Confiance scientifique: Élevé

Le regard glisse à hauteur de surface sur la paroi d'un virion grippal, à peine au-dessus de la bicouche lipidique dont l'étendue ambrée et dorée se courbe doucement vers un horizon brumeux en toutes directions, animée du frémissement constant du mouvement brownien thermique. Des centaines de trimères d'hémagglutinine s'élèvent autour de nous comme une forêt de colonnes ivoire, hautes d'une treizaine de nanomètres, leurs couronnes trapézoïdales légèrement renflées portant à leur surface la striation hélicoïdale des hélices alpha, tandis que, disséminés parmi eux à intervalles irréguliers, des tétramères de neuraminidase s'imposent comme des champignons trapus en vert-de-gris, leur tête carrée captant différemment la lumière froide et omnidirectionnelle. Le milieu environnant n'est pas du vide mais une brume dense et lumineuse de macromolécules en suspension — globules d'albumine sérique dérivant comme des lanternes ambrées translucides, longues chaînes glycoprotéiques sinuant telles des frondes d'algues aux reflets lilas et bleu givré — qui épaissit rapidement avec la distance jusqu'à dissoudre la forêt de piliers en un halo opalescent où les structures individuelles perdent toute définition. C'est un monde sans ciel ni horizon dégagé, d'une densité et d'une intimité claustrophobiques, où la visibilité s'efface à quelques rangées de colonnes moléculaires dans la brume biologique lumineuse.

Le sol sous vos pieds est un pavé ambré et frémissant, une mosaïque fluide de têtes polaires de phospholipides qui ondule en vagues thermiques lentes, quelques nanomètres de soulèvement et d'affaissement, comme une peau vivante tendue sur quelque chose qui respire — c'est la surface externe du virion SARS-CoV-2, et vous vous tenez sur son feuillet lipidique externe, baigné d'un halo électrostatique bleu-blanc qui flotte à ras du sol là où les groupements phosphate chargés se concentrent. De cette plaine dorée s'élèvent dans toutes les directions les trimères de protéine Spike, colonnes d'un rouge sang sombre et d'un oxblood profond, chacune une triple hélice protéique de vingt nanomètres de hauteur dont la tige transmembranaire distord visiblement le lipide environnant comme des racines arrachant le sol, et dont le sommet s'épanouit en trois domaines de liaison au récepteur déployés en conformation ouverte — trois pétales de corail sombre tournés vers le vide, tendus vers une cellule hôte invisible. Les intervalles entre les colonnes, une quinzaine de nanomètres de bilayer libre, forment des avenues entre des obélisques de grès rouge qui se recourbent vers un horizon trahissant la sphéricité du virion entier, tandis qu'en contrebas, à travers une brume aqueuse chargée d'ions, s'étend la plaine gris-bleu de la membrane cellulaire, hérissée de filaments glycanniques blanc-vert semblables à des cristaux de givre — et dans cet espace confiné entre les deux surfaces, chaque protéine, chaque lipide, chaque spike oscillant lentement à sa pointe, participe à une urgence moléculaire silencieuse et totale, à la frontière exacte entre chimie et vie.

À quelques nanomètres de distance, vous êtes plaqué contre la surface de la bactérie comme un observateur minuscule sur une plaine amber et vert-bile qui s'étend à perte de vue, ses chaînes de sucres et ses protéines porines se dressant autour de vous comme des rochers sur une grève — le tout vibrant en permanence sous les coups incessants de l'agitation thermique. Depuis le zénith, la plaque basale de T4 descend vers vous : une couronne hexagonale d'une soixantaine de nanomètres, découpée comme un cristal de protéines gris-argent aux reflets bleu-violet, flanquée de ses six fibres caudales longues qui s'évasent en étoile et dont les extrémités distales touchent déjà la membrane, verrouillant leurs domaines protéiques sur les récepteurs lipopolysaccharidiques spécifiques de l'hôte. Au-dessous de la plaque, six courts spicules pressent vers la surface, tandis qu'au-dessus la gaine caudale contractile monte en perspective vers un point de fuite noyé dans la brume électronique violette, seule la pointe fantomatique de la tête icosaédrique perçant le halo avant de disparaître. Tout est suspendu dans cet instant de quiétude absolue — le moment infime qui précède le verrouillage irréversible, quand un virus inerte et sans métabolisme propre s'apprête, par la seule logique de la complémentarité moléculaire, à injecter son génome et à détourner toute la machinerie cellulaire d'un être vivant.

