
Te encuentras suspendido en el borde de un cañón en espiral que desciende ante ti como un anfiteatro tallado en cristal vivo, las paredes del surco oral de un *Paramecium caudatum* arqueándose a ambos lados en la penumbra de contraste de fase, todo plata brillante contra un vacío carbón absoluto. Miles de organelos ciliares compuestos —membranelas y cirros dispuestos en filas densas sobre las paredes del cañón— baten en ondas metacronales sincronizadas que recorren la superficie como marejadas lentas sobre un mar luminoso, generando una corriente de vorticidad que arrastra bacterias bacilares doradas en espiral descendente hacia el pozo citostomal, una abertura pulsante de oscuridad total donde la membrana tiembla con el trabajo continuo de la fagocitosis. Este movimiento no es silencioso sino mecánico en su sentido más físico: a números de Reynolds de milésimas de unidad, el agua es un medio viscoso donde la inercia no existe y cada golpe de cilio impone su geometría de presión directamente sobre las partículas vecinas, convirtiendo el batido colectivo en una bomba biológica de precisión milimétrica. Por encima de ti, la pared celular translúcida se arquea como una cúpula de vidrio esmerilado, y a través de ella el macronúcleo —esa masa riñoniforme ámbar que contiene centenares de copias del genoma poliploides— presiona contra el ectoplasma con su luminosidad granular difusa, mientras en las profundidades del endoplasma las vacuolas alimentarias derivan como nubes de tormenta en distintas fases de digestión lisosomal, cada una un universo cerrado de química enzimática moviéndose lentamente en la corriente citoplasmática.

Desde donde te encuentras, suspendido en la profundidad de esta columna líquida, la vista hacia arriba es la de una catedral en movimiento: centenares de cuerpos fusiformes de *Euglena viridis* ascienden en espiral lenta a través del medio amarillo-verdoso, cada célula de sesenta micrómetros brillando con un verde eléctrico tan saturado que parece emanar luz propia, sus cintas de cloroplasto enrolladas helicoidalmente como vidrio de jade en suspensión. Cerca del extremo anterior de cada célula arde un punto minúsculo de color terracota encendido —el estigma, un orgánulo fotorreceptor primitivo compuesto por gránulos de carotenoides dispuestos en capas— que orienta la célula hacia el gradiente luminoso cálido que desciende desde la superficie del agua, difuso y dorado, visible en lo alto como un disco de ámbar rodeado de líneas cáusticas en lenta ondulación. La fototaxia positiva que organiza esta columna no es coordinada sino emergente: cada célula responde individualmente al mismo gradiente de irradiancia, y la suma de esas respuestas independientes genera la apariencia de una procesión coherente, un pilar de organismos que colectivamente transforman química y luz en orden. El medio mismo no es transparente sino un caldo de clorofila diluida, y en esa neblina lima flotan fragmentos de materia orgánica y bacterias en deriva browniana, motes de plata sin forma definida; debajo de ti, la luz mengua y el verde se vuelve oliva y luego oscuridad densa y quieta, donde la columna se disuelve y el agua recupera su peso.

Te encuentras aplastado contra un mosaico de sílice fracturada —los restos de frústulas de diatomeas antiguas, sus matrices de poros hexagonales atrapando la luz refractada como fragmentos dispersos de vidrieras— mientras sobre ti se alza, aplastante y arquitectónica, la estructura que domina este mundo: el seudópodo avanzando de *Amoeba proteus*, que se eleva como un glaciar tallado en vidrio vivo, su cúpula hialina delantera opticamente inmaculada doblando la luz ambiental en arcos dorados cálidos que se desplazan sobre el sustrato como anuncios luminosos de una invasión silenciosa. Por dentro de ese tubo ectoplásmico transparente, la corriente de endoplasma avanza en una avalancha densa y granular —un río de bronce líquido cargado de vacuolas alimenticias color miel ahumada, mitocondrias que destellan como esquirlas de cobre, gránulos refráctiles que ruedan en el flujo como piedras en una riada— todo ello gobernado por la transición sol-gel, ese límite vivo donde el citoplasma fluido se solidifica en tiempo real hasta convertirse en pared estructural, marcado por franjas de interferencia lavanda y oro pálido que ondulan como petróleo sobre el agua. A esta escala, donde la inercia no existe y el medio acuoso se comporta como un continuo viscoso y pegajoso, el simple acto de avanzar de una sola célula equivale al desplazamiento de un glaciar: lento, inexorable, transformando la geometría del mundo con cada micrómetro conquistado.
