu acima de si é um halo âmbar espesso de fluido extracelular, e a curvatura do mundo viral é apenas perceptível nas bordas do campo de visão, lembrando-lhe que habita uma esfera solitária e electrostaticamente carregada, suspensa entre a célula hospedeira e a extinção.
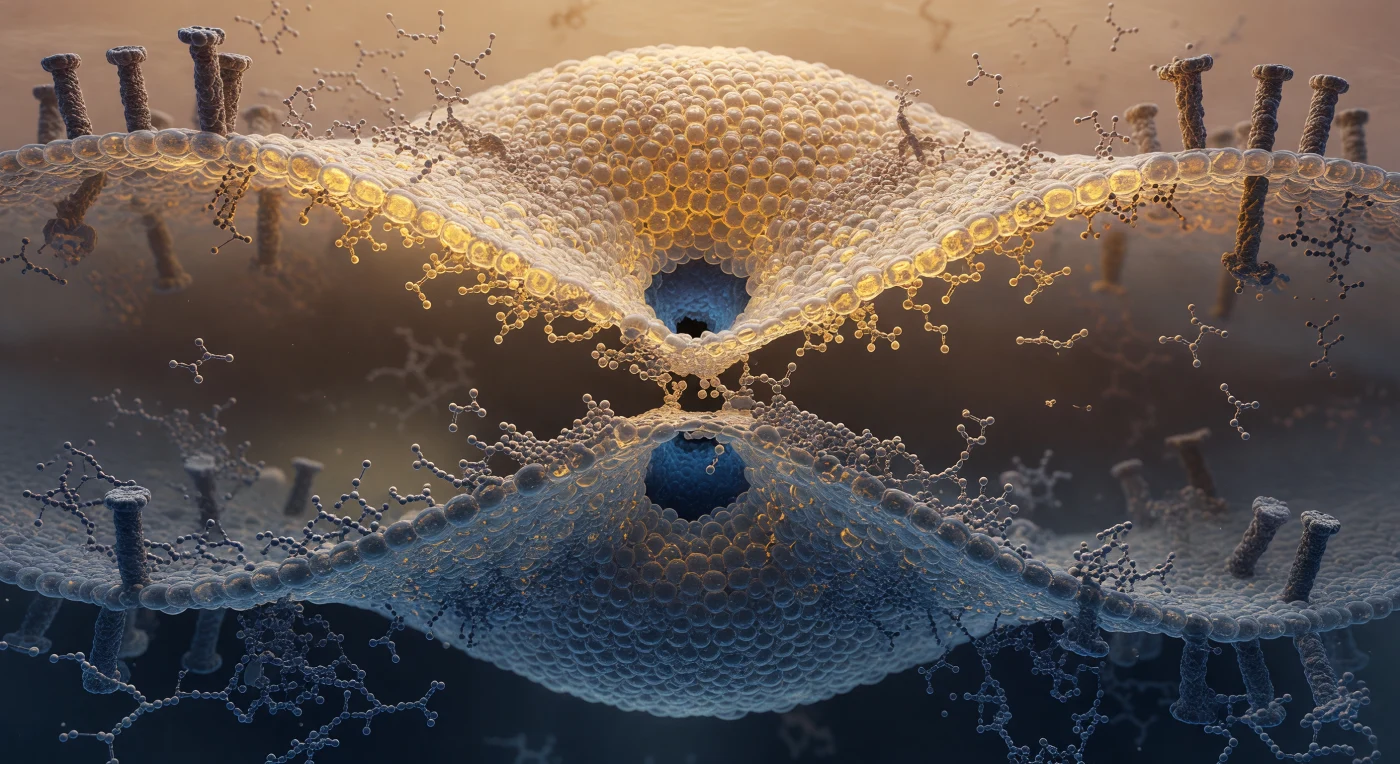
Suspenso no estreitamento de dois nanómetros que constitui o colo de uma ampulheta lipídica, o observador encontra-se no ponto exato onde duas membranas biológicas fundem os seus folhetos externos numa única monocamada contínua: acima, a membrana viral estende-se como uma abóbada de âmbar quente, os seus grupos fosfato brilhando em ocre e ouro escuro como esferas densamente compactadas do tamanho de rochas a esta escala; abaixo, a membrana endossómica replica a mesma geometria em tons de aço frio e cinzento ardósia, curvando-se para cima com uma textura molecular idêntica mas iluminada como se filtrada por água profunda. O próprio pedúnculo de hemifusão — a estrutura que o observador habita — representa um estado de transição extraordinariamente instável na via de fusão de membranas mediada por proteínas virais: os lípidos perderam aqui a sua organização em bicamada e as suas cadeias hidrofóbicas, normalmente ocultadas no interior apolar da membrana, ficam brevemente expostas numa região de geometria caótica onde os grupos fosfocolina apontam em ângulos desordenados, tremendo em micro-sacudidelas discretas à medida que a energia térmica se redistribui molécula a molécula. Ao centro do campo visual, um poro aquoso começa a abrir-se como um afinamento onde a barreira lipídica cede à pressão osmótica dos dois meios aquosos — o interior viral acima e o lúmen endossómico abaixo —, dois hemisférios escuros e levemente azulados que pressionam contra os últimos angstroms de matéria molecular antes da fusão completa que libertará o genoma viral no citoplasma da célula hospedeira.