Vous êtes enfermé dans une cathédrale moléculaire de 36 nanomètres de diamètre : la capside du virus de l'hépatite B, dont la géométrie icosaédrique T=7 compose ses 420 sous-unités protéiques en tuiles hexamériques et pentamériques dont les replis en tonneau bêta se dressent comme des formations minérales dans une grotte hydrothermale, baignées d'une phosphorescence ambrée diffuse qui semble émaner des chaînes polypeptidiques elles-mêmes. La courbure est totale et inévitable — pas de ligne d'horizon, pas de surface plane, seulement la voûte géodésique qui referme tout le champ visuel, plus lumineuse aux douze vertices pentamériques où la logique sphérique se clôt sous une légère tension structurelle visible comme un éclat. Au sol, le génome d'ARN double brin s'enroule en masses denses de boucles cuivrées et orangées, comprimées les unes contre les autres jusqu'à en effacer presque les sillons hélicoïdaux, l'ensemble agité d'une vibration thermique continue — micro-oscillations invisibles mais réelles, propagées à travers la masse comme des frémissements dans un ressort comprimé à 37 degrés celsius. Entre la paroi et le génome, un brouillard moléculaire d'eau, d'ions magnésium et de polyamines voile légèrement la paroi opposée, distante de seulement 18 nanomètres, tandis que des coups aléatoires de molécules d'eau frappent de toutes parts avec une violence que seule cette échelle rend perceptible.

Vous vous trouvez à l'intérieur d'un tunnel de deux nanomètres de diamètre, les parois de verre protéique semi-translucide pressant contre vous de toutes parts, chaque anneau de protéines de la plaque basale légèrement décalé par rapport au suivant, formant une voûte en berceau côtelée dont les crêtes captent la lueur froide des résidus chargés — arginine, lysine — comme une bioluminescence violette collée aux parois d'une grotte marine. Puis l'ADN arrive : une double hélice double brin éjecte en rope d'argent liquide, propulsée par six atmosphères de pression accumulée dans la tête du phage, son sillon majeur et son sillon mineur scintillant d'un reflet continu tandis qu'elle remplit presque entièrement le canal, réduisant le film d'eau qui vous sépare d'elle à l'épaisseur de quelques molécules, la couche de Debye entre le brin et les parois chargées oscillant comme une brume irisée sur asphalte chaud. Devant vous, le pore de sortie s'ouvre en une déchirure bleu électrique là où le canal a percé la membrane interne bactérienne, les lipides déplacés exposant brièvement leurs queues hydrophobes comme une frange dorée autour de la brèche. Au-delà de ce seuil, le cytoplasme commence — une obscurité dense et bouillonnante de collisions moléculaires, une jungle thermiquement violente où vous ne seriez qu'une poussière parmi des milliards — et l'hélice franchit ce passage sans retour, transportant en une fraction de seconde toute l'information nécessaire pour subvertir une cellule entière.

