
Você flutua na borda de um canyon espiral vivo, as paredes da goteira oral de um *Paramecium caudatum* arqueando-se em ambas as direções como um anfiteatro esculpido em vidro biológico, tudo banhado pelo contraste de fase que transforma cada superfície em prata luminosa contra o negro absoluto. Milhares de membranelas ciliares alinhadas nas paredes do sulco batem em ondas metacronais sincronizadas — vagas sucessivas que percorrem a parede como vento atravessando um campo de trigo prateado — gerando uma corrente de vórtice que arrasta bactérias em forma de bastonete, dourado-translúcidas pela refração da luz através de suas membranas lipídicas, em espiral descendente em direção ao poço citosomal, uma abertura de escuridão pulsante onde a membrana treme com o trabalho contínuo de engolfamento. A uma escala onde a água se comporta como mel e a inércia não existe, todo o movimento é dominado pela viscosidade e pela mecânica de superfície: cada batida ciliar, a cada vinte a quarenta hertz, é um ato de força bruta contra um fluido que resiste como massa viva. Acima de você, a parede celular translúcida arqueia-se como uma cúpula de vidro fosco e, através dela, o macronúcleo brilha em âmbar quente — uma presença em forma de rim pressionada contra o ectoplasma — enquanto no interior escuro da célula vacúolos digestivos derivam lentamente, cada um uma esfera selada em diferente estágio de dissolução enzimática, testemunhos silenciosos de capturas anteriores neste mesmo vórtice.

Você paira imóvel num meio que pressiona suavemente em todas as direções como glicerina morna, suspenso abaixo de uma congregação de *Euglena viridis* que sobe em coluna espiral em direção à luz difusa da superfície. Cada célula — um fuso de sessenta micrômetros envolto em película de proteínas que lhe dá brilho sedoso e metálico — carrega no seu interior fitas de cloroplasto enroladas em hélice de vidro jade, captando a claridade descendente e devolvendo-a como fagulhas de esmeralda e chartreuse que se multiplicam célula após célula até o topo da coluna. Perto do polo anterior de cada organismo arde um estigma minúsculo — uma mota terracota de menos de um quarto de micrômetro, fotorreceptor primitivo que orienta cada célula em fototaxia positiva, transformando a coluna inteira numa chaminé viva e ordenada que sobe pelo escuro oliváceo como fumaça com inteligência química. O meio em si não é transparente: está tingido de amarelo-esverdeado pelo vazamento coletivo de fluorescência de clorofila excitada pela luz filtrada lá no alto, e dentro desse halo lime partículas de detritos orgânicos e bactérias invisíveis tremem em agitação browniana como pó de prata suspenso no tempo. Abaixo de você, o verde-limão aprofunda-se em oliva e depois em azul-escuro impenetrável, onde nenhuma Euglena sobe e o líquido se torna pesado e quieto, lembrando que esta catedral de luz viva tem um chão de sombra.

Deitado contra o mosaico de sílica fragmentada — os restos de frústulas de diatomáceas cujas redes hexagonais de poros dispersam a luz como vitrais partidos enterrados no sedimento escuro — o olhar sobe em direção a um evento arquitectónico que domina o mundo inteiro: o pseudópode de *Amoeba proteus*, erguendo-se como uma geleira esculpida em vidro vivo, a sua capa hialina dianteira opticamente imaculada, curvando a luz ambiente em arcos dourados que varrem o substrato à sua frente como caustics solares a anunciar a passagem de uma nuvem. Dentro do tubo ectoplásmico transparente, o endoplasma avança em torrente — um rio denso e granular de bronze líquido e âmbar profundo, carregado de vacúolos alimentares cor de mel fumado, mitocôndrias que cintilam como fragmentos de cobre, e grânulos refractivos que tombam no fluxo como pedras numa cheia, tudo impulsionado pela conversão em tempo real de sol fluido em gel estrutural na fronteira sol-gel, onde franjas de interferência iridescente em lavanda pálida e ouro suave ripam como óleo sobre água. Este movimento lento — de 1 a 10 µm por minuto, governado por viscosidade e tensão superficial em vez de inércia — é uma das formas mais antigas de motilidade eucariota, um fluxo citoplasmático dirigido pela polimerização de actina na margem dianteira que transforma a própria substância da célula numa arquitectura em avanço contínuo. Mais atrás, o corpo da ameba recua numa montanha de citoplasma âmbar trémulo, cujos contornos distantes se dissolvem na névoa castanha-esverdeada da coluna de água, onde hastes bacterianas invisíveis derivam agitadas pelo movimento browniano, e toda a cena respira com a luz íntima e dourada de uma única célula a devorar o tempo.