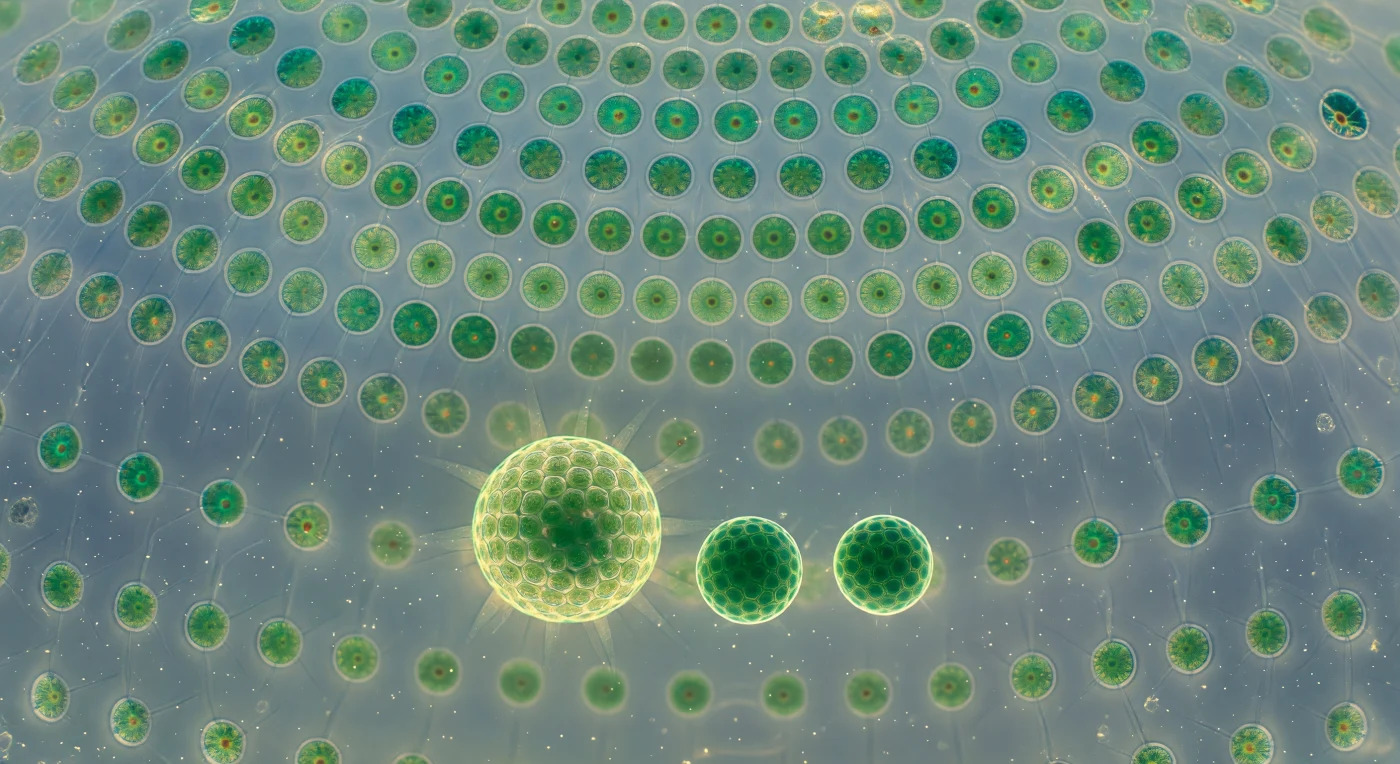
Desde el centro geométrico de la esfera, la vista se abre hacia una cúpula de vidrio vivo: miles de células somáticas biflageladas se distribuyen en una malla geodésica perfecta sobre la matriz de glicoproteína transparente, cada una un punto esmeralda de unos diez micrómetros engastado en la pared como una joya en ámbar pálido, todas conectadas entre sí por filamentos citoplasmáticos tan finos que solo se revelan como destellos plateados cuando la luz acuática los roza en el ángulo justo. El batido metacronal coordinado de sus flagelos recorre la superficie interior en ondas continuas de color verde esmeralda, una aurora lenta e incesante que mantiene a toda la colonia —un único organismo de miles de células— girando suavemente en la columna de agua. Flotando en el fluido acuoso del interior, tres colonias hijas de tamaños distintos se suspenden como planetas atrapados en una reliquia de cristal: la mayor ya luce su propia cúpula geodésica en pleno movimiento ciliar, la mediana resplandece con un jade más denso y saturado, y la más pequeña permanece quieta e inmóvil, una esfera uniforme de células aún sin flagelos activos. La luz difusa que penetra desde el exterior del estanque se refracta a través de la pared de glicoproteína y llena el interior con una luminiscencia azul-verdosa uniforme y envolvente, mientras que en cada célula de la cúpula arde un diminuto punto de óxido anaranjado —el ocelo, el organelo fotorreceptor— como una brasa encendida en medio del verde profundo. En este espacio que abarca apenas medio milímetro, la biología colonial alcanza una arquitectura que ninguna escala humana construyó jamás con semejante economía de materia.

La oscuridad que te rodea no es ausencia de luz: es un peso, una sustancia salada y orgánica que presiona desde todas las direcciones con la densidad silenciosa del océano profundo, y en ella flotas suspendido a la escala de una bacteria, invisible e inmóvil. Entonces la primera descarga detona a veinte micrómetros de tu posición: un estallido frío y cerúleo exactamente a 490 nanómetros, un azul-verde que no calienta ni hiere, y en su décima de segundo de existencia revela la silueta de una célula de *Noctiluca scintillans* —una esfera gelatinosa casi perfecta, su membrana plasmática captando su propia luz y doblándola hacia adentro en un halo brevísimo, su vacuola central ocupando la mayor parte del volumen como una linterna de aceite fantasmal mientras los orgánulos cargados de luciferina en el citoplasma periférico transitan del blanco-azul al verde profundo a medida que la reacción química agota su sustrato. La luz es confiscada antes de apagarse, y la oscuridad vuelve total e inmediata; pero ya una segunda descarga encadena a tres longitudes celulares de distancia, y luego otra detrás de ella, la perturbación mecánica del agua propagando la reacción en cascada de esfera en esfera, cada *Noctiluca* convirtiéndose en su propio testigo luminoso por una fracción de segundo antes de desaparecer, mientras copos de nieve marina —agregados de mucílago y fragmentos de frústulas de diatomeas— derivan entre los destellos y dispersan la luz cerúlea en halos neblinosos antes de que la infinidad negra los reclame de vuelta.

Suspensos justo en el borde del disco oral, miramos directamente hacia las profundidades de un *Stentor coeruleus* — una criatura unicelular que, a esta distancia, tiene la presencia de un abismo oceánico. Las membranelar que rodean la abertura forman una corona densa de paletas ciliadas translúcidas, cada una compuesta de cilios fusionados que baten al unísono en una barrida coordinada, generando una corriente giratoria que arrastra el agua circundante — y todo lo que flota en ella — hacia el pozo central de la citostoma: bacterias en bastón, fragmentos de algas verdes y detritos orgánicos espiralean hacia adentro como satélites cayendo en una singularidad gravitacional. El pigmento stentorina tiñe la pellícula en estriaciones paralelas de azul prusiano y aquamarina pálida, tan finas y regulares que la superficie del organismo parece una tela metálica tejida a escala molecular. A través de la pared semitransparente del cuerpo, el macronúcleo se revela como una cadena de perlas opalescentes — lóbulos discretos enhebrados verticalmente por el interior del citoplasma granular, cada uno refractando la luz cálida transmitida desde abajo como nódulos de cuarzo lechoso suspendidos en ámbar. Todo el organismo existe en un medio que no es vacío sino sustancia: el agua a esta escala es un continuo viscoso lleno de ruido browniano, y el movimiento de cada membranela representa una conquista cotidiana de la física de bajo número de Reynolds, donde la inercia no existe y solo la persistencia del latido ciliar sostiene el vórtice vivo.