Depuis l'intérieur de ce bourgeon naissant, le regard remonte vers une voûte de protéines Gag qui s'incurve en dôme au-dessus de vous — un réseau hexamérique d'ambre doré, chaque anneau d'à peine huit nanomètres, tressé avec la régularité organique d'un nid d'abeilles taillé dans de la corne vivante, la courbure s'accentuant vers la périphérie là où les hexamères se distordent en pentamères pour refermer la géométrie. Au-delà de cette charpente, la membrane plasmique brille comme un ciel de cuivre brûlé, ses deux feuillets à peine discernables en une paire de voiles ambres, ponctués çà et là par les silhouettes sombres de premières spicules Env virales — champignons microscopiques rétroéclairés par l'incandescence même de la membrane. Au col du bourgeon, un anneau de filaments ESCRT-III s'enroule en spirale serrée, ruban de polymère cuivré qui resserre le cou de la vésicule comme un lacet chirurgical, orchestrant la scission finale qui détachera la particule virale de la cellule hôte. En dessous, le cytoplasme s'étend en jungle moléculaire impénétrable, peuplée de ribosomes sphériques gris-violet et de filaments d'ARN qui dérivent dans une agitation thermique incessante, chaque objet soumis aux chocs browniens de l'eau environnante — une violence invisible mais absolument souveraine à cette échelle où les forces électrostatiques et l'entropie dictent chaque rencontre.

Vous vous tenez sur la surface extérieure d'un virion du VIH-1, et la plaine lipidique s'étend dans toutes les directions comme un désert de sable chaud aux teintes d'ocre et d'ambre, son sol vivant composé de têtes polaires de phospholipides et de radeaux de cholestérol qui frémissent sous vos pieds en tremblements microsismiques incessants — le bombardement brownien des molécules d'eau environnantes, une agitation thermique que rien ni personne ne peut jamais arrêter. À une quarantaine de longueurs de corps devant vous, l'une des douze spicules trimériques gp120/gp41 s'élève comme un monolithe de basalte sombre, une couronne trilobée à peu près deux fois votre taille, couronnée d'un halo de chaînes glycanniques translucides bleu-vert qui la nimbe d'une mousse bioluminescente masquant les