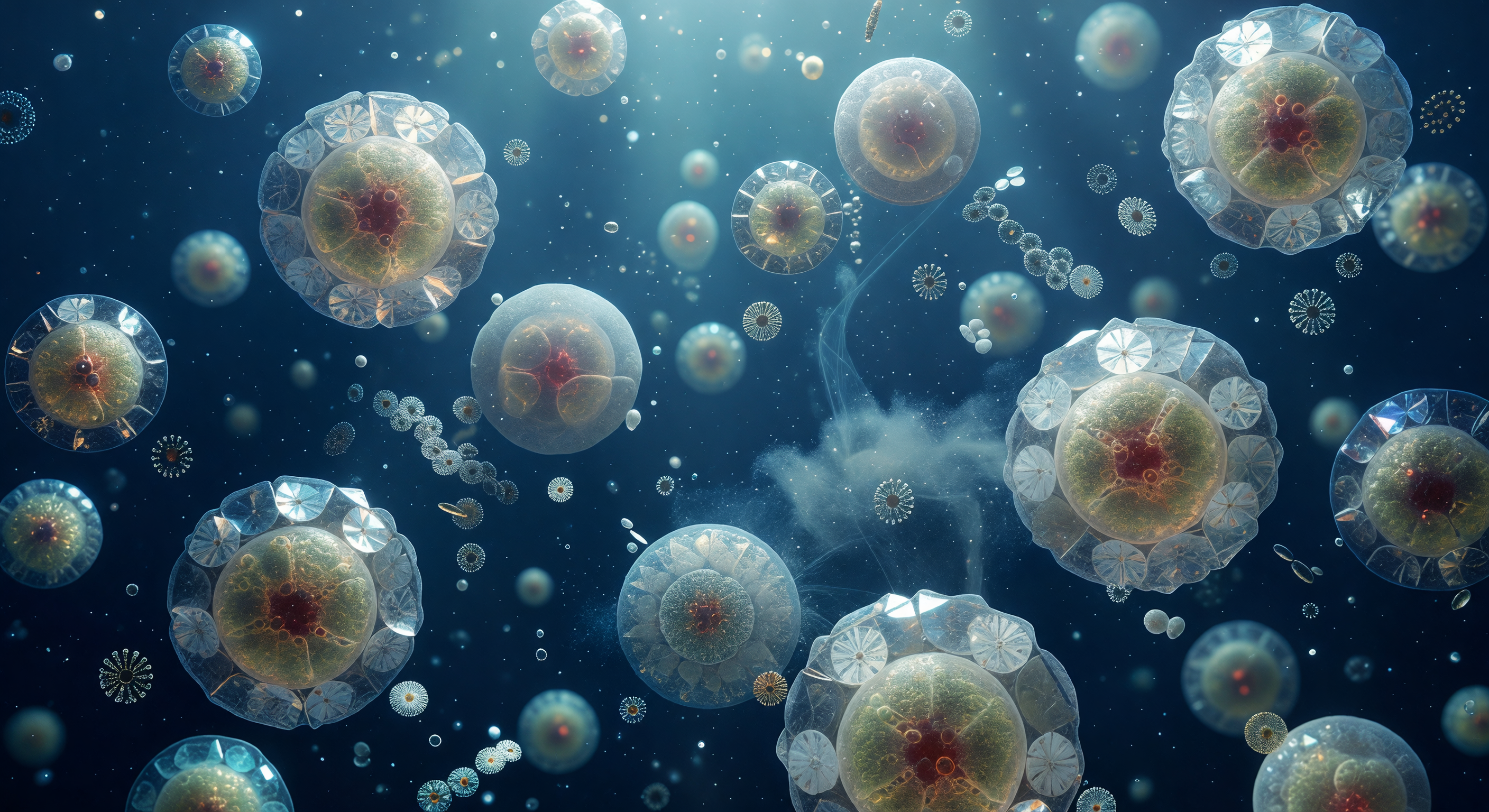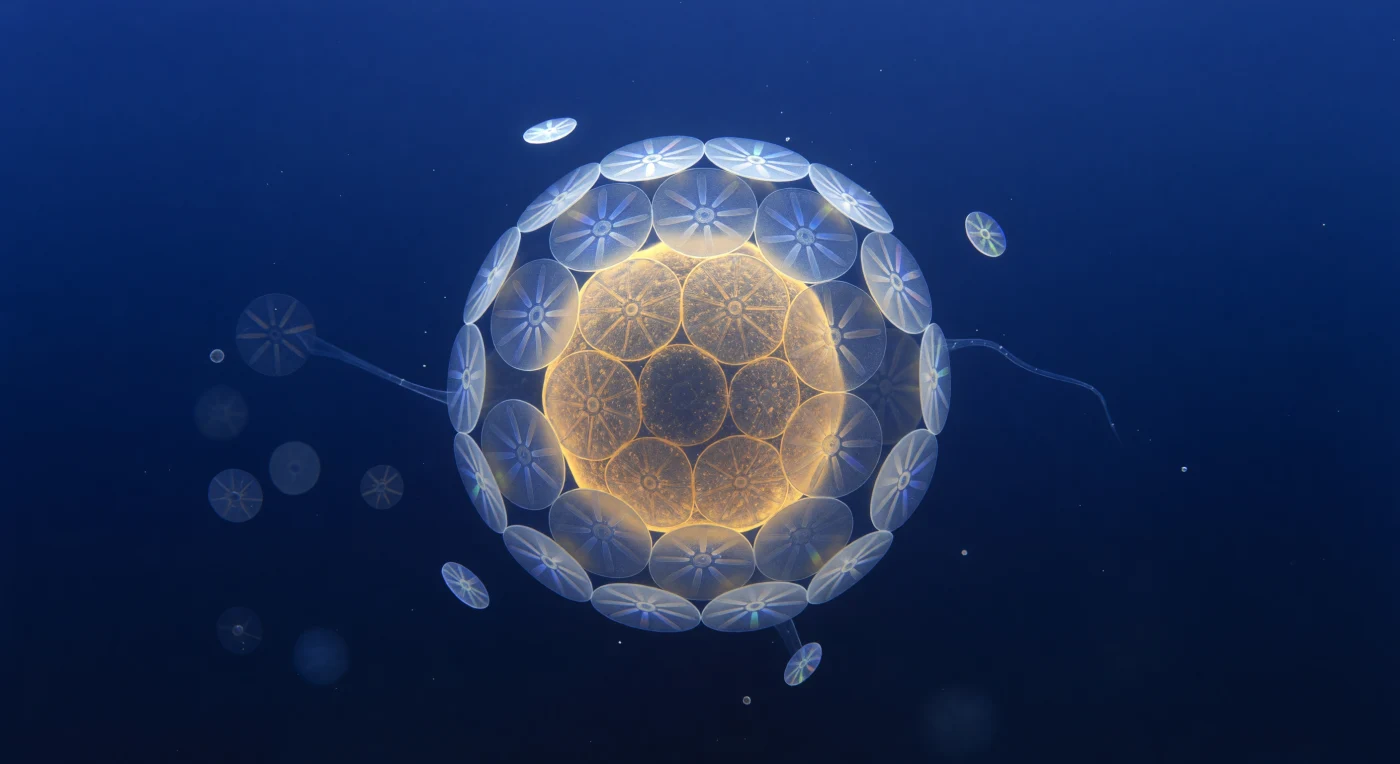
Ante ti se despliega una luna ámbar diminuta: la célula de *Emiliania huxleyi*, una esfera translúcida de ocho micrómetros cuyo interior encendido —cloroplastos apretados como ascuas de miel— derrama halos de azafrán y siena a través de la membrana plasmática. La coraza que la envuelve es una obra de ingeniería biomineral: veinte cocolitos de calcita entrecruzados con la regularidad de un relojero, cada radio cristalino convergiendo en un eje central que, al atrapar los fotones difusos de 480 nanómetros filtrados desde la superficie, fractura la luz en destellos efímeros de violeta helado, verde menta y blanco frío antes de ceder al penumbra índigo de sus vecinos. Más allá de la célula, tres o cuatro cocolitos desprendidos giran lentamente en el ruido térmico del agua, discos luminosos que lanzan prismas fugaces mientras el medio que los rodea —un ultramar sin equivalente terrestre, azul comprimido de fotones que ya han atravesado metros de columna de agua— se vuelve niebla índigo a unos cincuenta diámetros celulares de distancia. En los márgenes del campo visual asoman los contornos fantasmales de otras coccoesferas y un filamento de exopolímero transparente que atraviesa la escena como vidrio hilado, recordando que este mundo aparentemente vacío es en realidad una arquitectura densa e invisible, simultáneamente tan sólida como la piedra caliza y tan frágil como una pompa de jabón.