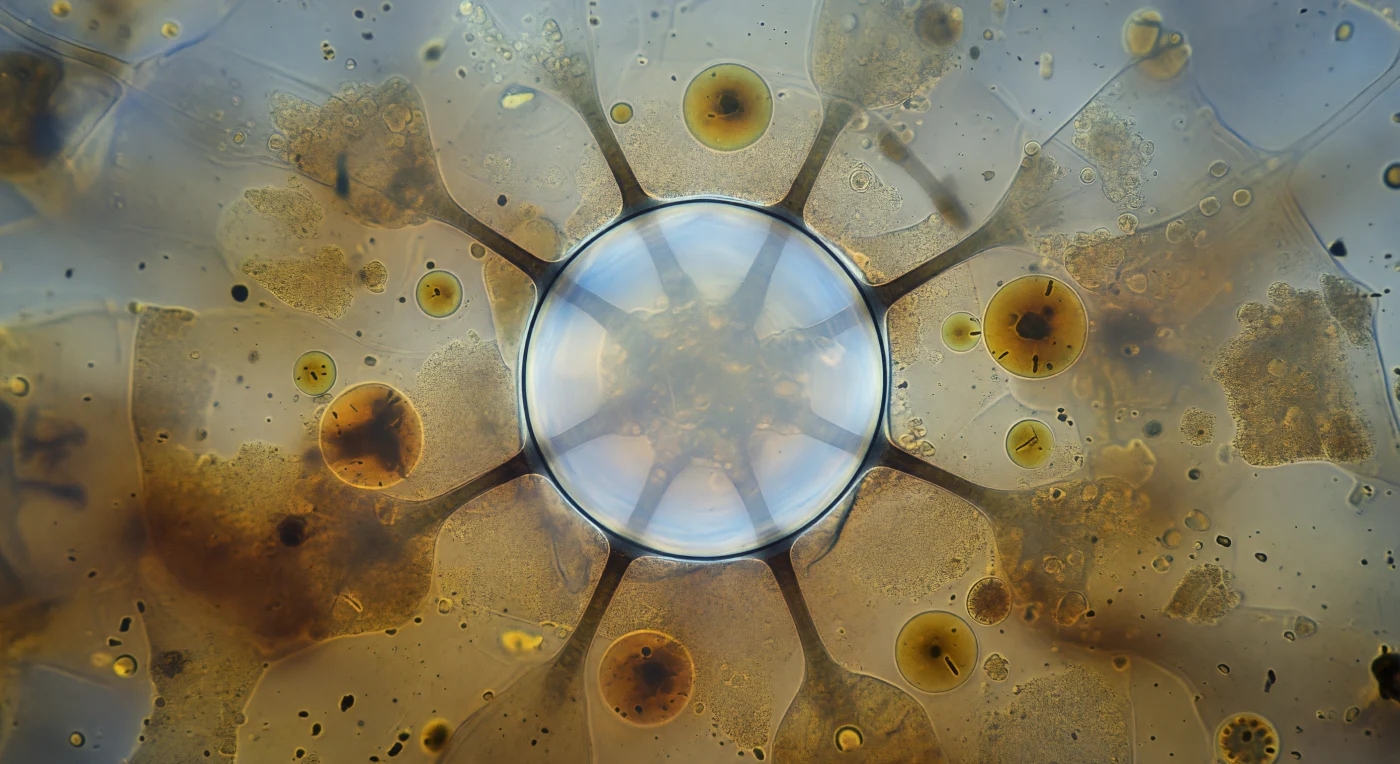
Dentro del cuerpo translúcido de un *Paramecium multimicronucleatum*, la vacuola contráctil ocupa el campo visual entero como una cúpula de vidrio vivo: una esfera de treinta micrómetros de diámetro, su membrana reducida a una línea de tensión plateada, su interior irradiando una luminosidad acuosa pálida que delata la carga osmótica acumulada durante decenas de segundos de bombeo incesante. Seis túbulos nefridiales se extienden desde su ecuador como radios oscuros que se disuelven en la penumbra granular del endoplasma, canales de membrana especializada que han estado drenando el citosol circundante gota a gota, forzando el exceso de agua hacia este depósito central antes de su expulsión al exterior a través del poro de descarga. La vacuola contráctil es la solución evolutiva de los protistas de agua dulce a un problema físico implacable: vivir en un medio hipotónico significa que el agua entra sin cesar por ósmosis a través de la membrana plasmática, y sin este sistema de bombeo activo —impulsado por ATPasas vacuolares que consumen energía metabólica en cada ciclo de cinco a sesenta segundos— la célula se hincharía hasta lisarse. A su alrededor, las vacuolas alimenticias de color ámbar derivan como planetas en órbita lenta, y el citoplasma granuloso presiona desde todas las direcciones con su constante corriente viscosa, mientras la esfera brillante aguanta, tensa hasta el límite absoluto de su geometría, un instante antes del colapso total que liberará toda su carga al mundo exterior.