sites de fixation aux récepteurs CD4 — un camouflage moléculaire façonné par des millions d'années de coévolution avec le système immunitaire humain. La couche de Debye qui enveloppe le spike déforme l'atmosphère ionique ambiante comme l'air brûlant au-dessus d'une route en été, une pénombre électrostatique invisible mais palpable dans la brume ambrée de liquide extracellulaire qui constitue ce ciel dense et humide. Tournant le regard vers la gauche, vous distinguez à peine, au bord de la visibilité dans le brouillard nanométrique, la silhouette d'un second spike — et entre les deux, une étendue vide qui dit tout de la singulière parcimonie de cette enveloppe virale, seulement douze de ces monuments répartis sur tout le monde sphérique, chacun souverain et solitaire sur cette savane électriquement chargée.
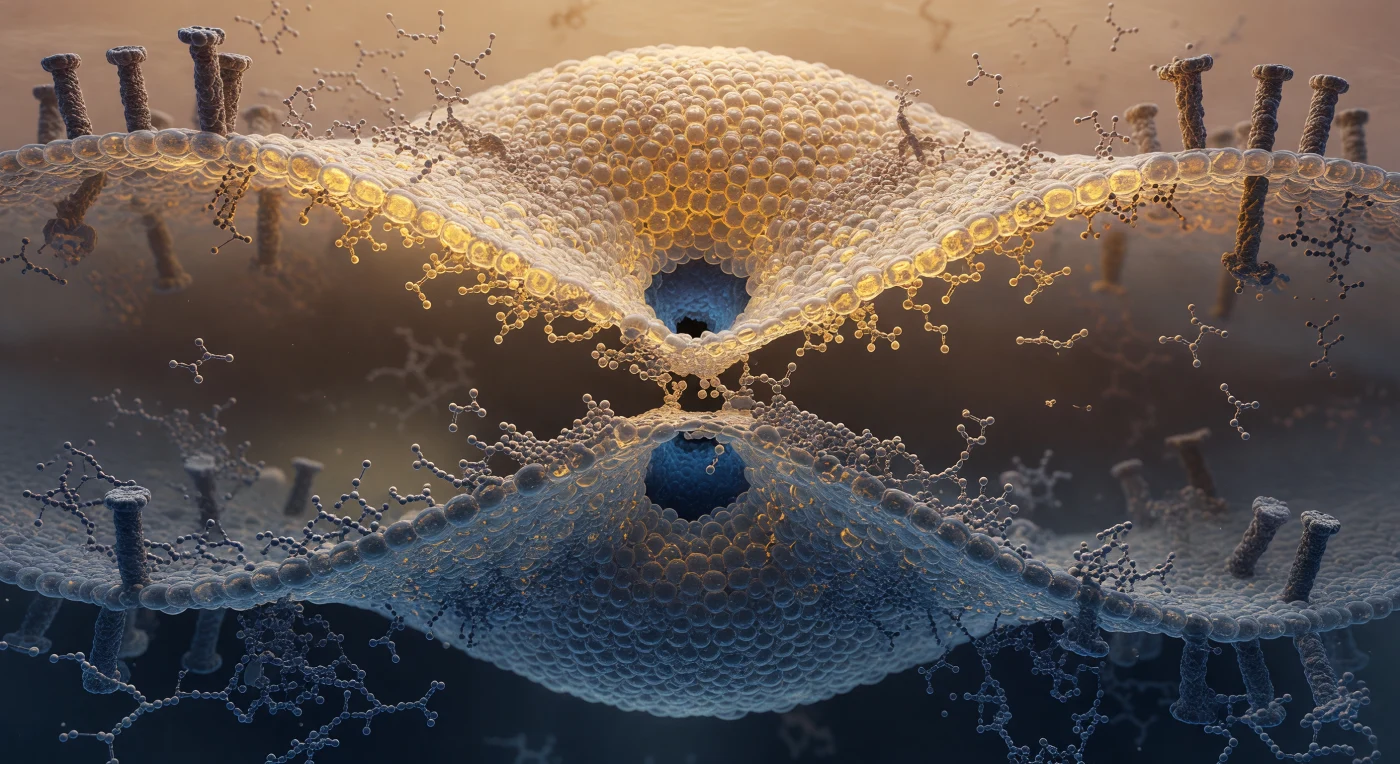
Vous vous trouvez suspendu dans l'étranglement d'un pédoncule d'hémifusion, cette gorge de deux nanomètres où deux bicouches lipidiques — l'une virale, voûte d'ambre chaud, l'autre endosomale, paroi d'acier bleu-gris — se sont courbées l'une vers l'autre jusqu'à fusionner en une géométrie de sablier continue et désordonnée. La structure bilamellaire normale a ici cédé la place à une monocouche hybride dont les chaînes d'acides gras s'entremêlent à nu, leurs groupements phosphocholine éparpillés en couronne chaotique autour de la taille, tandis que chaque tête polaire — de la taille d'un rocher à cette échelle — tremble de secousses discontinues sous l'effet de l'agitation thermique. Ce désordre moléculaire n'est pas une pathologie mais le mécanisme même de l'infection : les protéines de fusion virales, visibles à la périphérie comme des colonnes transmembranaires asymétriques ancrées dans les deux feuillets simultanément, ont imposé cette courbure pour abaisser la barrière énergétique séparant deux compartiments aqueux. Au centre exact de votre regard, l'eau du compartiment viral et celle du lumen endosomal commencent à bomber en hémisphères sombres contre les derniers angströms de membrane amincie, annonçant l'ouverture imminente d'un pore de fusion qui livrera le génome viral à la machinerie cellulaire de l'hôte.