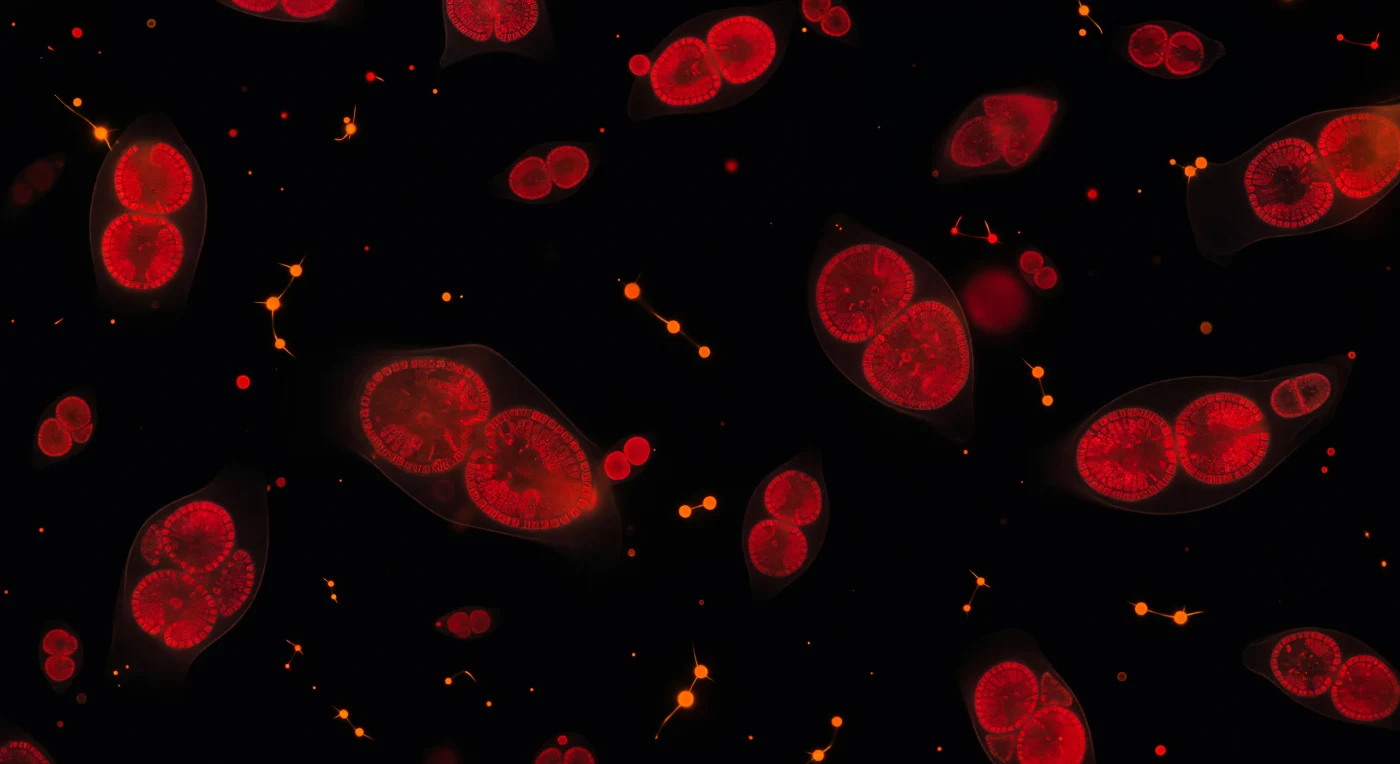
En la oscuridad absoluta de unos pocos micrómetros, decenas de células de fitoplancton arden como brasas de carmesí profundo, su autofluorescencia de clorofila emitiendo fotones a 680 nm desde el interior de sus propias membranas sin ninguna fuente de luz exterior. Cada célula revela cloroplastos en forma de riñón doble —estructuras bilateralmente simétricas densas en pigmento— cuyas membranas tilacoides apiladas en láminas atrapan esa luz roja autogenerada y la profundizan hacia el borgoña en los pliegues más gruesos. Dispersas entre las masas carmesí, las cianobacterias portadoras de ficoeritrina destellan como chispas anaranjadas de temperatura más alta: organismos procarióticos más pequeños y geométricamente simples que los eucariotas que los rodean, algunos alineados en pares o cadenas que trazan constelaciones mínimas en el medio invisible. El propio fluido se delata apenas en halos de materia orgánica disuelta y vesículas de membrana que dispersan el resplandor celular en débiles aureolas rojizas de un micrómetro de grosor, confirmando que habitamos una suspensión acuosa y no el vacío. Este campo de estrellas comprimido en una gota representa la base fotosintética de la cadena trófica marina: cada fotón capturado por estos pigmentos impulsa la fijación del carbono que sostiene, en última instancia, ecosistemas de escala oceánica.