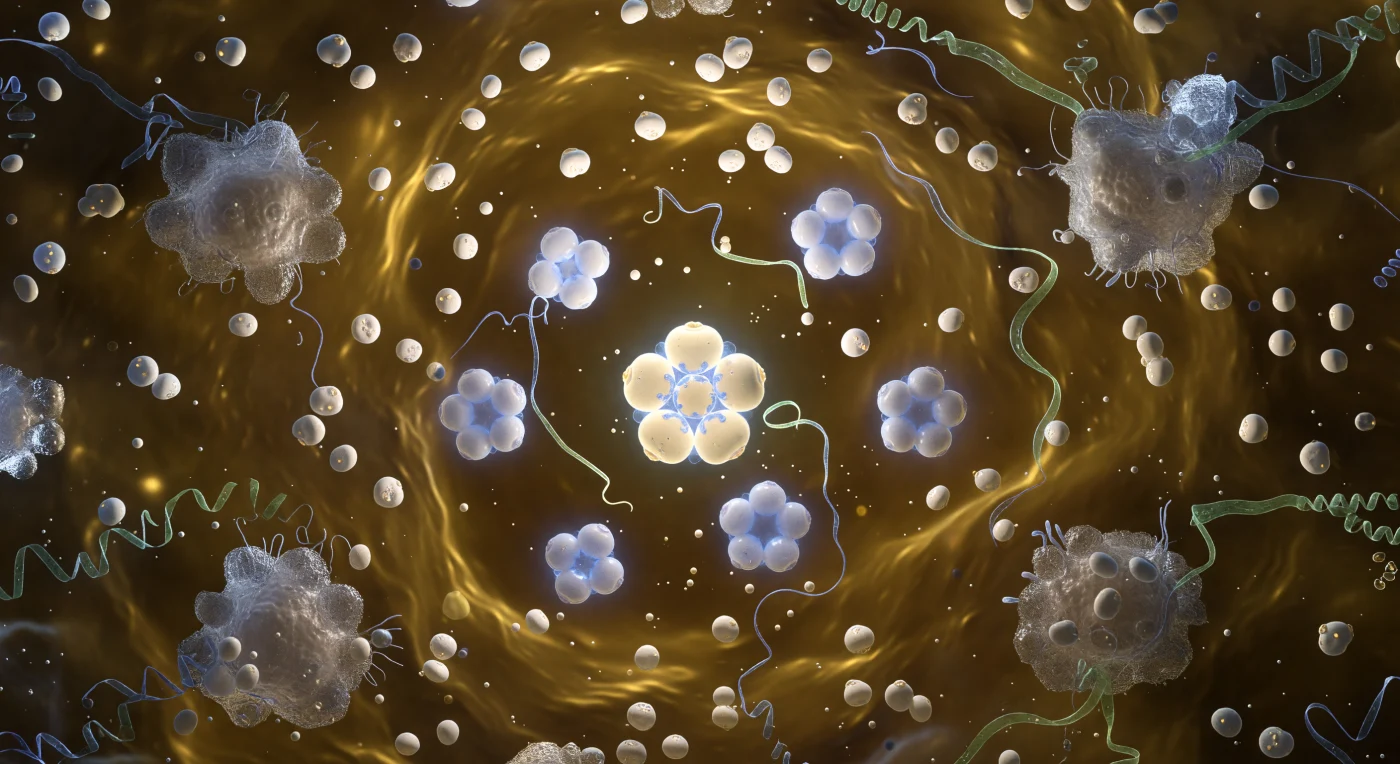

Suspenso no eixo central da haste do Vírus do Mosaico do Tabaco, o observador encontra-se envolto por um túnel cilíndrico de quatro nanómetros de diâmetro, cujas paredes se elevam em espiral dextrorsa formada por 2130 subunidades proteicas que se enroscam com uma regularidade de autómato — cada subunidade uma sanduíche compacta de folhas beta em ocre quente e âmbar, os seus bordos interlockados a ângulos precisos que conferem à parede interior a textura de uma bobine torneada em porcelana molecular. Encaixado numa calha espiral a quatro nanómetros do eixo, o genoma de RNA de cadeia simples percorre o interior como um fio de jade fosforescente, mantido em abraço electrostático pelos resíduos de arginina carregados positivamente das proteínas da cápside — uma geometria de encaixe perfeito que representa uma das mais estudadas soluções de empacotamento de ácidos nucleicos conhecidas na virologia estrutural. As paredes da haste estremeceem com vibrações picoscalares, cada subunidade a respirar e a flexionar no lugar, e a pressão osmótica e electrostática das moléculas de água e dos iões monovalentes que preenchem o canal axial cria uma tensão constante num recipiente que existe no limiar da estabilidade termodinâmica. Ao fundo do corredor — trezentos nanómetros de distância absoluta, mas à escala vivida uma perspectiva que se estende como a nave de uma catedral — a haste abre-se num portal circular de luz pálida e fria, a entrada do mundo exterior onde o movimento browniano impera e a identidade estrutural desta máquina molecular está a cada instante a ser renegociada com o ambiente.
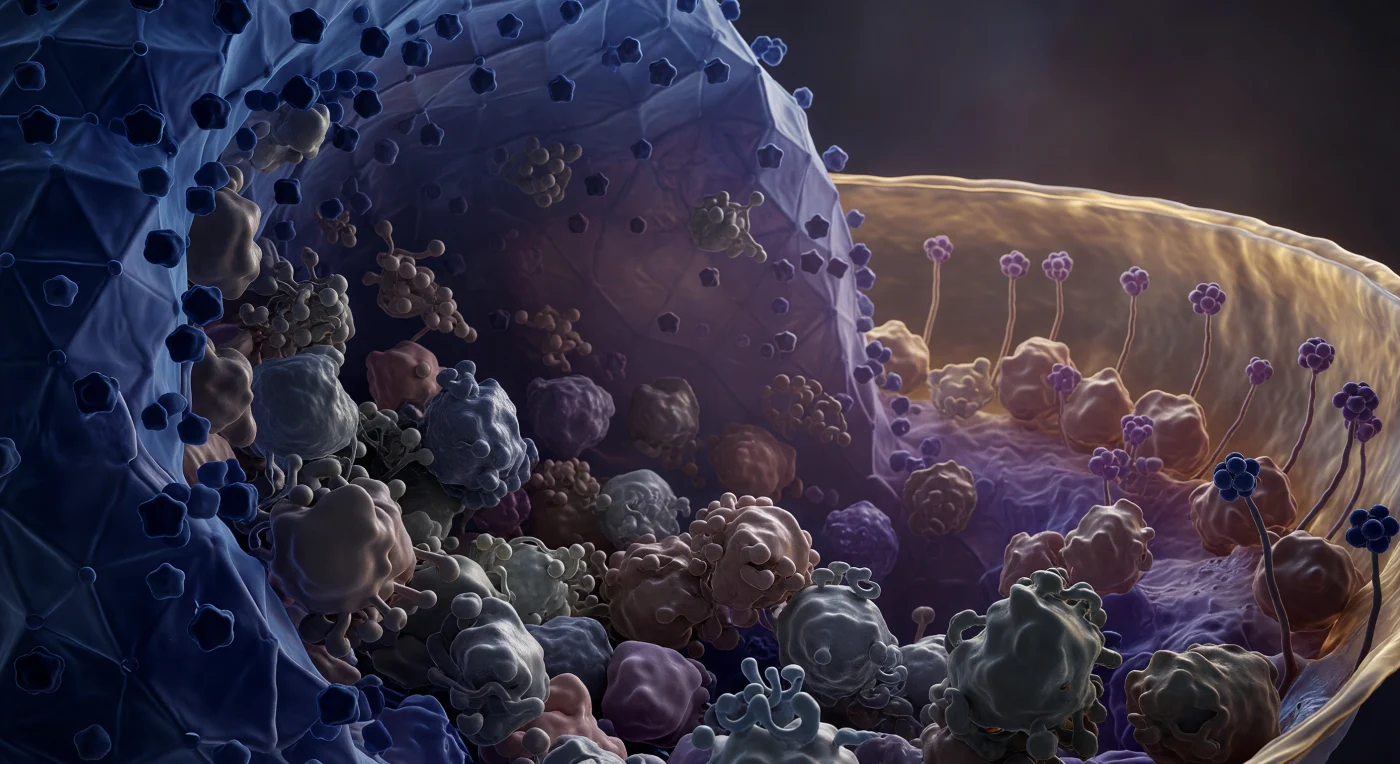