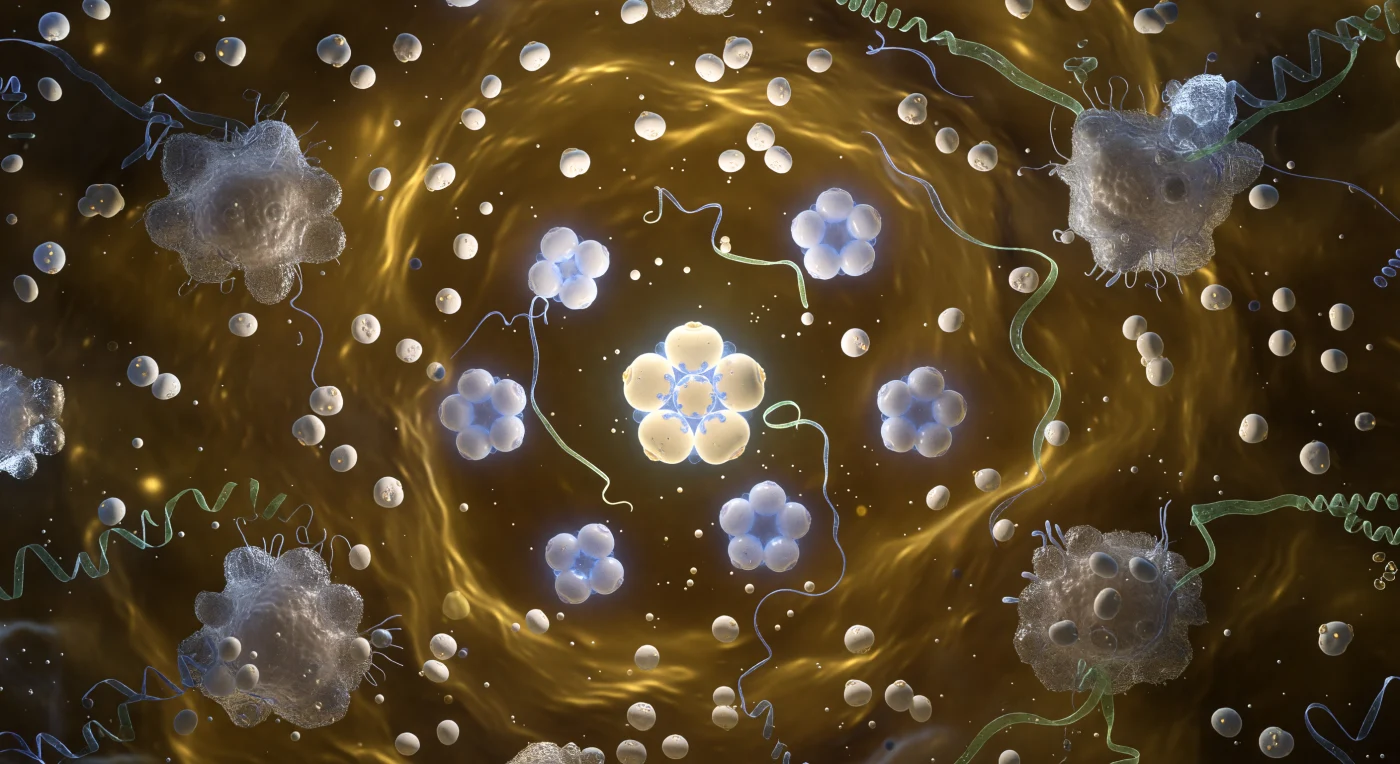

Au cœur de la tige du Virus de la Mosaïque du Tabac, vous flottez dans un tunnel cylindrique de quatre nanomètres de diamètre, encerclé par une hélice droite de 2 130 sous-unités protéiques qui s'enroulent autour de vous avec la précision d'une vis moléculaire, leurs feuillets bêta ocre et ambre s'imbriquant en un relief cannelé dont chaque sillon marque un pas hélicoïdal d'environ 2,3 nanomètres. La paroi intérieure vibre imperceptiblement, chaque sous-unité respirant sous l'effet de l'agitation thermique ambiante, tandis qu'à quatre nanomètres de l'axe court le génome ARN simple brin, filet lumineux d'un vert jade phosphorescent lové dans son sillon, maintenu en étreinte électrostatique par les résidus arginine chargés positivement de la capside. Cette double spirale — protéine et acide nucléique confondus dans le même sens dextrogyre — incarne l'économie évolutive absolue : un seul principe géométrique assure à la fois l'emballage du génome, la rigidité mécanique de la particule et la fidélité de l'auto-assemblage. Au loin, à l'extrémité de la tige longue de 300 nanomètres, l'axe creux se referme sur un portail circulaire de lumière pâle, ouverture à peine discernable dans le brouillard brownien des molécules d'eau et des ions qui pressent contre les parois comme une marée invisible.