El océano que te rodea ha dejado de ser agua: es una suspensión mineral viva, opaca como tiza diluida, donde diez millones de ruedas de calcita por mililitro dispersan la luz solar de vuelta hacia el cielo antes de que pueda penetrar más allá de la palma de tu mano. La superficie se extiende en todas direcciones como una plancha cegadora de jade blanquecino y turquesa cremoso, surcada por cintas paralelas de color leche condensada —las windrows de Langmuir— que revelan los vórtices contrarrotantes ocultos justo bajo la interfaz, concentrando las partículas más ligeras en líneas visibles desde el horizonte. Donde el bloom termina, la frontera corta el cielo con una precisión casi quirúrgica: a un lado, la pared de tiza turquesa que se extiende hasta el punto de fuga; al otro, el azul cobalto del océano abierto, oscuro y transparente como tinta por contraste, extraordinariamente nítido después de tanta opacidad. Mirando hacia abajo, tu propio cuerpo sumergido desaparece tras apenas cinco centímetros de claridad, engullido por una luminosidad blanco-verdosa que no proviene de ninguna dirección concreta sino de todas a la vez, fotones atrapados en un laberinto de geometría calcárea generada por organismos de cinco micrómetros que, en su conjunto, han alterado la reflectividad de miles de kilómetros cuadrados de Atlántico Norte. Un leve aroma a dimetilsulfuro —esa nota verde y marina que las células liberan al lisarse— impregna el aire sobre esta superficie que es, simultáneamente, organismos vivos, mineral precipitado y sistema meteorológico biogénico tendido sobre el mar.