Você existe suspenso dentro de uma das arquitecturas mais densamente habitadas da natureza — o tegumento do herpesvírus, uma camada proteica amorfa comprimida entre duas paredes curvas de natureza radicalmente distinta. À sua esquerda, a parede capsidial icosaédrica ergue-se como um penhasco facetado em azul cobalto profundo, a sua geometria triangulada curvando-se para fora de vista como uma falésia geodésica de escala impossível, os vértices pentaméricos a sobressair como nódulos escurecidos numa superfície que é simultaneamente cristalina e brutalmente sólida. À sua direita, a quarenta nanómetros de distância — uma distância que aqui equivale a um horizonte — a bicamada lipídica do invólucro vibra em âmbar e ouro mel, os seus dois folhetos resolvidos como uma fina dupla banda trêmula, as hastes glicoproteicas ancoradas na sua face interna a oscilar como flora dos abissos numa corrente imperceptível. Entre estas duas arquitecturas, você está imerso no tegumento propriamente dito: VP16, UL36 e dezenas de outras proteínas pressionam-se umas contra as outras em volumes lobados irregulares de violeta acinzentado e malva empoeirado, as suas superfícies húmidas de moléculas de água ligadas, as suas fronteiras a dissolver-se onde os contactos hidrofóbicos fundem uma molécula na seguinte — uma densidade absoluta onde não existe canal aberto nem corredor vazio, cada nanómetro cúbico reclamado por algum braço ramificado ou domínio compacto. Este espaço tegumentar não é simplesmente uma camada passiva de enchimento: é uma zona de regulação dinâmica onde proteínas virais remodelem o citoesqueleto do hospedeiro, suprimem respostas imunitárias inatas e orquestram o transporte do virião ao longo dos microtúbulos — tudo isto comprimido numa espessura que a luz visível sequer consegue resolver, e que o calor molecular mantém num tremor constante e contido, cada superfície a pressionar fractalmente a sua vizinha neste silêncio mais densamente habitado que qualquer outra sala do mundo vivo.