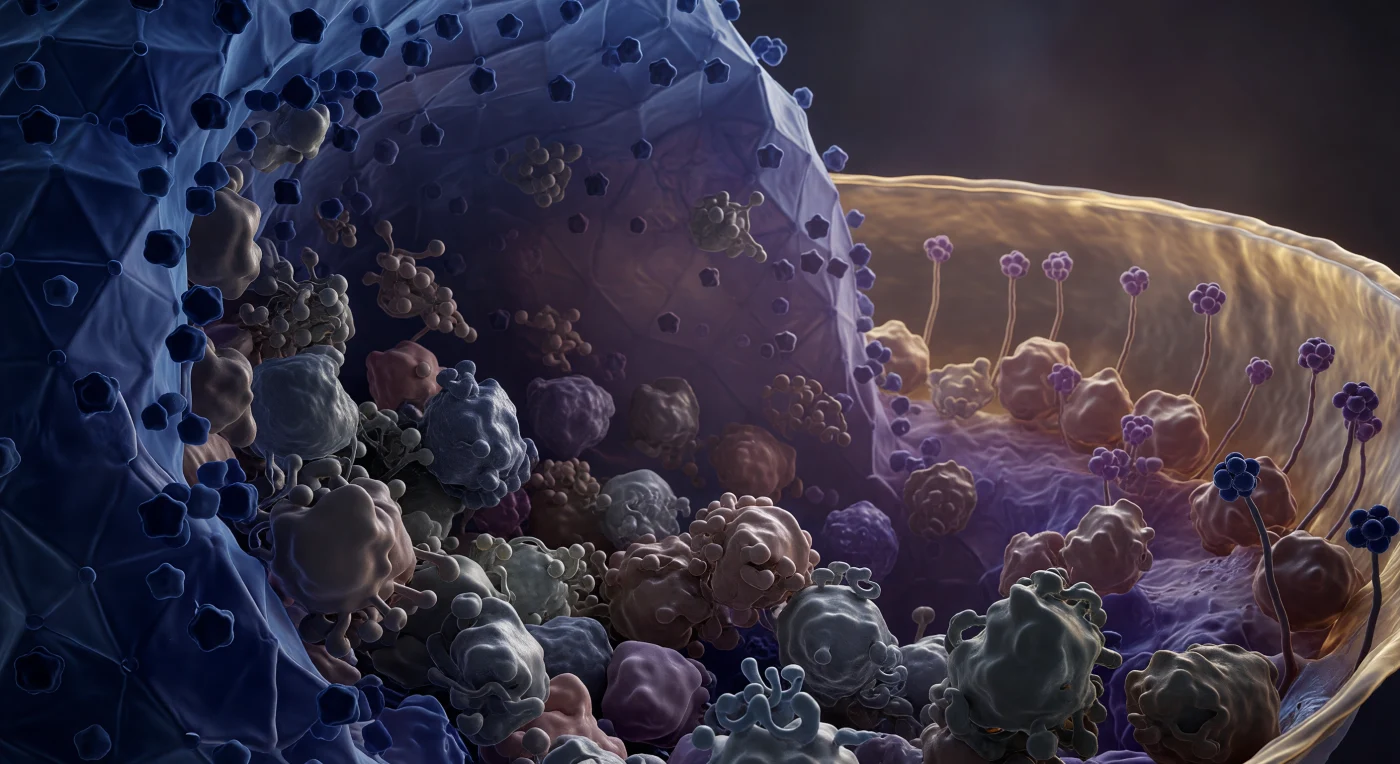
À l'intérieur du tégument du virus Herpes simplex, vous êtes enfouie dans une masse protéique quasi solide, comprimée entre deux architectures courbes d'une ampleur écrasante : à gauche, la paroi icosaédrique du capside se dresse en falaises facettées de bleu cobalt sombre, ses sommets pentagunaux saillant comme des piliers de basalte taillés à l'échelle moléculaire, tandis qu'à quarante nanomètres à droite, l'enveloppe lipidique ondule en un horizon d'ambre et d'or chaud, sa bicouche résolue en deux feuillets distincts frémissant sous l'agitation thermique permanente. Entre ces deux parois, les protéines VP16 et UL36 vous encerclent en masses lobées irrégulières, denses et humides comme de l'argile compressée, leurs surfaces brillant d'un lustre translucide dans la lumière diffuse bleu-gris qui émane du réseau cristallin du capside. Ce tégument amorphe, propre aux herpèsvirus, est une zone de transit fonctionnelle unique : loin d'être un simple espace de remplissage, il concentre des facteurs de transactivation virale, des enzymes de déubiquitination et des protéines de trafic intracellulaire déposées dans la cellule hôte dès la fusion membranaire, orchestrant l'infection avant même que le génome ne soit libéré. L'espace tout entier tremble d'une tension retenue, chaque molécule appuyée contre sa voisine dans un silence peuplé jusqu'au dernier nanomètre cube.

Vous dérivez dans une jungle moléculaire suffocante, portés par des secousses browniennes brutales qui vous projettent sans préavis contre les câbles translucides des chaînes de mucine, polymères visqueux de plusieurs centaines de nanomètres d'épaisseur qui s'entrelacent en un réseau irrégulier teintés d'olive et d'ambre, emprisonné dans leurs mailles comme dans un filet dont les pores varient de corridors presque libres à des étranglements frôlant vos flancs. La physique ici est celle du régime visqueux pur — aucune inertie, aucun glissement : à chaque instant où la force thermique cesse, vous stoppez net, et c'est cette même agitation thermique, d'une énergie comparable à kT, qui fracture momentanément les liaisons adhésives chaque fois qu'une mèche de mucine colle