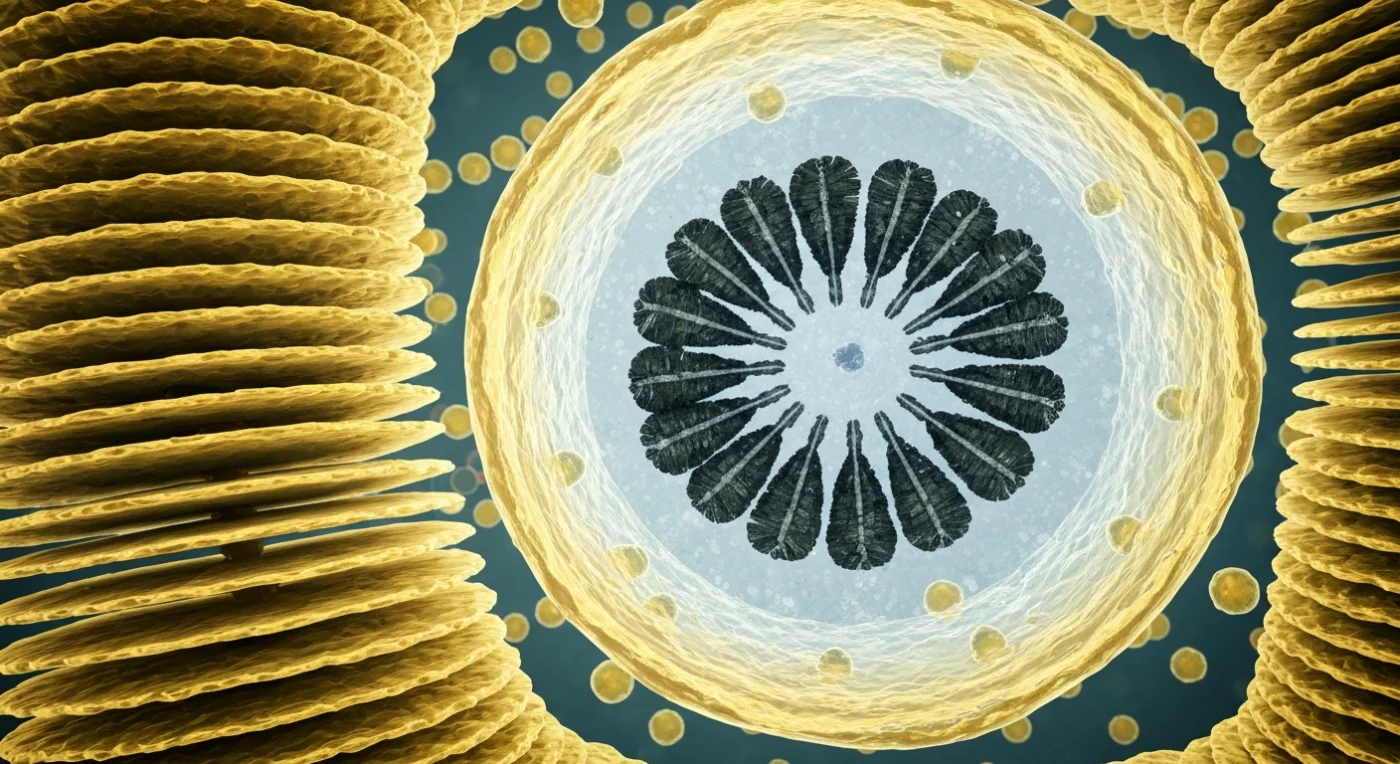
Ante ti se extiende el interior de una fábrica invisible: láminas de membrana dorada apiladas en paralelo con una precisión casi arquitectónica, los cisternas del aparato de Golgi, cada una de apenas unas decenas de nanómetros de grosor, translúcidas como vidrio de ámbar batido y ligeramente luminiscentes en su borde trans, donde se inflan en vesículas que se desprenden y derivan lentamente por el citoplasma como faroles cargados de precursores de calcita. Todo el espacio circundante está ocupado por una niebla densa de ribosomas que tapizan las superficies del retículo endoplasmático en constelaciones apretadas, dando a cada membrana una textura rugosa, casi aterciopelada, que recuerda al papel de grano fino visto bajo luz rasante. Dominando el campo visual como una cúpula industrial, la vesícula del cocólito se hincha con sus paredes de ámbar translúcido albergando en su interior acuoso y frío un disco basal de polisacárido —iridiscente y frágil como seda de araña— sobre el que se ensambla, en tiempo real, un anillo naciente de prismas de calcita de color carbón oscuro con facetas internas que devuelven destellos plateados oblicuos. Se trata del proceso por el cual Emiliania huxleyi fabrica sus placas minerales: iones de calcio que atraviesan la membrana vesicular en pulsos invisibles y cristalizan con una geometría tan precisa que parece ingeniería, todo ello en silencio absoluto dentro de un citoplasma tan abarrotado que cada nanómetro cúbico está ocupado, y donde la viscosidad del medio convierte cada movimiento en un gesto lento, deliberado e inexorable.

Ante ti flota una esfera de fuego contenido en la oscuridad absoluta: una cocosfera formada por quince ruedas de calcita perfectamente engranadas, cada una ardiendo con el patrón de extinción en cruz de Malta que la luz polarizada arranca de su estructura cristalina, negro aterciopelado contra blanco de primera orden tan denso que parece tener presión interna. En los bordes engrosados de cada escudo distal, donde la diferencia de camino óptico aumenta marginalmente, el blanco cede a un oro pálido y a un marfil cálido, como luz de vela sobre nieve, revelando la arquitectura de treinta prismas de calcita interbloqueados que componen cada cocolito individual. Alejándose de esta esfera central en todas las direcciones, cocolitos libres derivan en el campo visual como una correa de asteroides: algunos de frente, mostrando la simetría completa de su catedral cristalográfica; otros de canto, finas lunas crecientes luminosas girando imperceptiblemente en la deriva browniana. La profundidad del espacio negro se establece únicamente por la progresiva pérdida de nitidez de los cocolitos más distantes, reducidos a manchas luminosas suaves, sin ninguna otra referencia que la geometría de estos organismos que construyeron esta arquitectura de calcita ion por ion, en la oscuridad, en el frío, a miles de metros sobre el fondo oceánico.