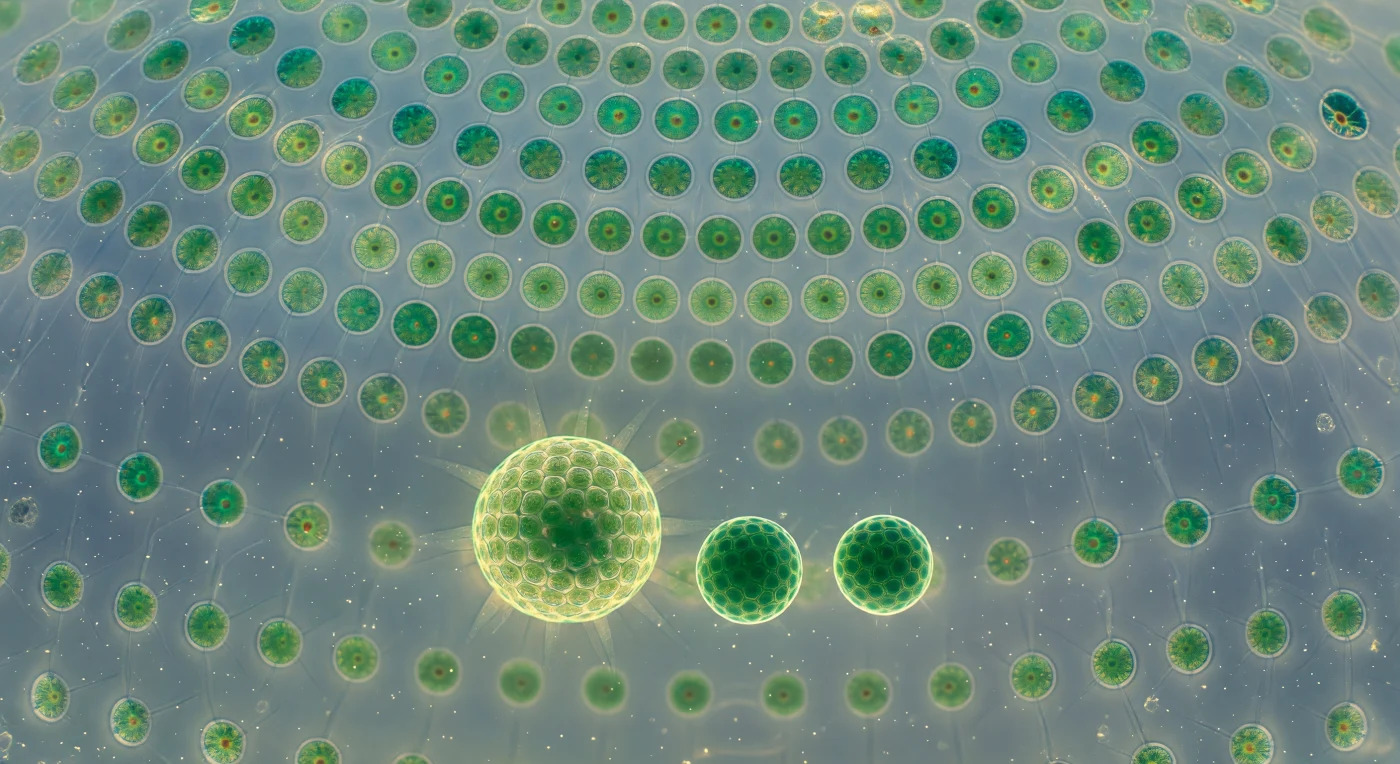
O observador encontra-se suspenso no centro de uma esfera viva com cerca de meio milímetro de diâmetro — *Volvox globator*, uma colónia de milhares de células biflageladas encravadas numa matriz de glicoproteínas transparente como vidro, ligadas entre si por filamentos citoplasmáticos tão finos que só se revelam como prateados quando a luz os atravessa em ângulo certo, tecendo uma abóbada geodésica que se curva acima e em redor como a nave de uma catedral. Cada célula somática tem o seu próprio cloroplasto de verde esmeralda e um ocelo cor de ferrugem — um organelo fotorreceptor minúsculo que arde como uma brasa no interior de cada corpo celular — e os seus flagelos batem em ondas metacrónicas coordenadas que percorrem a superfície interior da esfera como vento sobre um campo de musgo luminoso, fazendo rodar lentamente toda a colónia na coluna de água. No fluido aquoso interior flutuam três colónias-filha em diferentes estádios de maturação: a maior já possui a sua própria cúpula geodésica completa e cintilante, a segunda é mais densa e opaca, saturada de citoplasma rico em reservas, e a menor não passa ainda de uma bola oca de células verde-uniforme com os flagelos ainda sem função, cada uma suspensa no meio líquido com a precisão quieta de um planeta dentro de um relicário de cristal. Esta é a condição de organismo colonial nas suas fronteiras mais ambíguas — uma entidade onde a divisão entre indivíduo e comunidade dissolve-se na geometria, onde cada célula é autónoma na sua bioquímica mas irreversivelmente integrada na coordenação hidrodinâmica e fotossensorial do conjunto. A luz difusa que penetra pela água da lagoa atravessa a parede de glicoproteínas e espalha-se pelo interior como uma luminescência azul-esverdeada uniforme, sem sombras duras, apenas a profundidade suave de um espaço inteiramente vivo.

O abismo oceânico envolve tudo com um peso e uma textura que a palavra "escuridão" não consegue conter — é uma negridão com densidade química, com o leve arrasto de correntes térmicas e flocos de neve marinha que descem silenciosamente como tecido translúcido numa catedral sem luz. Então uma detonação de fogo frio a 490 nanómetros rasga o nada: um *Noctiluca scintillans* revela-se por uma décima de segundo como uma esfera gelatinosa quase perfeita, a sua membrana plasmática curvando a própria luz emitida enquanto a vacuola central ocupa a maior parte do volume e os pequenos organelos carregados de luciferina ainda transitam de azul-branco para um verde-teal profundo enquanto a reação química esgota o seu substrato. A descarga resulta de uma perturbação mecânica na água — o menor turbilhão, a passagem de um fragmento de mucilagem — que ativa scintilões, vesículas citoplasmáticas onde a luciferina reage com a luciferase num clarão de quimioluminescência fria, sem calor, sem custo térmico, pura eletroquímica marinha. Uma segunda célula dispara três comprimentos de célula mais longe, depois uma terceira atrás dela, a reação em cadeia propagando-se pela água negra em anéis sobrepostos de luz glacial, cada flash imprimindo a sua pós-imagem azul-esverdeada antes de a escuridão absoluta reivindicar tudo de volta como se a luz nunca tivesse existido.

Olhamos diretamente para baixo, suspensos sobre o abismo azul-cobalto que é o disco oral de um *Stentor coeruleus* — uma espiral viva de membranelas compostas que agita a água em redor num giro lento e hipnótico, sugando bactérias e algas para o funil citostomático como uma drenagem gravitacional de arquitetura biológica. O bordo do disco pulsa com iridescência azul-prussiana, os grânulos de estentorína distribuídos em estriações paralelas tão finas sob a película que se lêem como um têxtil metálico, alternando aquamarina pálida e índigo profundo. Através da película semitransparente do corpo trombeta que se estende abaixo, o macronúcleo revela-se como um rosário de nódulos opalescentes — um fio de pérolas fluviais suspenso no citoplasma granuloso, enquanto vacúolos alimentares âmbar e castanho-escuro derivam na corrente endoplasmática em diferentes fases de digestão. O meio aquoso não é vazio mas uma névoa âmbar subtilmente texturada, carregada de matéria orgânica em suspensão que difunde a luz transmitida como vidro fosco aquecido, dando à cena a qualidade de um instante geológico congelado enquanto a corrente espiral que o gerou continua o seu curso ancestral.