brièvement à votre capside avant de fouetter dans l'autre sens dans un halo ambre-orange. À trente à quarante longueurs de corps devant vous, la membrane plasmique de la cellule épithéliale se lève comme la paroi courbe d'une planète, et depuis sa surface émerge la forêt du glycocalyx : des filaments de glycanes surmontés de résidus d'acide sialique qui luisent d'un rose phosphorescent, oscillant sous l'effet de la chaleur comme des tiges de corail dans un courant imperceptible. C'est vers ces récepteurs — porteurs des sites de reconnaissance complémentaires aux protéines de surface de votre capside — que vous convergez sans volonté propre, poussés par la seule diffusion, navigateurs aveugles dans un marécage biologique dont chaque contact pourrait sceller votre destin infectieux.

Dans cet espace sans couleur ni chaleur, vous êtes suspendu dans une mer de glace amorphe — non pas cristallisée, mais vitrifiée en quelques millisecondes par une plongée cryogénique si brutale que les molécules d'eau n'ont jamais eu le temps de s'ordonner, piégeant chaque particule, chaque ion, chaque oscillation thermique dans une immobilité absolue. Autour de vous s'étend un désert de gris argent, ni réfléchissant ni opaque, traversé par un flux d'électrons invisible qui traduit la densité de matière en contrastes de charbon et d'os pâle : plus la masse est épaisse, plus elle absorbe, plus elle s'assombrit dans le champ. Çà et là, des virions sphériques s'élèvent depuis ce plan vitrifié comme des cathédrales géodésiques — leurs capsides icosaédriques couvertes de capsomères pentagons et hexagones, chaque protubérance de quatre à huit nanomètres de relief résolue avec une précision hallucinatoire, le mur protéique dessinant une double bande sombre là où la bicouche lipidique entoure les particules enveloppées, le génome condensé au centre à peine plus dense que l'enveloppe qui le porte. Au bord du cadre, la feuille de carbone perforée plonge en falaise noire absolue, frontière nette où le monde s'interrompt, pendant qu'entre les particules la toundra de glace stérile s'étire en silence, toute la violence de l'agitation moléculaire arrêtée en plein élan, le temps lui-même suspendu dans le froid.