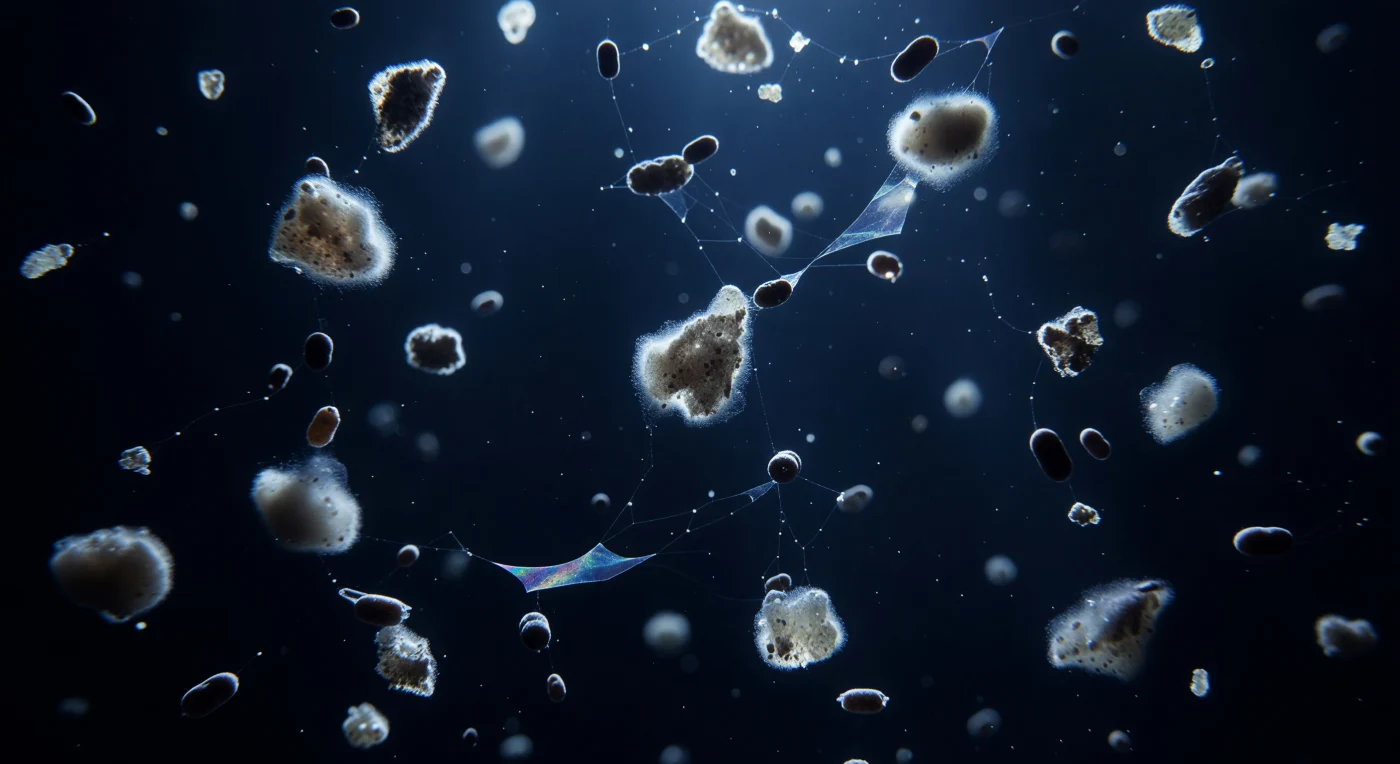
Suspendido a doscientos metros de profundidad, el observador flota en una columna de agua de un índigo casi absoluto, atravesada por una procesión silenciosa de copos de nieve marina que descienden desde arriba: masas irregulares de uno a cuatro milímetros, construidas con frústulas de diatomeas muertas, fragmentos de calcita cocolitífera, mucus polimérico y detritos celulares compactados durante días de agregación lenta. Los últimos fotones atenuados que sobreviven desde la zona eufótica impactan contra los cristales de calcita incrustados en cada flóculo y los encienden en un halo blanco-azulado tenue —luz dispersada entre 450 y 490 nanómetros— que hace que cada agregado lleve consigo su propia linterna fantasmal hacia el abismo, mientras su interior gris-marrón concentra la memoria química de lípidos, proteínas, polisacáridos y minerales fused en materia que cae. Entre los copos, los pellets fecales cilíndricos de color marrón oscuro descienden tres a diez veces más rápido, compactos y lisos, atravesando el campo de nieve como pequeños proyectiles, mientras filamentos de partículas de exopolímeros transparentes —TEP— conectan los agregados en una red iridiscente casi invisible, revelándose solo en sus bordes como membranas de jabón colapsadas. Todo el conjunto constituye la bomba biológica de carbono del océano en acción: cada flóculo que alcance el sedimento entierra carbono fijado por fotosíntesis, un archivo geoquímico que ha caído sin testigos durante cientos de millones de años y que hoy regula el clima del planeta.