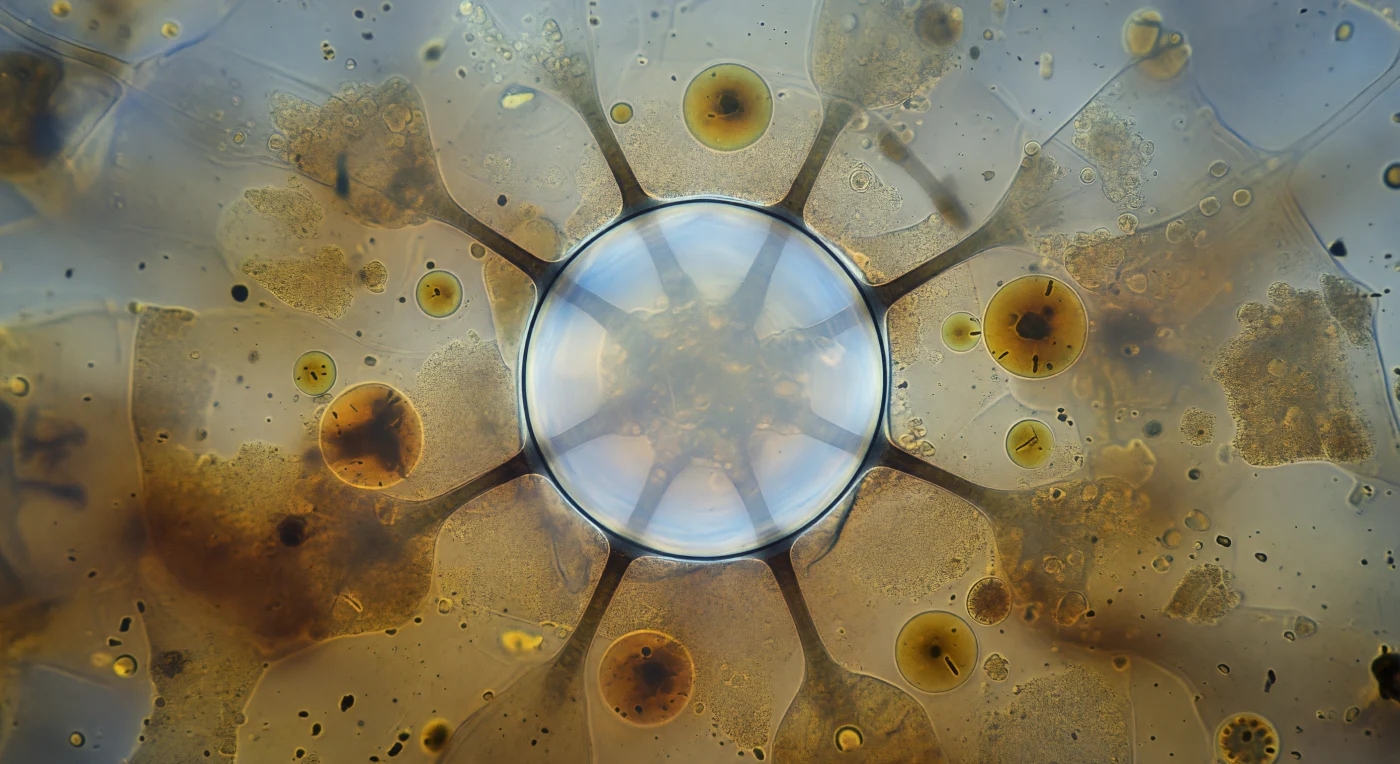
O vacúolo contrátil domina o campo visual como uma cúpula perfeita de vidro vivo — uma esfera de trinta micrómetros cujas paredes são tão finas que parecem menos uma fronteira do que uma tensão congelada, o seu interior brilhando com uma luminescência aquosa pálida que trai o peso osmótico de centenas de segundos de água acumulada. Seis túbulos nefridiais irradia em torno do equador da esfera como raios mergulhados na névoa do citoplasma, estruturas tubulares de membrana especializada que conduzem continuamente a água osmótica para o interior da organela, funcionando como um sistema coletivo de drenagem activo contra um gradiente de pressão permanente — pois o *Paramecium*, habitando meios hipotónicos, enfrenta uma entrada de água constante pela osmose e expulsaria por colapso celular se este ciclo parasse. Os vacúolos alimentares — esferas âmbar irregulares de cinco a quinze micrómetros que derivam pelo citoplasma circundante — contêm resíduos de bactérias em digestão lisossomial, os seus interiores a escurecer da periferia para o centro onde as enzimas hidrolíticas já transformaram organismos inteiros em gradientes de ocre e castanho. A cena inteira está suspensa no milissegundo anterior ao colapso total: a membrana no seu limite absoluto de curvatura, o campo de citoplasma granular pressionando de todos os lados, o mundo interior do *Paramecium* a vibrar com a tensão contida de um sistema prestes a libertar toda a sua carga acumulada numa única contracção explosiva e involuntária.