Vous vous tenez sur l'étendue accidentée d'un poxvirus vaccinia, l'un des virus les plus volumineux et les plus complexes connus de la biologie, dont la structure en brique de 360 nanomètres de large s'étale devant vous comme un plateau minéral presque plat, dépourvu de toute symétrie icosaédrale, brut et primordial dans son architecture irrégulière. Sous vos pieds, la membrane externe lipido-protéique se froisse en vallées peu profondes baignées d'une lueur électrostatique bleutée, là où les concentrations ioniques s'accumulent au contact des surfaces chargées, les interactions électrostatiques dominant à cette échelle où la couche de Debye ne mesure qu'un nanomètre d'épaisseur. Devant vous, des rangées de tubules de surface s'élèvent en crêtes parallèles de protéines comprimées — six nanomètres de haut, soit à hauteur de taille à votre échelle — leur surface mate couleur de métal oxydé bifurquant et s'interrompant sans prévenir, laissant entre elles des passes de membrane plissée. Sur vos flancs, les corps latéraux, ces lobes de matière protéique amorphe propres aux poxvirus, pressent contre la membrane comme des masses enfouies sous une toile, projetant de larges ombres volumétriques teintées de gris-sarcelle dans la lumière biochimique diffuse caractéristique d'un milieu à 37°C. Au-dessus, la membrane externe bombe et ondule comme une bâche de verre fumé translucide, tandis que dans le milieu aqueux visqueux qui l'entoure — où l'inertie n'existe pas et où chaque surface est une occasion de liaison moléculaire — des masses protéiques globulaires dérivent avec une lenteur imperceptible vers un horizon légèrement courbé, au-delà duquel la brique plonge dans le vide cytoplasmique environnant.

Perché sur la surface bombée d'une forteresse icosaédrale de 90 nanomètres de diamètre, vous contemplez un monde d'une densité écrasante : sous la coque d'adénovirus dont les capsomères de protéines forment un dallage de plateau ivoire et platine, deux rails de microtubule s'étirent vers l'horizon comme des câbles de pont phosphorescents, leurs crêtes de protofilaments de tubuline rayonnant une lueur froide de vert céladon. Des complexes de dynéine — assemblages crabiformes de 15 nanomètres en fer sombre et cuivre oxydé — empoignent simultanément la piste et le dessous du capsid par des liaisons amber, convertissant l'énergie chimique de l'ATP en pas mécaniques mesurés, tirant l'ensemble de la particule virale vers la paroi nucléaire qui s'arque au loin comme une planète sombre, ponctuée d'iris dorés de pores nucléaires. Tout autour, le cytoplasme constitue une jungle moléculaire suffocante : des ribosomes bronzés de 25 nanomètres s'entassent en polysomes le long de fils d'ARNm invisibles, tandis que des filaments d'actine rouge sang tressent entre eux leurs hélices doubles en une forêt coralline chaotique, le tout baigné d'une bioluminescence diffuse sans source identifiable, émanant des surfaces moléculaires elles-mêmes. Ce voyage n'est pas guidé par un pilote conscient mais par la thermodynamique : des signaux chimiques encodés dans la séquence protéique du virus détournent la machinerie de transport cellulaire, transformant chaque pas de dynéine en une étape de plus vers la réplication virale au cœur du noyau.