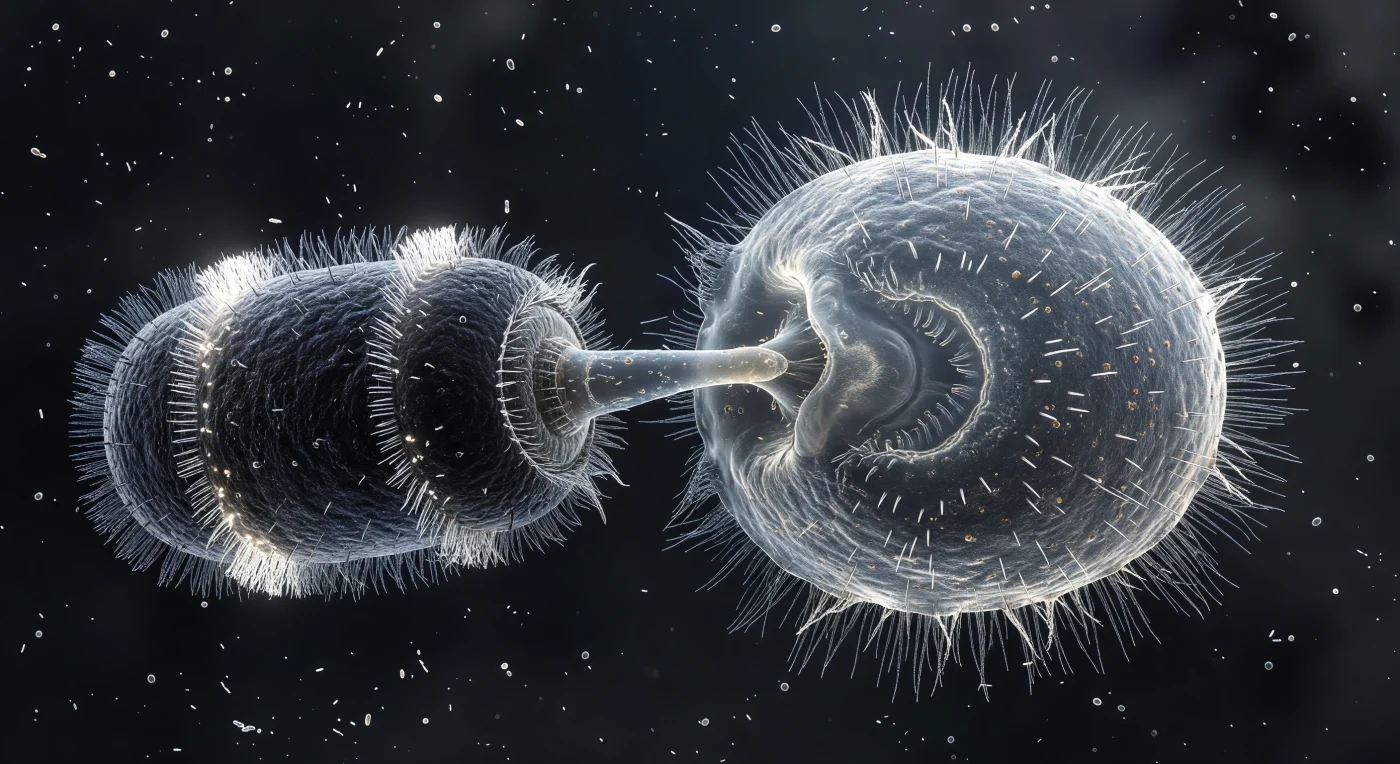
En el centro del encuadre, dos organismos se enfrentan en un instante de violencia biológica suspendida: a la izquierda, el *Didinium nasutum* aparece como un cilindro oscuro y compacto, cargado de potencia mecánica, sus dos coronas ciliares brillando como collares de escarcha blanca mientras su probóscide completamente extendida hunde su punta en el flanco del *Paramecium* y produce una concavidad brillante y tensa donde la pélícula cede bajo la presión del ataque. A la derecha, el *Paramecium* —una boya translúcida de citoplasma granular salpicado de vacúolas digestivas en distintas fases— responde con una descarga masiva y simultánea de tricocistos, proyectando hacia el medio una corona densa de filamentos blancos e increíblemente finos que se extienden varias longitudes corporales como una explosión de vidrio hilado congelada en su instante de máxima extensión. Los cilios del *Paramecium*, normalmente organizados en una onda metacrónica suave, están atrapados en un momento de inversión caótica, cada filamento girando en dirección propia, incapaces de sostener la coordinación habitual. El medio circundante, casi negro, está atravesado por una niebla bacteriana de bacilos y cocos a distintos planos de foco, creando una profundidad tridimensional que subraya la escala monumental de esta depredación microscópica.

Te encuentras a escasos micrómetros del fondo marino, suspendido en una luz azul-verde que parece emanar del propio agua como si el océano entero fuera un vitral difuso, y ante ti se alza la arquitectura calcárea de un foraminífero vivo — *Ammonia tepida* — cuyas cámaras globulares en espiral logarítmica brillan como piedra de luna tallada, cada pared de calcita finamente translúcida revelando apenas la sombra del citoplasma interior que las anima. Desde la apertura umbilical, oscura como la boca de una cueva ribeteada de materia orgánica ambarина, irradia en todas direcciones la red de reticulópodos: filamentos casi invisibles, hilos de vidrio húmedo tendidos sobre el sedimento que solo se delatan por el tráfico incesante de gránulos de organelas color ámbar y oro fluyendo en procesiones bidireccionales, algunos convergiendo hacia la concha, otros alejándose, de modo que la trampa entera se percibe viva a través del puro movimiento. A tu izquierda, una frústula de diatomea — sus paredes de sílice grabadas con matrices de poros a escala nanométrica que dispersan destellos prismáticos en la luz transmitida — está atrapada por varios filamentos convergentes y es arrastrada lentamente hacia la apertura, milímetro imperceptible tras milímetro, como presa en una telaraña tejida de vidrio vivo. Los granos de sedimento bajo tus pies son mundos en sí mismos: cuarzos del tamaño de peñascos, láminas de mica con superficies especulares que capturan destellos fríos y fugaces, pellets fecales colonizados por bacterias que se manifiestan solo como una neblina gris difusa, toda una geología íntima que recuerda que este organismo de cámaras nacaradas lleva construyendo trampas de reticulópodos sobre suelos como este desde hace quinientos millones de años.

Suspendido en el agua parda y turbia de un estanque, el espectador alza la mirada hacia una bóveda de dimensiones catedralicias: la teca de *Arcella vulgaris*, una cúpula de quitina semitransparente que ocupa el cielo entero como el techo ámbar de una capilla iluminada desde el interior, irradiando tonos miel dorado donde la luz transmitida atraviesa sus paredes más delgadas y oscureciéndose hasta el ámbar profundo en el grueso reborde ecuatorial. En el centro exacto de esa bóveda, un orificio perfectamente circular se abre hacia abajo como un portal oscuro rodeado de un labio quitinoso nítido, la apertura permanente por la que el organismo extiende sus lobópodos hacia el mundo exterior. Cuatro de esos pseudópodos descienden ahora hacia el espectador: cilindros de ectoplasma hialino, tan transparentes como el vidrio soplado, que captan la luz transmitida y brillan con un frío azul-blanco frente al cálido fondo ámbar, mientras minúsculos gránulos derivan en su interior como polvo en suspensión. Todo ocurre a escala de entre cien y doscientas micras, en un régimen donde la viscosidad del agua gobierna cada movimiento y la extensión glacialmente lenta de cada lobópodo representa, para el organismo, un esfuerzo titánico de reorganización citoplasmática.