Você deriva num labirinto viscoso de tons âmbar-esverdeados, sacudido sem cessar por impulsos térmicos que, à sua escala, equivalem a tempestades: cada molécula de água da vizinhança possui energia cinética suficiente para deslocar violentamente um virião de cem nanómetros, pois a força de Brownian motion domina completamente qualquer tentativa de movimento direcional num regime onde o número de Reynolds é infinitesimalmente pequeno. As cadeias de mucina que o rodeiam — glicoproteínas de alto peso molecular ligadas por pontes dissulfeto e interações hidrofóbicas — formam uma rede tridimensional com poros que variam entre cem e quinhentos nanómetros, criando ora corredores abertos de fluido dourado-translúcido, ora gargantas onde os filamentos quase tocam a sua cápside icosaédrica; quando o contacto acontece, resíduos de ácido siálico nas extremidades das cadeias de mucina reconhecem as proteínas de superfície do virião através de interações electrostáticas e de van der Waals, produzindo adesões transitórias que retardam a difusão e prolongam dramaticamente o tempo de travessia desta barreira química evoluída. No horizonte, a superfície epitelial emerge como uma parede planetária de membrana plasmática — uma bicamada lipídica com domínios ricos em colesterol que se reorganizam lateralmente em microssegundos — coroada pelo glicocálice, uma floresta densa de cadeias de glicano com cinco a cinquenta nanómetros de comprimento, cujos terminais de ácido siálico constituem precisamente os receptores que algumas hemaglutininas virais procuram, transformando aquela orla luminosa rosada na fronteira entre a errância no muco e o primeiro passo irreversível da infecção.