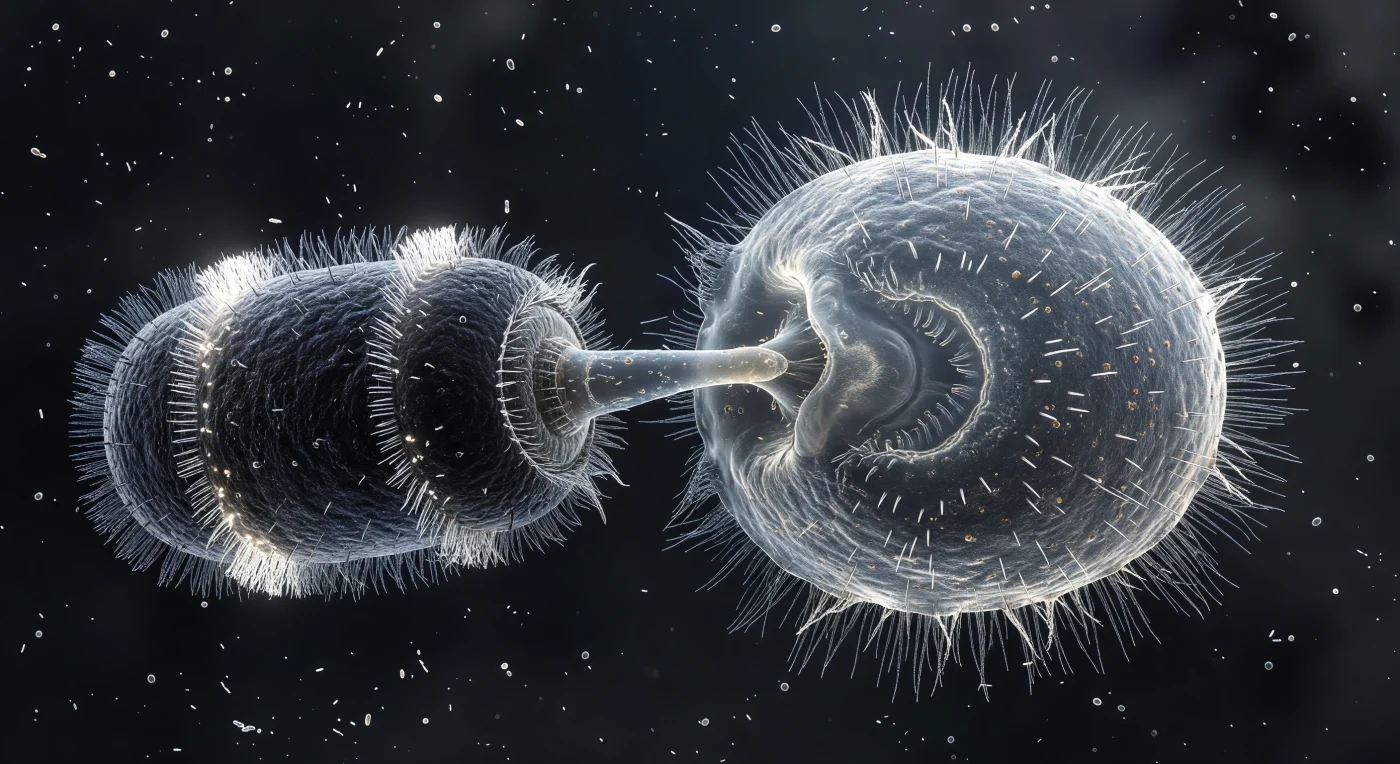
O cilindro escuro do Didinium ocupa a cena como uma máquina de guerra biológica, os seus dois coronéis ciliares brilhando em prata-branca como halos congelados num medium aquoso onde a inércia não existe e cada movimento é governado pela viscosidade. Da extremidade anterior, o probóscide totalmente estendido pressiona a película do Paramecium, criando uma côncavidade de vidro esticado que capta a luz fria do fundo e marca o epicentro de um ato de predação que dura apenas milissegundos — o Didinium é um especialista obrigatório que se alimenta quase exclusivamente de Paramecium, implantando nematocistos do probóscide para ancorar e paralisar a presa antes de a engolir inteiramente, mesmo quando esta é consideravelmente maior do que ele. O Paramecium responde com a única defesa que possui em tempo real: a descarga explosiva e simultânea dos seus tricocistos, filamentos de proteína cristalina lançados para o exterior formando uma coroa densa de fios brancos que se estendem pelo meio circundante como vidro fiado em explosão lenta, uma resposta que pode repelir alguns predadores mas que, contra o Didinium, raramente altera o desfecho. A inversão ciliar do Paramecium — a sua resposta de fuga clássica, um recuo de 10 a 200 milissegundos desencadeado por influxo de cálcio — está visivelmente fragmentada, o padrão metacronal normal desintegrado numa franja prateada e desordenada de batimentos conflituosos, enquanto o meio envolvente, uma névoa de bactérias a diferentes planos focais, recorda que estes dois organismos, medidos em dezenas a centenas de micrómetros, habitam um universo tridimensional onde os vizinhos invisíveis são a maioria.

Hoveres acima de um fundo marinho silto-arenoso banhado por uma luz azul-esverdeada que parece emanar da própria água, o que se ergue à frente não é uma rocha nem um grão de areia, mas um organismo vivo — a carapaça calcária de *Ammonia tepida*, um foraminífero bentónico cujas câmaras globosas, empilhadas em espiral logarítmica, brilham como mármore polido, cada parede de calcite translúcida o suficiente para deixar adivinhar a sombra tênue do citoplasma interior. A partir da abertura umbilical, quase invisível contra a névoa ambiente, irradia uma rede de reticulópodos — filamentos anastomosados tão finos como fios de vidro molhado, traídos apenas pelo tráfego bidirecional de grânulos âmbar que fluem ao longo das suas superfícies em correntes contínuas, transportando nutrientes para o interior e resíduos para o exterior com uma urgência silenciosa. À esquerda, uma frústula de diatomácea — as suas paredes de sílica gravadas com matrizes de poros à escala nanométrica que difratam a luz azul-esverdeada em breves fragmentos prismáticos — está aprisionada por múltiplos filamentos convergentes e é arrastada impercetivelmente em direção à abertura, como presa numa teia de vidro vivo. Este organismo unicelular, sem cérebro nem sistema nervoso, constrói arquitectura mineral de precisão geométrica, estende uma rede de caça de vários comprimentos corporais e coordena o transporte intracelular ao longo de dezenas de pseudópodos em simultâneo — tudo através da física e da química de uma única célula eucariota.

Suspenso na água turva, olhas para cima e o que domina o teu horizonte inteiro é o ventre âmbar de um *Arcella vulgaris* — uma abóbada de quitina semi-transparente que se arqueia sobre ti como o teto de uma catedral em resina lacada, banhada por uma luz quente de mel dourado onde a iluminação transmitida atravessa as suas paredes mais finas e adenando-se em castanho-âmbar profundo ao longo do espesso rebordo equatorial. No centro preciso dessa cúpula, a abertura circular desce em direção a ti como a boca de um túnel em Portal escurecido, o seu lábio quitinoso lançando um halo de sombra fina contra o ouro envolvente, e através dele quatro lobopódios hialinos estendem-se para baixo como dedos de vidro — cilindros espessos de ectoplasma completamente transparentes, a progredir de forma glacial enquanto grânulos minúsculos derivam no seu interior como partículas suspensas em âmbar. A água que te rodeia não está vazia: fragmentos dourado-acastanhados de matéria vegetal decomposta pairam a meia profundidade, bactérias em bastonete derivam em grupos demasiado pequenos para serem individualmente resolvidos, e a iluminação geral — difusa, filtrada por camadas sobrepostas de matéria orgânica — confere à cena uma atmosfera de lanterna quente, enquanto a cúpula se curva para a periferia da tua visão, dissolvendo-se na escuridão aquosa como a borda de um mundo visto por baixo.