En la penumbra verdosa que lo abarca todo, la pared de la célula se interpone como una ventana de ámbar ligeramente nervada, una membrana arquitectónica cuyas estrías longitudinales capturan la luz transmitida en finas líneas plateadas, recordando el nervado de una linterna gótica. Más allá, el interior del organismo aparece como un bosque denso e impenetrable: centenares de *Chlorella* endosimbiontes llenan el citoplasma en una mosaico casi continuo de discos verde hierba, cada uno apenas cuatro micrómetros de diámetro pero modelado por la iluminación de contraste diferencial en una pequeña moneda con volumen propio, su borde iluminado y su media luna en sombra. Donde los simbiontes se amontonan en el margen ectoplasmático su verde combinado se vuelve casi opaco, mientras que los canales de citosol líquido que los separan brillan como ámbar citrino translúcido, salpicados de gránulos lípidicos que derivan como ámbar oscuro entre los intersticios. En el corazón de esa selva interior, el macronúcleo emerge como un claro renal de lavanda pálida, suave y granular, una pradera alpina iluminada en medio del dosel, donde reside la maquinaria poliploidea que coordina la expresión génica de esta fábrica fotosintética viviente. En el borde exterior del organismo, los cilios proyectan al agua circundante un halo de filamentos plateados casi atmosférico, cada uno demasiado fino para resolverse individualmente, pero juntos formando una franja luminosa que estremece la frontera entre el interior habitado y el mar viscoso del mundo microscópico.
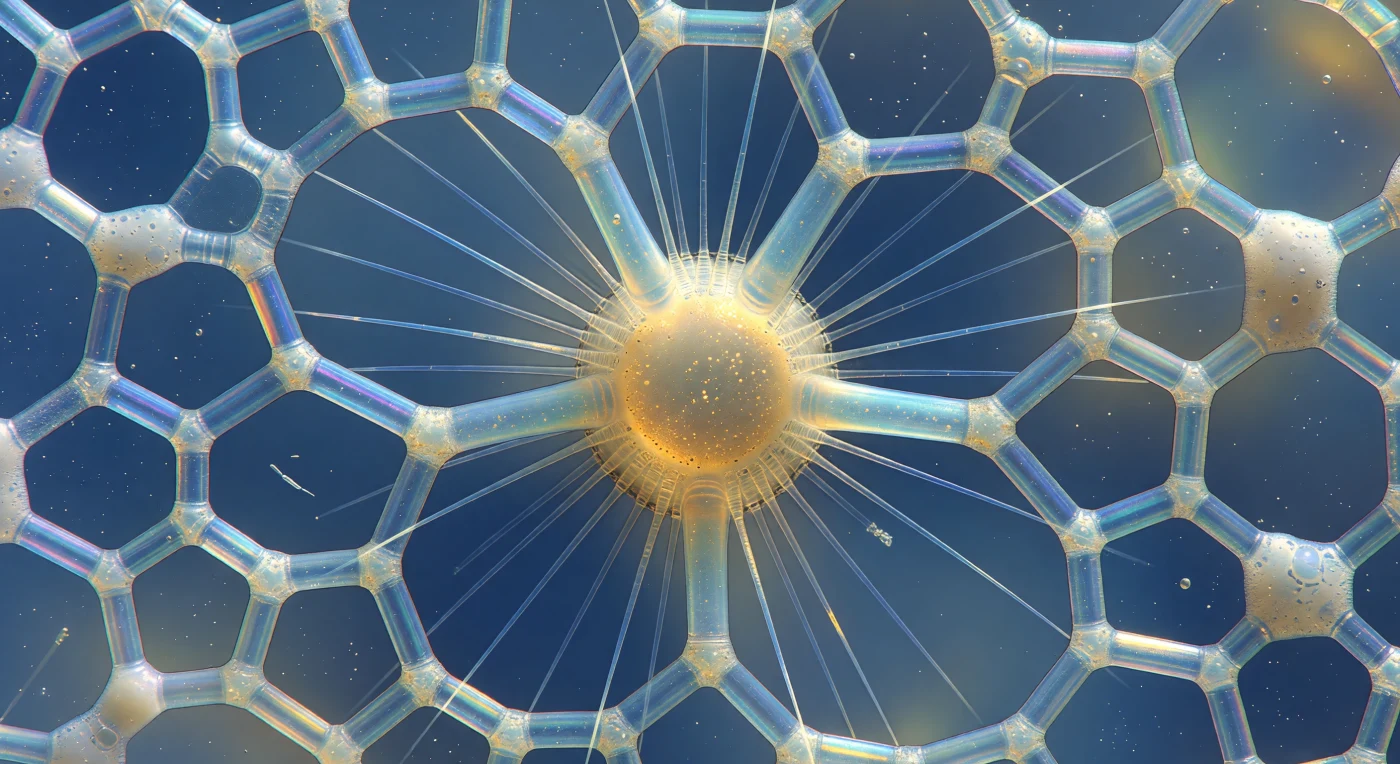
Suspendu à la hauteur équatoriale d'un radiolaire polycystine, votre regard traverse une architecture de silice opale amorphe dont les parois — aussi fines que du verre filé — fragmentent la lumière descendante en franges d'interférence qui glissent du violet aux bords aigus vers le cobalt le long des barreaux principaux, flambant en or pâle et ambre chaud là où la silice s'épaissit aux nœuds. Chaque pore hexagonal ou pentagonal ouvre comme une fenêtre de cathédrale sur l'eau océanique profonde, un halo de cerulean diffus, et là où deux couches du treillis se superposent en perspective, des auréoles de moiré naissent et se dissolvent au moindre frémissement thermique. Au cœur de l'organisme, la capsule centrale rayonne d'une luminosité ambrée et résineuse — membrane protéique perforée à peine visible en bordure sombre, cytoplasme intérieur chargé de gouttelettes lipidiques et de cristaux de sulfate de strontium scintillant comme des étincelles d'or — tandis que les axopodes s'élancent vers l'extérieur en filaments de verre parfaitement rectilignes, chacun une aiguille réfractive prismatique se perdant dans la brume cerulean ou blazant argent-blanc face à la lumière. Entre les spicules du squelette et l'éventail des axopodes, une écume de cytoplasme vacuolisé s'accroche en bulles translucides, chaque vacuole une minuscule lentille qui concentre momentanément les couleurs d'interférence en points lumineux fugaces, pendant qu'une bactérie en bâtonnet culbute librement dans la plus grande ouverture hexagonale — mesure involontaire et vertigineuse de l'échelle réelle de cette cathédrale minérale.