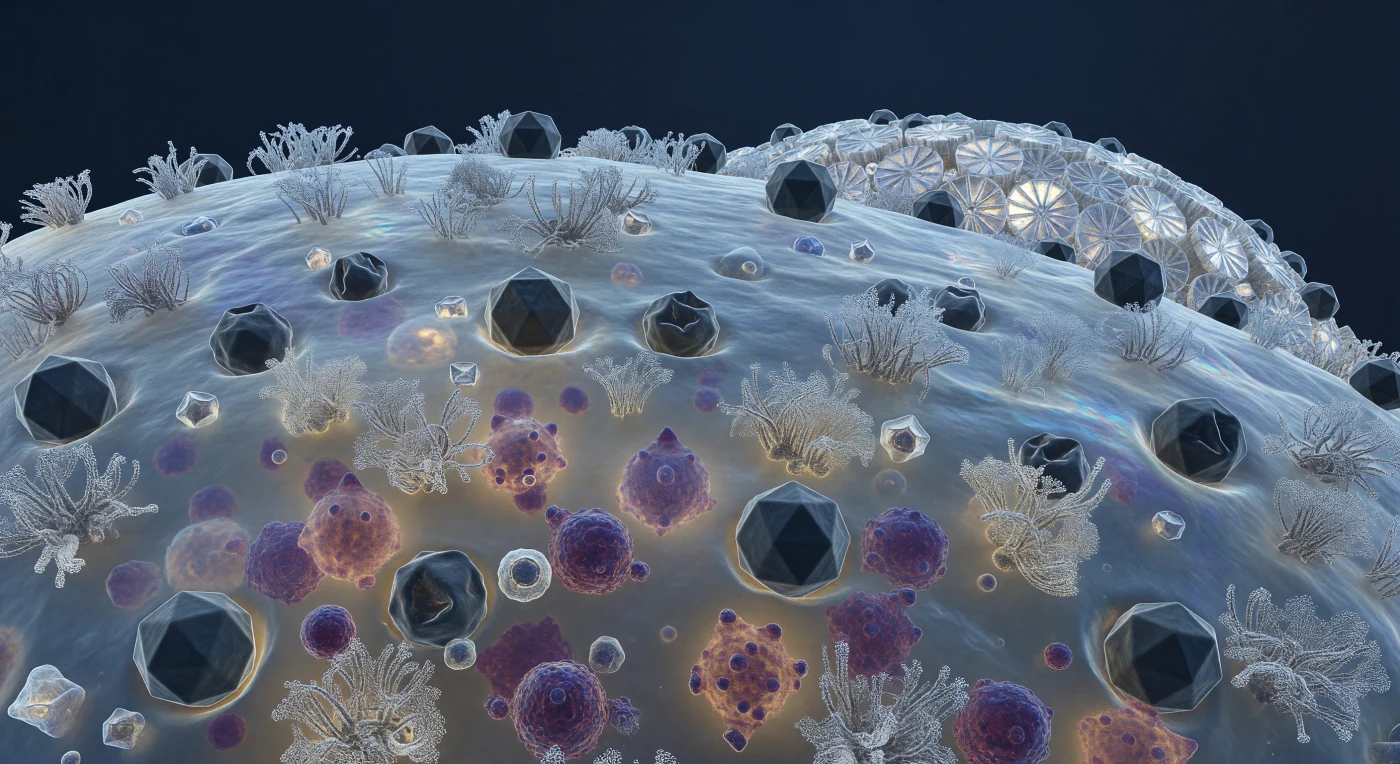
Desde apenas trescientos nanómetros de distancia, el observador se encuentra suspendido sobre una llanura biológica que lo abarca todo: la superficie exterior de una coccolitofora en plena crisis, donde la membrana plasmática se extiende como una piel azul-gris translúcida, temblorosa bajo la agitación browniana perpetua, salpicada de bosques de glicoproteínas receptoras que emergen como formaciones coralinas retorcidas y cubiertas de azúcar. Contra este terreno vivo se apretujan decenas de cápsides virales icosaédricas de EhV —bloques de pizarra oscura de apenas doscientos nanómetros, perfectos en su geometría de veinte facetas cuando recién llegados, aplastados y hundidos como cúpulas geodésicas reventadas cuando ya han inyectado su genoma— imprimiendo en la membrana pequeñas depresiones de rendición molecular. A través de esa pared translúcida, el citoplasma brilla como ámbar velado, y en su interior masas densas de color púrpura y magenta —nuevos viriones ensamblándose por millares sobre andamiajes de nucleoproteínas— presionan contra la membrana desde adentro, abombándola hacia afuera en los focos de replicación más intensa, mientras la célula es reescrita en silencio desde su propio interior. Hacia el horizonte curvo de la cocosfera, los bordes de las placas de calcita —ruedas blancas y cerámicas con rayos y aros precisos— se alzan como contrafuertes que dispersan en destellos prismáticos la poca luz fría que desciende desde metros de océano arriba, enmarcando una catástrofe íntima y extraordinariamente bella que se desarrolla un enlace molecular a la vez.

En las profundidades de cien metros bajo la superficie del océano subtropical, el observador flota suspendido en un crepúsculo índigo de una precisión espectral extraordinaria: la luz que llega desde arriba no es oscuridad ni claridad, sino una luminancia azul cobalto de 475 nanómetros que parece emanar del agua misma, como si el océano hubiera aprendido a autoemitirse. A esta escala —la de una sola célula de dinoflagelado, treinta micrómetros de altura— las criaturas del máximo de clorofila profunda se revelan como linternas de ámbar dispersas en el vacío frío: cloroplastos que arden en capas de membrana doradas, núcleos grises con cromosomas permanentemente condensados, frústulas de diatomeas transparentes como vidrio de sílice que atrapan los últimos fotones disponibles y los devuelven como destellos de brasa. Por encima de esta constelación viva existe una discontinuidad de índice de refracción —la termoclina— que distorsiona el resplandor lejano de las células superiores en lentes fantasmales, mientras que por debajo el índigo se extingue gradualmente en negro absoluto, texturizado únicamente por los chispazos blancos de nieve marina que desciende en espiral hacia el abismo, cada fragmento un edificio en miniatura de polisacáridos, escudos de calcita entrelazados de *Emiliania huxleyi* y materia biológica atrapada en hilos de mucus translúcido. Este es el límite entre la fotosíntesis y la oscuridad perpetua, una frontera que no corta sino que se desvanece: las células doradas se vuelven más escasas, su calor se atenúa, hasta que solo permanece el azul y luego nada.