Você está pairando a milímetros do equador de um *Paramecium bursaria*, olhando através da sua película âmbar-dourada como se fosse vidro de catedral — e o que se revela do outro lado é um mundo inteiramente verde. Centenas de endossimbiontes *Chlorella* preenchem o citoplasma em mosaico contínuo, cada disco de clorofila com cerca de quatro micrómetros de diâmetro, tão próximos uns dos outros que o interior da célula imita uma copa florestal vista de baixo, camada após camada de pigmento afundando em sombra vegetal. A iluminação de contraste de interferência diferencial esculpe cada *Chlorella* em volume discreto — uma meia-lua iluminada, uma meia-lua em sombra — enquanto entre elas os canais citoplasmáticos brilham em âmbar citrino, pontuados por vacúolos alimentares e grânulos lipídicos que derivam como esferas castanhas na sol citoplasmático. No centro dessa floresta comprimida abre-se uma clareira pálida em lavanda-cinzento: o macronúcleo, corpo reniforme de textura cromatínica suave, cercado de verde como um prado alpino envolto em bosque denso. Na margem exterior da célula, os cílios projetam-se no meio aquoso como uma franja de prata viva, coletivamente luminosa, lembrando luz a partir-se em espuma de onda — o sinal exterior de uma arquitetura biológica que, aqui dentro, é absolutamente exuberante.
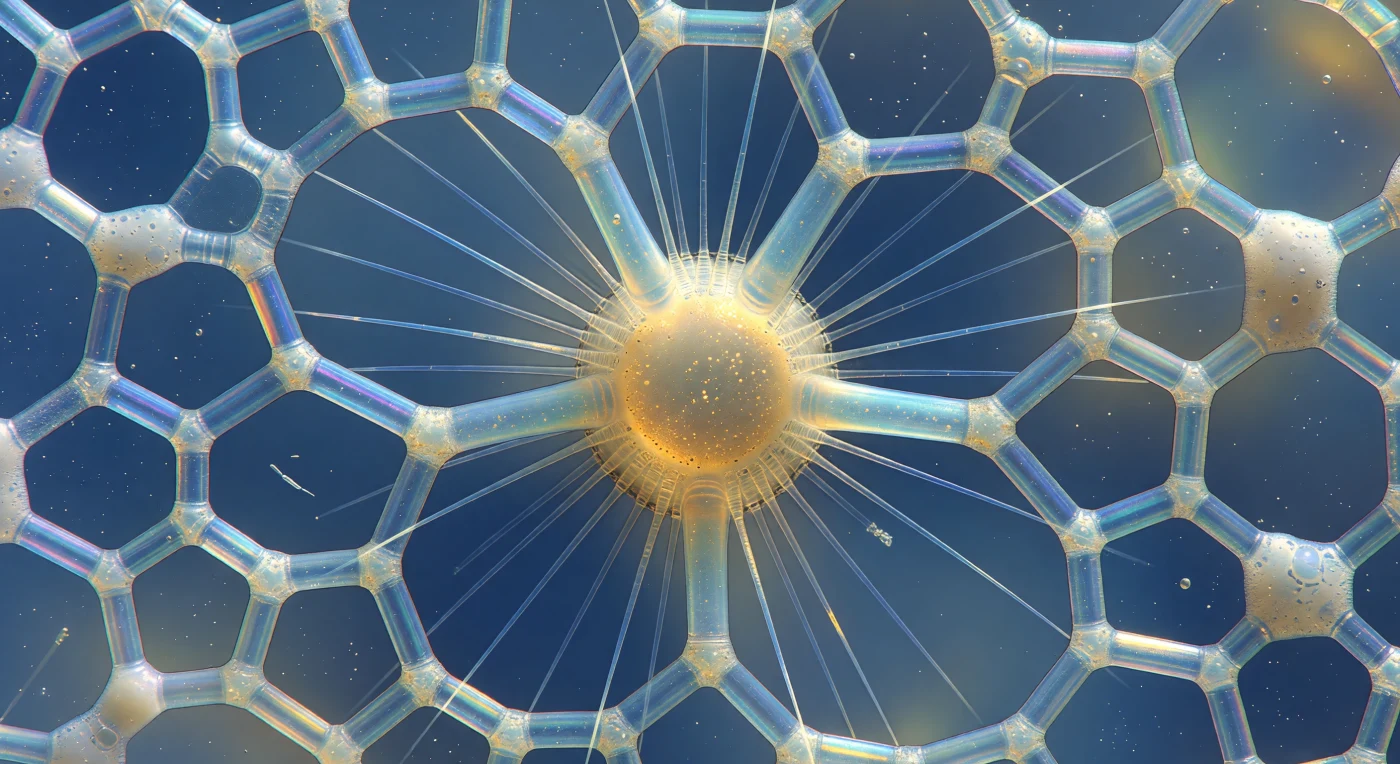
Suspenso a pouco mais de uma dezena de micrômetros do equador de um radiolário polycistino, o campo visual inteiro é consumido por uma arquitetura de sílica amorfa — feixe após feixe de ópalo vítreo, cada um com espessura de um fio de vidro soprado, curvando-se e perfurando o espaço à volta como a clerestória de uma catedral afogada no oceano aberto. A luz azul-esverdeada que desce da superfície fragmenta-se ao atravessar a malha em franjas de interferência que variam entre o violeta intenso nos ângulos agudos e o âmbar dourado nos nós mais espessos, enquanto cada poro hexagonal e pentagonal se abre como uma janela que enquadra o azul cerúleo infinito lá fora, transformando o esqueleto inteiro num relicário iluminado contra um crepúsculo perpétuo. No núcleo, a cápsula central pulsa com uma luminosidade âmbar e resinosa, sua membrana proteica perfurada mal visível como um anel escuro que delimita o sanctum interior onde cristais de sulfato de estrôncio — o lastro mineral do organismo — cintilam como fagulhas de ouro dentro do citoplasma em lenta convulsão. Dos axópodos irradiando para fora, filamentos de vidro perfeitamente retos refractam a luz ambiente em finas lascas prismáticas, enquanto entre as espículas externas uma camada de ectoplasma vacuolado se agarra em grumos de espuma translúcida, cada vacúolo uma lente minúscula que recolhe e distorce as cores de interferência em pontos luminosos fugitivos — e uma única haste bacteriana, imensamente menor, tomba em espiral através do hexágono mais próximo, revelando, sem qualquer aviso, a verdadeira dimensão desta catedral mineral que a evolução esculpiu em silício e silêncio.