Nos deslizamos apenas por encima de un suelo de sílice cuya extensión hacia el horizonte difuso nos habla de una inmensidad construida enteramente a escala de lo invisible: cilindros de Cyclotella volcados como columnas de mármol en ruinas, frústulos pennados inclinados como quillas de barcos hundidos, triángulos de Triceratium y secciones de Coscinodiscus alzándose como escombros arquitectónicos de cristal tallado, cada uno entre veinte y ciento cincuenta micrómetros de anchura y sin embargo capaz de fraccionar la luz transmitida en franjas de interferencia que oscilan del cobalto al oro rosa con el más mínimo desplazamiento de nuestra perspectiva. La sílice no es del todo transparente ni del todo opaca —posee la translucidez lechosa del ópalo de gema, surcada de fracturas internas que abren arcos prismáticos en la luz filtrada desde arriba, como si un sol frío alumbrara a través de kilómetros de agua helada—, y entre los fragmentos mayores una polvareda de estría rota y válvulas sueltas forma un limo centelleante que rellena los intersticios con destellos espectrales en perpetuo movimiento. Al borde del campo de ruinas, flagelados de color ámbar derivan entre las torres de frústulos como pequeños elipsoides dorados suspendidos en la viscosidad del medio acuoso, donde la inercia no existe y cada desplazamiento es una negociación continua con fuerzas de superficie y gradientes químicos; más cerca, un heliozoo Actinophrys flota como una lámpara de fibra óptica viviente, sus axópodos —varillas rectilíneas endurecidas por haces de axonemas— irradiando en todas direcciones y capturando filamentos de luz azul transmitida entre las ruinas de cristal, un candelabro orgánico que preside desde la frontera entre lo inerte y lo vivo.
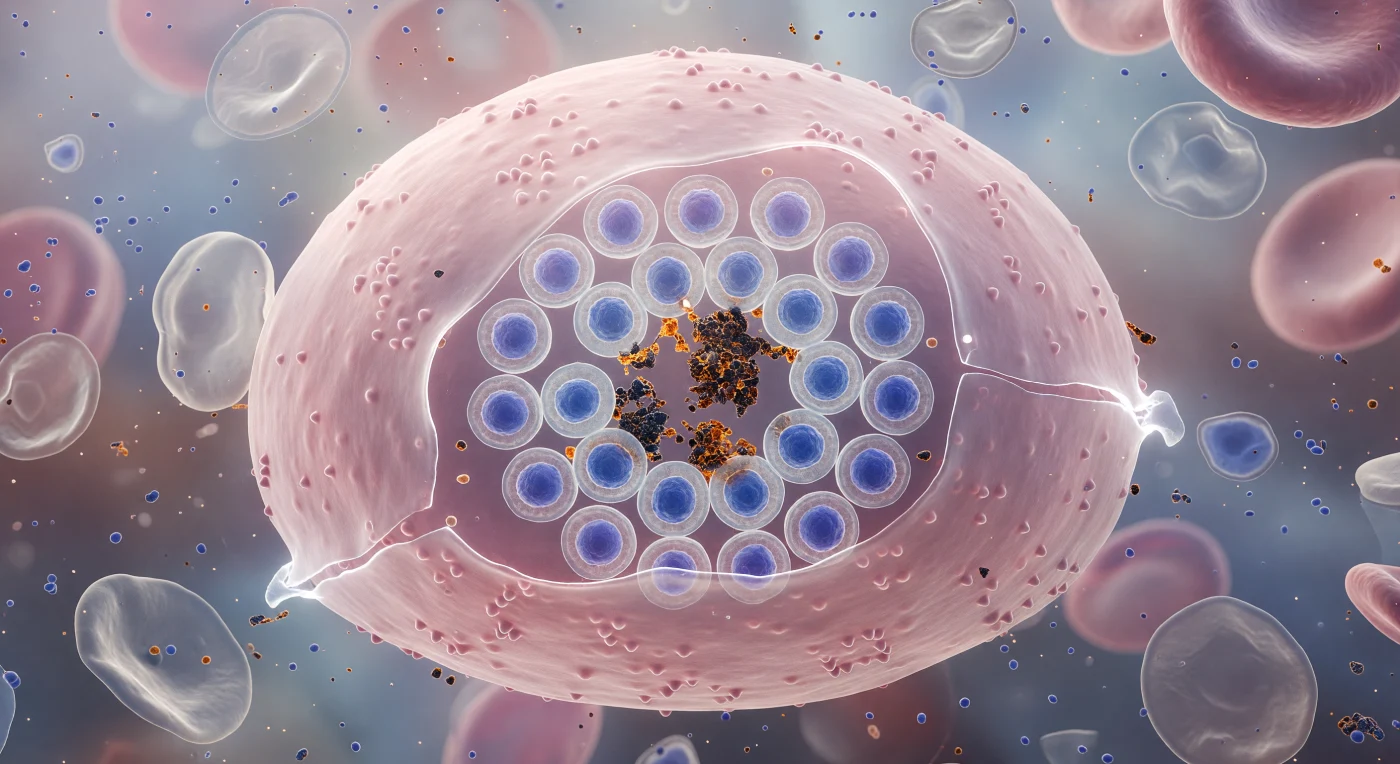
Suspendido a la escala de una bacteria, el observador se encuentra pegado contra la membrana exterior de un eritrocito infectado: una vasta cúpula rosada que se curva en todas direcciones como el casco de un dirigible translúcido y a punto de estallar, su bicapa lipídica adelgazada hasta la casi transparencia por la presión osmótica que se acumula en su interior. A través de esa pellícula fantasmal se distingue una roseta de veinticuatro merozoitos de *Plasmodium falciparum* dispuestos con precisión geométrica —cada núcleo teñido de azul cobalto profundo, rodeado de su propio halo citoplasmático pálido— formando una rueda perfecta en torno a un núcleo central de gránulos de hemozoína: pigmento de malaria cristalizado en ámbar oscuro y negro azabache, producto inerte de la digestión de hemoglobina que refracta la luz en destellos cálidos contra el frío violeta ambiental. La tensión cromática entre la membrana salmón y los núcleos parasitarios que la presionan desde dentro es casi violenta: las regiones más adelgazadas brillan de un blanco casi luminoso, mientras que las células fantasma vecinas —eritrocitos ya reventados, colapsados en pálidas envolturas grises y arrugadas— derivan más allá, recordando el destino inmediato de esta célula. Todo el sistema biológico está suspendido en la fracción de segundo anterior a la ruptura, cuando los veinticuatro merozoitos listos para la invasión se derramarán hacia el torrente sanguíneo circundante, perpetuando uno de los ciclos parasitarios más mortíferos de la historia de la vida.