Suspenso no interior de um oceano de gelo que não é gelo — uma matriz amorfa vitrificada em milissegundos, antes que qualquer ordem cristalina pudesse formar-se — o observador encontra-se imobilizado dentro de um vácuo transparente e absoluto, cinzento-prateado em todas as direções, sem reflexo, sem refração, apenas transmissão fria e total. Emergindo desta planície estéril a intervalos irregulares, esferas colossais impõem-se com a clareza desconcertante de um contraste de densidade pura: virions icosaédricos suspensos a meio do filme vítreo, as suas superfícies revelando uma arquitetura geodésica de precisão quase perturbadora — capsómeros pentagonais e hexagonais dispostos em relevo de quatro a oito nanómetros, cada saliência e cada canyon distinguíveis como massas discretas de grafite e osso pálido, sem sombra, sem cor, apenas a gramática monótona da espessura proteica contra o gelo. Nas partículas envelopadas, espículas glicoproteicas trímeras projetam-se para o exterior como coroas de espinhos rombudos, cada um individualmente resolvido, separado do vizinho por distâncias imensas de puro vácuo vitrificado; ao fundo do campo visual, a orla do filme de carbono cai como um escarpamento geológico de negro absoluto, o limite duro de um mundo onde a violência da termodinâmica molecular foi arrestada em silêncio permanente, cada flutuação browniana congelada a meio da sua trajetória, cada ião aprisionado na posição exata em que o tempo parou.

Você está de pé sobre a superfície de uma das partículas virais mais complexas já estudadas — o poxvírus vaccinia — e o que se estende à sua frente não se parece com nenhuma outra arquitetura viral conhecida: sem simetria icosaédrica, sem geometria repetida, apenas um planalto acinzentado e assimétrico de proteínas comprimidas que se estende por 360 nanómetros como uma paisagem marciana vista ao nível dos joelhos. Sob os seus pés, a membrana externa lipoproteica encontra-se suavemente enrugada, captando uma luz bioquímica difusa e azulada nas suas dobras rasas, enquanto à frente e atrás se erguem cristas paralelas de túbulos de superfície — paredes de cordas proteicas comprimidas com seis nanómetros de altura, textura mate e granulada, da cor do estanho oxidado com leves reflexos verde-oliva — que bifurcam, incham e terminam abruptamente, deixando entre elas passagens baixas de membrana amassada onde acumulações iónicas emitem um brilho electrostático branco-azulado. A luminosidade não vem de nenhuma fonte única: é a radiância bioquímica ambiente de um meio a 37°C, que aquece todas as superfícies com um fulgor interior suave enquanto compete com o azul mais frio da carga electrostática da membrana, e acima de tudo isso essa mesma membrana externa abaulada e translúcida pende como uma lona de vidro fumado, o seu interior a cintilar como gelo glaciar pálido. É uma arquitectura primordial e crua — um dos objectos biológicos mais estranhos e sofisticados da natureza, tornado paisagem desolada de cristas proteicas e vales de membrana que se curvam até um horizonte nebuloso onde o tijolo viral cai para o vazio citoplasmático circundante.

Suspenso sobre a superfície facetada do capsídeo adenoviral, o observador contempla uma paisagem de cúpulas proteicas em marfim pálido e platina fria, cada capsômero um planalto molecular cujas arestas capturam uma luminescência difusa que emana das próprias superfícies moleculares, sem origem definida. Diretamente abaixo, os dois trilhos paralelos do microtúbulo estendem-se como cabos de uma ponte suspensa, a sua superfície resolvida em cristas de filamentos de tubulina que brilham em verde-prateado fosforescente, cada saliência de 4 nm um testemunho da precisão geométrica com que a célula organiza o seu transporte interno. Flanqueando o trajeto, complexos de dineína avançam em posturas assimétricas de esforço mecânico — formas de 15 nm em ferro escuro e cobre oxidado, os seus domínios de ligação ao cargo agarrando o ventre do capsídeo através de ligantes que pulsam em âmbar, traduzindo força molecular em deslocamento direcional com uma urgência completamente silenciosa. Em redor, o citoplasma impõe-se como uma selva de densidade esmagadora: ribossomas em bronze tarnished pressionam de todos os lados em polissomas de três a oito unidades, filamentos de actina vermelha entrelaçam-se em geometrias caóticas de coral iluminado, e ao fundo a parede curva do envelope nuclear ocupa todo o campo visual superior como um planeta próximo, as suas NPCs manchas douradas mal resolvidas na penumbra violeta que irradia do interior do núcleo.