Ao avançarmos rente ao fundo sedimentar, somos recebidos por uma vasta paisagem de ruínas geométricas em vidro — fragmentos de frústulas de diatomáceas amontoados em camadas densas, cada um esculpido em sílica opalina com a precisão de cristal lapidado, os seus arrays de poros nanométricos difratando a luz transmitida em franjas de ouro pálido, cobalto e rosa que se deslocam a cada fração de movimento do nosso ponto de vista. Cilindros de *Cyclotella* tombados evocam colunas de mármore desmoronadas, enquanto frústulas pennadas repousam inclinadas como quilhas de navios naufragados, as suas estrias a lançar rendilhados de sombra sobre o silte de válvulas partidas que cintila entre elas. Toda esta arquitectura mineral é o legado silencioso de organismos unicelulares fotossintéticos que, ao morrer, afundam as suas carcaças de sílica amorfa até formarem camadas de sedimento — a diatomite — que pode atingir espessuras de dezenas de metros ao longo de milhões de anos. No limite das ruínas próximas, flagelados cor de âmbar derivam nas colunas de água abertas entre as torres de frústulas, e uma *Actinophrys* heliozoária paira como um lustre de vidro suspenso, os seus axópodos rígidos — sustentados por feixes de microtúbulos axonemais — irradiando em todas as direções e capturando filamentos de luz azul transmitida como fibras óticas vivas, organismo e ruína habitando em simultâneo a mesma escala vertiginosa de quietude.
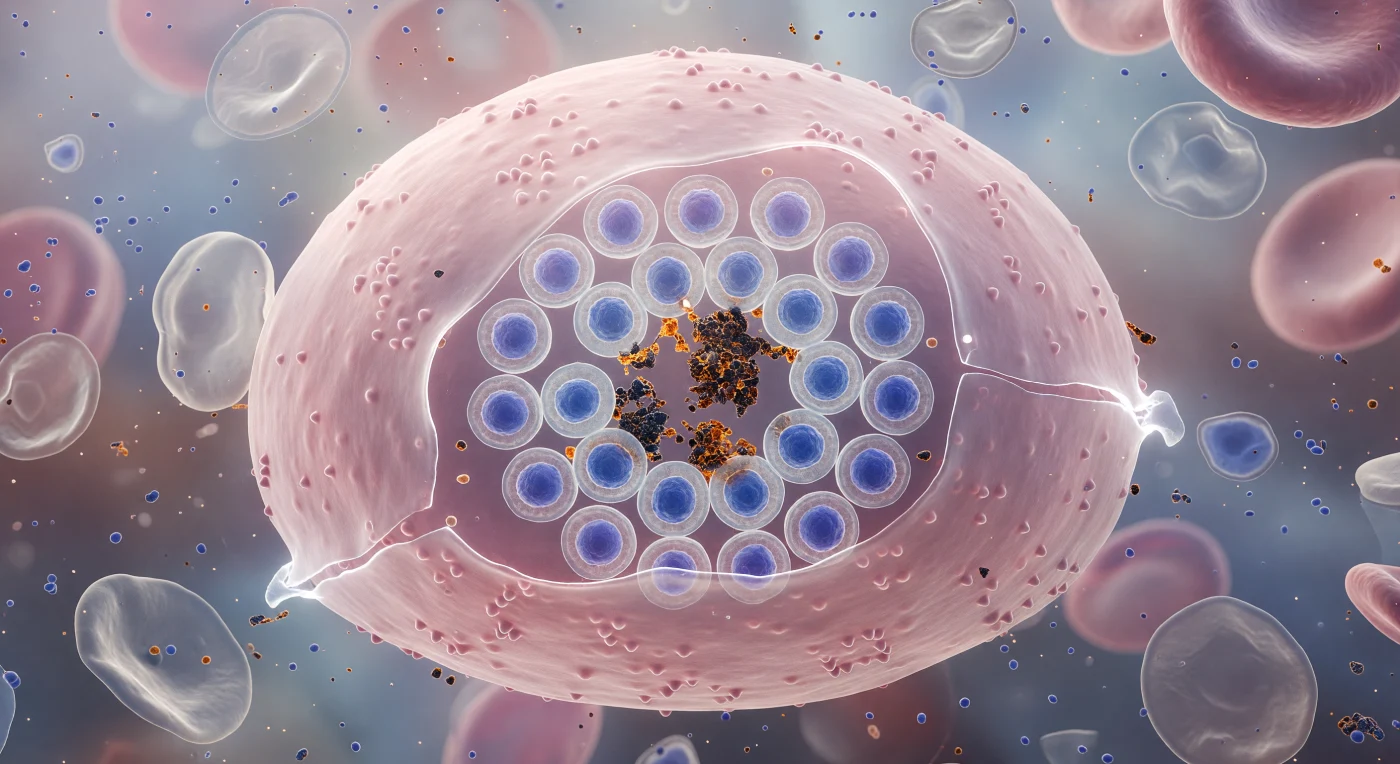
Você paira a uma distância quase irreal da superfície de uma célula vermelha do sangue infectada, cuja membrana esticada forma uma cúpula translúcida de cor rosa-salmão que se curva em todas as direções como o casco de um dirigível prestes a explodir. Através dela, pressiona-se de dentro uma roseta de vinte e quatro merozoítos de *Plasmodium falciparum*, cada um com o núcleo corado em azul-cobalto profundo, dispostos em roda com uma precisão geométrica quase vegetal, ao redor de grânulos de hemozoína — pigmento malárico cristalizado, denso e âmbar-negro, resíduo inerte da digestão da hemoglobina pelo parasita. A membrana, consumida até à quase-transparência pelo crescimento interno, treme sob a pressão osmótica acumulada, os seus pontos mais finos brilhando quase brancos como vidro iluminado por detrás, enquanto células fantasma vizinhas — envelopes cinzento-pálido de eritrócitos já rompidos — derivam enrugadas no meio levemente turvo de detritos proteicos libertados por ruturas anteriores. A tensão cromática entre o rosa quente da membrana e o azul frio dos núcleos parasitários que nela premem é simultaneamente bela e violenta: toda a cena suspensa no último instante antes da rutura liberar para o sangue um novo exército de invasores prontos a infetar células vizinhas.