Suspendido sobre el flanco de *Ceratium tripos*, el observador contempla una inmensidad de placas tecales de celulosa que se extienden en todas direcciones como las llanuras iluminadas de un continente extraterrestre: cada panel poligonal brilla con una calidez ámbar-dorada y siena quemado, calentado desde dentro por la luz transmitida azul-verdosa que se filtra a través del interior vivo de la célula. Las suturas entre placas se alzan como escarpes tectónicos con precisión geométrica, dividiendo la superficie en un mosaico de territorios pentagonales y hexagonales cuyas aristas proyectan sombras suaves sobre las estriaciones paralelas y los poros circulares perfectos que puntúan la armadura a intervalos regulares, cada uno un pozo oscuro que perfora las capas laminares de la teca. Atravesando el centro del encuadre de izquierda a derecha, el cíngulo ecuatorial abre un cañón de índigo sombreado en cuyo fondo apenas se distingue el flagelo transverso enrollado como una cinta dormida, captando tenuemente la luz dispersa como un hilo de iridiscencia pálida contra la oscuridad. A la izquierda, el cuerno apical asciende como una aguja monumental de ese mismo ámbar vivo, arqueándose con lenta majestuosidad antes de desvanecerse más allá del borde superior del encuadre, arrojando una sombra larga sobre las placas cercanas. Esta arquitectura orgánica —construida con la precisión de estratos geológicos— es en realidad la coraza viviente de un organismo unicelular marino, un dinoflagelado cuya teca celulósica no es hueso ni mineral, sino polímero biológico ensamblado célula a célula en el plancton oceánico.
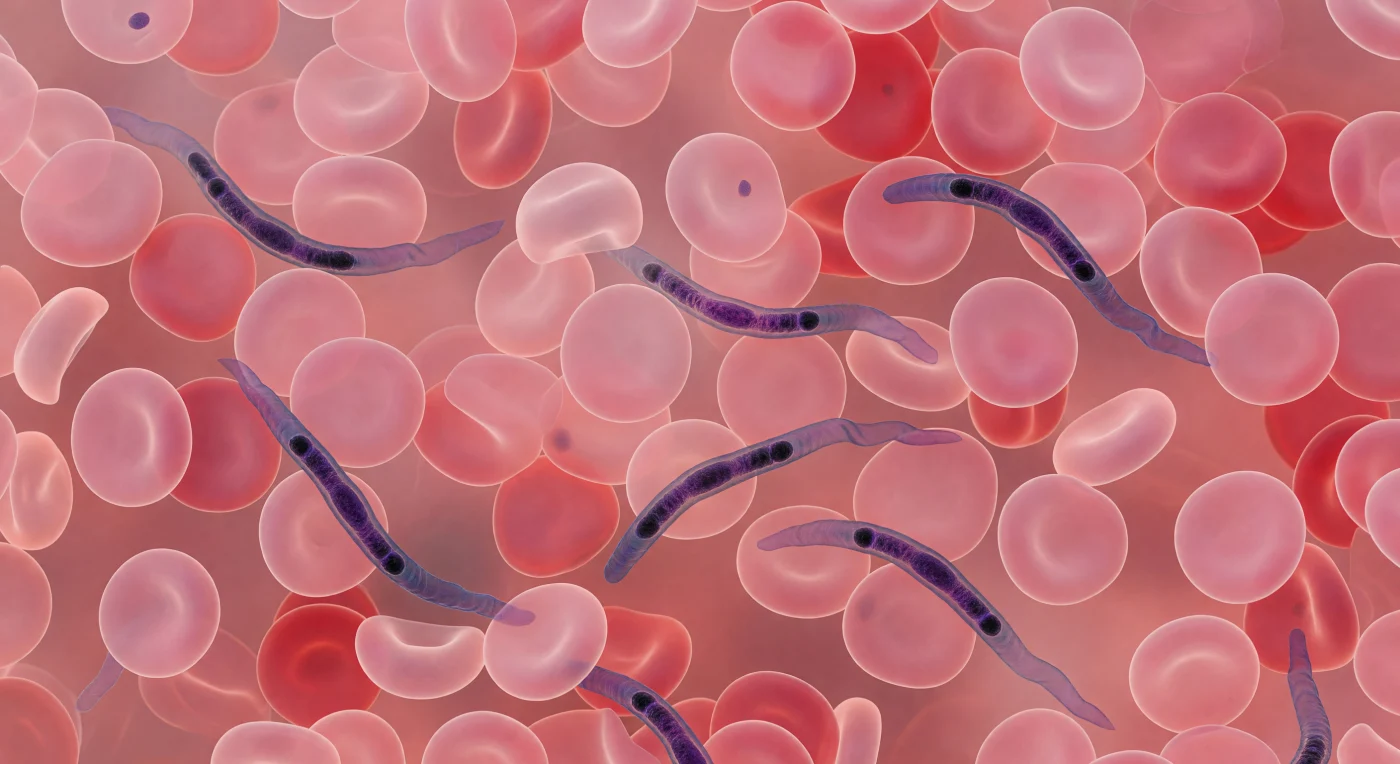
Entre los discos rosados que colman cada plano de visión hasta donde alcanza la mirada, varias formas violeta-azuladas se retuercen con lentitud calculada: son tripanosomas, parásitos unicelulares de quince a treinta micrómetros de longitud cuya membrana flagelar ondula como una cinta de seda empujando el cuerpo entre los glóbulos rojos con la eficiencia fría de organismos que han coevolucionado con la sangre de vertebrados durante decenas de millones de años. Cada célula roja que llena el campo es un disco bicóncavo de hemoglobina encerrado en una membrana lipídica de apenas siete nanómetros de espesor, siete micrómetros de diámetro, sin núcleo ni mitocondrias, reducida a su función esencial de transporte de oxígeno; amontonadas en todas las direcciones forman una geometría aplastante de carne rosada que deja solo angostos corredores donde los parásitos navegan. El kinetoplasto visible en cada tripanosoma como un punto casi negro cerca del extremo posterior es en realidad una red extraordinaria de ADN mitocondrial circular, único entre los eucariotas, que regula la bioquímica de un organismo capaz de alterar su cubierta de glicoproteínas para escapar sistemáticamente de la respuesta inmune del hospedador. La tinción de Giemsa congela esta negociación parasitaria en una paleta de violetas y rosas que, a esta proximidad, transforma el torrente circulatorio en algo que parece más paisaje que fluido: un mundo denso, claustrofóbico e infinitamente vivo.