Te encuentras suspendido en el milímetro más superficial del océano abierto, flotando junto a un *Ceratium tripos* que se alza ante ti como una catedral de ámbar translúcido: sus tres cuernos huecos —uno apical, dos antapicales— se extienden casi medio milímetro de punta a punta, suficiente para llenar tu campo visual como un autobús de ciudad visto desde la acera de enfrente. La superficie de la célula está revestida de placas de anfiesma que teselan su cuerpo en un panal de paneles alveolares iridiscentes, y los primeros fotones rosados y azul acero del alba, refractándose hacia abajo a través de la interfaz aire-agua que tiembla apenas dos longitudes de cuerpo por encima de ti, recorren el casco como llamas frías que pulsan sobre los cloroplastos interiores —discos dorados y cálidos empaquetados holgadamente dentro de la célula— en barreduras prismáticas lentas y cambiantes. En el ecuador de la célula, el flagelo transverso late en la ranura del cíngulo con un temblor plateado e hipnótico: un filamento casi invisible que, colectivamente, impulsa a este organismo unicelular en su ascenso diario hacia la luz mediante un mecanismo que la física de Reynolds tan bajo hace completamente distinto al nado macroscópico, pues aquí la viscosidad lo es todo y la inercia no existe. En la distancia media, enorme como una torre de cristal y suspendido en una bruma bioluminiscente azul-verdosa ya agonizante, un copépodo de un milímetro aguarda inmóvil: sus primeras antenas abiertas como remos translúcidos, su ojo captando la luz superficial como un ámbar oscuro y único, su escala tan aplastante que cada una de sus patas plegadas supera en tamaño al *Ceratium* entero que flota ante ti.

Te encuentras inmerso en una niebla viva de color jade, una suspensión tan densa de células de *Ostreococcus tauri* —cada una de apenas 0,8 micrómetros de diámetro— que el propio medio acuoso ha adquirido su tinte colectivo, como agua de mar vista a través de vidrio opaco. A esta distancia, la percepción de profundidad desaparece en cuestión de pocos largos de célula: lo que se encuentra a tres micrómetros de distancia ya está parcialmente velado por el banco de organismos interpuestos, cada uno de ellos una esfera casi perfecta cuyo tamaño rivaliza con la longitud de onda de la luz verde que los ilumina, oscilando en ese límite difuso entre partícula y onda. Las células más próximas se resuelven como pequeñas linternas de color esmeralda translúcido, con su único cloroplasto —tan grande que ocupa casi todo el interior celular— brillando en rojo profundo de autofluorescencia cuando un fotón lo excita en el ángulo preciso, antes de regresar al verde frío al continuar su rotación. *Ostreococcus tauri* es el eucariota fotosintético más pequeño conocido, desprovisto de pared celular y reducido a lo estrictamente esencial: membrana, cloroplasto, mitocondria y núcleo apretados en un volumen de menos de 0,3 femtolitros, un prodigio de compresión evolutiva que convierte el océano en un reactor bioquímico de escala invisible. El movimiento browniano es total e incesante, y la niebla verde respira sin pausa en todas direcciones, sin horizonte, sin suelo, sin techo: solo el pulso lento y probabilístico de miles de millones de seres que, sumados, tiñen el planeta desde el espacio.
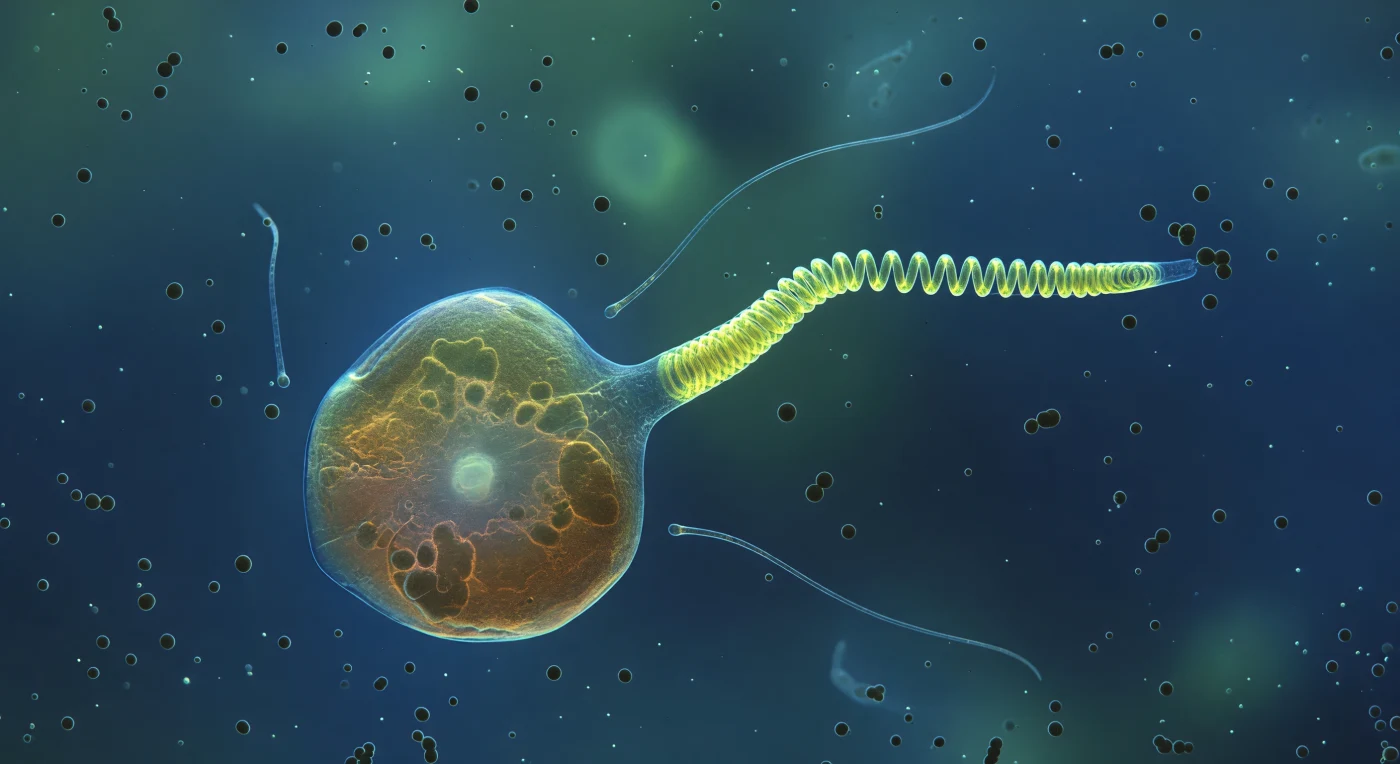
En este instante suspendido en el agua oceánica, el ojo se enfrenta a una *Chrysochromulina* cuyo cuerpo ambarino y semitransparente ocupa el campo visual como una arquitectura viva: la membrana plasmática vibra bajo la presión browniana del medio, mientras los cloroplastos proyectan sombras oscuras contra un núcleo pálido que brilla desde el interior como una lámpara cubierta de seda. Lo que domina la escena no es la quietud de la célula sino la violencia silenciosa que se desarrolla desde uno de sus polos: el haptonema —estructura única de los haptofitos, distinta de los flagelos en su arquitectura interna de microtúbulos dispuestos en anillo— se encuentra