A superfície que se estende em todas as direções é construída em celulose viva — as placas thecais de um *Ceratium tripos*, cada painel poligonal brilhando em âmbar-dourado profundo enquanto a luz azul-esverdeada do microscópio atravessa o interior fotossintético da célula e se refrata pelo material das placas, transformando cada território em mosaico de mel e siena queimada. As suturas entre as placas erguem-se como escarpas tectónicas, linhas geométricas precisas que dividem a superfície em domínios pentagonais e hexagonais texturizados por microcristas paralelas e poros circulares — arquitectura orgânica construída com a regularidade de uma obra de engenharia molecular, camada por camada de polissacarídeos organizados em lâminas. A cortar o terreno de lado a lado, o cíngulo equatorial abre-se como um canhão de índigo sombreado, e no fundo dessa fenda o flagelo transversal repousa enrolado, captando a luz difusa como um fio de iridescência pálida contra o escuro. Ao fundo, o corno apical sobe em espiral majestosa e desaparece para além do enquadramento, projetando a sua sombra âmbar sobre as placas vizinhas num lembrete de que esta paisagem continental não é rocha nem mineral, mas um único organismo unicelular — planctónico, deriva na coluna de água, e metabolicamente iluminado por dentro.
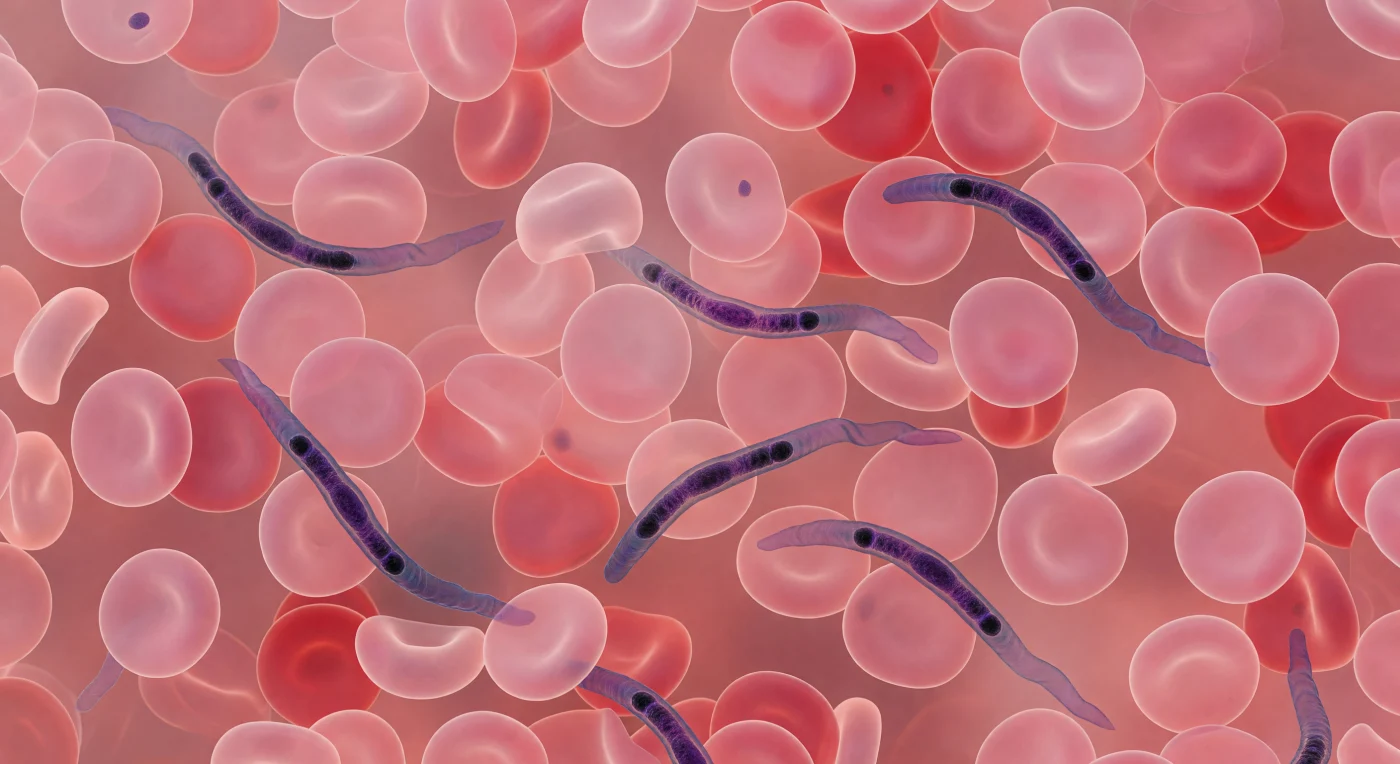
Dentro da corrente sanguínea, o olhar mergulha num labirinto sufocante de discos bicôncavos rosa-pálido que se estendem em todas as direções, cada glóbulo vermelho uma almofada translúcida de sete micrómetros cujo centro afinado deixa passar um halo âmbar de luz, enquanto os bordos engrossam num toro de coral-carmim mais denso — juntos formam um pavimento vivo e infinito que aperta de cima, de baixo e de todos os lados. Entre eles serpenteiam vários *Trypanosoma brucei*, corpos alongados de quinze a trinta micrómetros que se inscrevem no espaço intersticial como caligrafia azul-violeta, navegando cada corredor estreito entre os glóbulos com ondulações laterais lentas impulsionadas pela membrana flagelar — uma fita de seda rippling ao longo de um dos bordos que faísca em lavanda e índigo a cada propagação de onda. No interior de cada parasita, condensações cromáticas marcam o núcleo, de tom púrpura quase negro, e o cinetoplasto, um ponto ainda mais escuro encravado na extremidade posterior como uma semente compacta — estrutura singular da mitocôndria tripanosomática onde reside uma rede concatenada de ADN circular que coordena o metabolismo energético do parasita. A paleta Giemsa satura a cena inteira: aço-violeta contra rosa-carne, a única dinâmica neste mundo de pressão suave e lotação imóvel são as membranas flagelares dos tripanosomas, ondulando em arcos graciosos enquanto o plasma envolve tudo num âmbar translúcido e viscoso.