La superficie del estanque no es un límite, sino un mundo: suspendido a doscientas micras por debajo de la interfaz aire-agua, el observador mira hacia arriba y encuentra un techo de mercurio ondulante que devuelve la luz ámbar de la tarde en destellos rotos, mientras columnas de oro líquido descienden en causticas movedizas que barren la comunidad como faros lentos. En primer plano, los fusiformes esmeraldas de las Euglena —cada una un torpedo de clorofila de quince a ochenta micras— llenan el campo como una multitud apretada de linternas vegetales, y entre ellas las Chlamydomonas brillan como piedras de jade con un punto de brasa roja incandescente, su estigma cargado de carotenoides, el color más saturado de toda la escena. Los Coleps ruedan despacio entre sus vecinos: cilindros blindados con mosaicos de placas de carbonato cálcico que fragmentan la luz en facetas parpadeantes, como porcelana rota que girara en el agua, proyectando pequeñas sombras duras cada vez que un rayo rasante los atraviesa. Más al fondo, las Chilomonas derivan como óvalos de vidrio óptico apenas opacificados por sus reservas de paramilo, mientras la neblina bacteriana —no células individuales sino un medio dispersante colectivo, una bruma plata-azulada— suaviza los contornos de todo lo que queda a más de unos cientos de micras, y un fragmento de filamento de Spirogyra atraviesa el plano de fondo como una viga de cristal, su hélice de cloroplasto todavía visible en verde jade a través de la pared celulósica translúcida. Todo vibra con la densidad cinética de un ecosistema completo comprimido en el grosor de unas pocas páginas apiladas, donde la tensión superficial, los gradientes químicos y los remolinos de las cilias vecinas son las únicas fuerzas que importan.

Desde dentro del caparazón del nauplius, el mundo exterior se revela como una trampa de geometría silenciosa: los filamentos reticulopodiales de *Globigerina bulloides* —casi invisibles, de apenas 0,2 a 0,5 µm de diámetro— se traicionan únicamente por el flujo continuo de vesículas ambarinas y partículas oscuras que avanzan en procesión ordenada a lo largo de cada hebra, como mercancía moviéndose por cables tensados. El caparazón de quitina color naranja-ámbar curva en los márgenes del campo visual, sus placas segmentadas y ligeramente iridiscentes donde los depósitos lipídicos atrapan la luz difusa del azul-verde marino, mientras los tres pares de apéndices larvarios se sacuden contra algo que la óptica apenas puede registrar. En la distancia superior derecha, el test calcáreo de la foraminífera domina el espacio como un monolito de tiza —blanco crema, multilocular, su superficie perforada por centenares de apertures diminutas desde las que emergen los mismos filamentos que ahora convergen sobre nosotros— y la escala relativa entre depredador y presa revela una asimetría arquitectónica brutal: cuatro o cinco veces nuestra propia dimensión, frío e indiferente. No hay violencia repentina: solo la retracción lenta y mecánica de una red que ya se ha cerrado, tirando del cuerpo larvario hacia el interior del colcoide marino cargado de partículas en suspensión, donde la nieve marina deriva en arcos brownianos y los objetos más distantes se disuelven en la neblina luminosa del pelágico abierto.

Suspendido en la columna de agua oceánica a veinte metros de profundidad, el mundo entero se convierte en una catedral de luz cobalto saturada, donde la radiación solar ha sido filtrada hasta que solo sobrevive la longitud de onda de 460 nanómetros, descendiendo en haces volumétricos suaves que pulsan con el ritmo lento de la marejada en la superficie. En este espacio flotan las Acantarias, organismos unicelulares cuyo esqueleto de veinte espículas de sulfato de estroncio —uno de los minerales biogénicos más raros del océano— irradia hacia afuera según una simetría geométrica precisa dictada por la ley de Müller, y esos cristales birrefringentes fragmentan la luz cobalto en coronas prismáticas de blanco glacial, violeta pálido y oro espectral que se expanden alrededor de cada célula como halos fríos. Entre estas estrellas minerales, las loricas de los tintínidos cuelgan como minúsculos jarrones de vidrio ámbar, construidas de cocolitos y partículas inorgánicas aglutinadas, y en sus bordes abiertos los cilios compuestos apenas se resuelven como un temblor luminoso de casi invisibilidad, recordatorio de que estas estructuras son trampas activas de fitoplancton impulsadas por ondas mecánicas que operan a frecuencias de veinte a cuarenta ciclos por segundo. Cruzando el encuadre a múltiples profundidades focales, los copos de nieve marina —agregados de mucus, detritos y colonias desintegradas— derivan con bordes suaves y tonos cálidos de ámbar y crema, ofreciendo un contraste de fragilidad orgánica contra la precisión mineral de las espículas cristalinas, todo suspendido en una columna de luz viva que no tiene suelo ni techo visible desde aquí.

Ante tus ojos se extiende una llanura inmensa de barro resquebrajado, un mosaico de placas poligonales de arcilla siena y ámbar oscuro separadas por grietas profundas que se hunden en sombra como cañones de un planeta desecado. Una luz dorada rasante barre este paisaje mineral desde un ángulo bajo, encendiendo cada arista de arcilla como una cresta luminosa mientras el interior de cada fisura se pierde en una oscuridad tabaco y húmeda. Dispersos sobre estas mesetas y acurrucados junto a los bordes de las grietas, los quistes de protistas ocupan el terreno como pequeñas joyas pulidas: las esferas de *Colpoda*, de unos 15 µm de diámetro, atrapan la luz oblicua y revelan su doble pared quitinosa como un halo translúcido, mientras los tecas de *Arcella* —cascos abovedados de proteína ensamblada en subunidades geométricas— sellan sus aperturas con un tapón pálido que los convierte en cápsulas perfectas de vida suspendida. Entre ellos, las colonias de *Euglena* en estado de palmela se extienden como láminas de mucílago reseco con un brillo biológico leve, sus bordes enroscados hacia arriba y teñidos de un verde oliva apagado donde los cloroplastos duermen con sus puertas metabólicas cerradas, esperando el regreso del agua para despertar.