Estás suspenso a duzentos micrómetros abaixo da superfície de uma lagoa de água doce, e o teto do mundo é um espelho mercúrio ondulante — a face inferior do filme superficial — que devolve a luz da tarde em colunas âmbar-douradas que se refractam em fitas cáusticas deslizando lentamente sobre a comunidade como holofotes submersivos. Directamente diante de ti, uma multidão de espindles esmeraldas de Euglena pulsa com verde botânico saturado, os seus películos proteicos criando uma iridescência metálica dourada e prateada conforme a luz oblíqua percorre as suas flancos estriados, enquanto entre eles os Chlamydomonas ardem como lanternas de jade com um ponto ocular vermelho-tijolo — grânulo de carotenóide — que constitui o ponto de cor mais intenso de todo o campo visual. Os Coleps cilíndricos rodopiam lentamente entre eles, o mosaico de placas de carbonato de cálcio fragmentando a luz em facetas cintilantes como porcelana partida em queda, enquanto Chilomonas semi-transparentes derivam como óvalos de vidro óptico e uma névoa bacteriana azul-prateada dissolve suavemente o contraste à distância como neblina rasa. No plano de fundo, um fragmento de filamento de Spirogyra atravessa a cena como uma viga de cristal, a sua hélice de cloroplasto ainda visível através da parede celulósica translúcida — e tudo vibra numa densidade cinética suspensa, cada superfície molhada e refractiva, cada organismo uma lente viva dobrando a luz âmbar da lagoa em algo irredutível e complexo.

O mundo inteiro é azul-esverdeado e translúcido, visto de dentro para fora através das placas de quitina âmbar da própria larva — uma janela de vitral cor de âmbar que filtra a luz fria e difusa da coluna de água pelágica, onde partículas de neve marinha derivam em arcos brownianos lentos e células bacterianas cintilam como pontos de luz branca suspensa. A larva neste instante é a presa: seis apêndices articulados trêmulos alcançam o vazio e encontram resistência invisível, filamentos reticulopodiais de *Globigerina bulloides* com apenas 0,2 a 0,5 µm de diâmetro, revelados unicamente pelo trânsito contínuo de vesículas âmbar e partículas mitocondriais deslizando ao longo de cada fio como contas num rosário em movimento — correntes de cargo celular que denunciam a existência de uma rede que a luz sozinha quase não consegue revelar. A testa calcítica de *Globigerina*, branca-giz com subtons creme e amarelo-pálido, paira no distante canto superior como uma fortaleza mineral esférica e câmarada, quatro a cinco vezes maior que a larva capturada, suas centenas de aberturas superficiais emitindo os fios que agora convergem de todas as direções num cone de forças suaves e inexoráveis. Não há violência nem movimento brusco — apenas a geometria mecânica e fria de uma armadilha já fechada, os filamentos contráteis enrijecendo imperceptivelmente enquanto puxam o corpo da nauplius, milímetro por micrómetro, em direção ao monólito de carbonato de cálcio que aguarda imóvel na névoa luminosa.

Ao olhar para cima a partir de vinte metros de profundidade, o mundo revela-se como uma catedral de luz cobalto saturada — a radiação solar filtrada por duas atmosferas de água do mar até restar apenas o comprimento de onda de 460 nanómetros, que desce em colunas volumétricas e pulsantes que seguem o ritmo suave da ondulação à superfície. Nesta coluna de água viva, os Acantharia flutuam como estrelas caídas em plena explosão, cada um construído em torno de vinte espículas cristalinas de sulfato de estrôncio dispostas em simetria geométrica rigorosa segundo a lei de Müller, cujos eixos vítreos fragmentam a luz cobalto em coroas prismáticas de branco gelo, violeta pálido e ouro espectral que irrompem ao redor de cada organismo como halos frios. Entre estes organismos, lorica hialinas de tintinídeos — tubos cónicos de cocolitos aglutinados, âmbar na base e quase transparentes no bordo aberto — pendem em ângulos oblíquos na coluna, com os cílios compostos na abertura mal resolúveis como um frémito luminoso de quase invisibilidade. Flocos de neve marinha de mucus e detritos orgânicos derivam pelo enquadramento em múltiplas profundidades de campo, os seus contornos suaves e tons quentes de âmbar e creme contrastando com a precisão geométrica das espículas minerais, toda a cena suspensa numa coluna de luz viva sem chão nem tecto visíveis deste ponto de vista.

Num vasto deserto de argila rachada, a paisagem estende-se como um planeta deserto visto do rés do chão: planaltos poligonais de lama castanha-ocre separados por canhões de sombra tabaco, onde a luz dourada rasante ilumina cada aresta como uma crista incandescente. Espalhados por esta planície fraturada, os quistos dos protistas repousam como artefactos perfeitos — as esferas âmbar dos *Colpoda*, com as suas paredes duplas de quitina a capturar a luz oblíqua como gemas polidas, e as cúpulas dos *Arcella*, a textura geométrica das suas subunidades proteicas visível apenas a esta distância íntima, cada abertura selada com um tampo translúcido de material cístico pálido. Entre eles, os agregados de *Euglena* em palmela contraíram-se em lâminas irregulares de mucilagem ressequida com um brilho biológico subtil, as suas clorofilas adormecidas retendo um reflexo de verde-dourado apagado. Esta é a biologia da resistência absoluta: cada forma esférica encerra um organismo completo em diapausa metabólica, o genoma comprimido e protegido, aguardando que a água regresse e reative a maquinaria molecular suspensa há semanas, meses ou anos. A perfeição suave e simétrica de cada quisto contrasta deliberadamente com o caos fraturado do substrato mineral à sua volta — a ordem biológica forjada contra a desordem geológica.