a medio camino entre dos estados físicos incompatibles, su tercio proximal ya colapsado en una hélice fluorescente amarillo-verdosa tensa como un resorte cargado, mientras el tramo distal aún persiste como varilla rígida que comienza a pandear en la zona de transición, liberando en milisegundos la energía elástica acumulada. A ambos lados del cuerpo celular, los dos flagelos —orgánulos locomotores de latido ultrarrápido, aquí completamente relajados— derivan como hilos de plata casi invisibles en la columna de agua, indiferentes al estallido mecánico que ocurre entre ellos. En el entorno inmediato, esferas bacterianas de un micrómetro flotan como planetas mate y oscuros, atrapadas en el gradiente químico que la célula proyecta hacia el agua circundante, mientras el medio mismo —lejos de ser vacío— dispersa la luz en una luminosidad lechosa de materia orgánica disuelta que convierte el fondo en un azul cobalto infinito y sin horizonte.

Desde la proa del buque, la mirada cae directamente sobre una de las fronteras más nítidas que ofrece la superficie planetaria: a babor, el océano índigo oscuro y transparente donde la luz se hunde metros hacia la oscuridad sin encontrar nada; a estribor, una masa de agua opaca, mineral, de un turquesa-blanco calcáreo que parece iluminada desde dentro, como si la columna de agua misma fuera la fuente del resplandor. Esa luminiscencia interna no es un efecto óptico superficial, sino la consecuencia directa de decenas de miles de millones de cocolitos —placas de calcita de dos a cuatro micras, cada una un prisma geométrico en rotación lenta— que dispersan los fotones del sol de tarde en todas las direcciones de manera casi lambertiana, convirtiendo el agua en algo parecido a piedra caliza pulverizada en suspensión. A lo largo de la línea de separación, la circulación de Langmuir ha ordenado la espuma en hileras paralelas que corren con el viento, marcando los bordes donde las dos masas de agua se rozan en remolinos horizontales invisibles; justo en ese límite, un alcatraz se cierra en flecha y perfora la superficie, y el géiser de agua blanca que levanta —capturado un instante por el sol bajo antes de desplomarse— parece la firma efímera de toda la productividad concentrada en esa franja de medio metro. En el aire flota un tenue dulzor azufrado, el dimetilsulfuro exhalado por las células de cocolitofóridos bajo estrés, y hacia el horizonte la distinción entre océano y cielo se disuelve en una neblina pálida producida por la dispersión colectiva de placas de calcita que se extienden hasta donde alcanza la vista.

Ante ti se extiende una sección transversal de un testigo de sedimento del fondo marino, diez centímetros de anchura que concentran millones de años de lluvia calcárea procedente de la zona fótica: una sucesión de láminas crema y hueso blanco que el sidelight rasante convierte en un relieve de cráteres y arcos diminutos, cada banda representando siglos de floración y colapso en las aguas superficiales. Eres del tamaño de un único cocólito, unos tres micrones de diámetro, y el mundo que te rodea es un laberinto impenetrable de ruedas de calcita entrelazadas con precisión cristalina, sus radios trigonales y sus bordes levantados captando la luz oblicua como ventanas de rosetón en el suelo de una catedral sumergida. Entre las ruedas más próximas, todavía íntegras, y las más profundas donde la diágénesis ha comenzado a suavizar los bordes y borrar los radios, se lee el tiempo como una transición del orden al olvido. Esta es la materia prima de la tiza de Dover: diez millones de coccolitofóridos por centímetro cúbico, su armadura de carbonato cálcico sobreviviendo a todo rastro orgánico de su biología, lípidos mineralizados, cloroplastos disueltos, pero la geometría de especie aún legible en la simetría cristalina para quien sepa mirar. El silencio es absoluto y total: ningún gradiente químico, ningún movimiento browniano, solo la densidad aplastante de los muertos que fundaron los continentes.
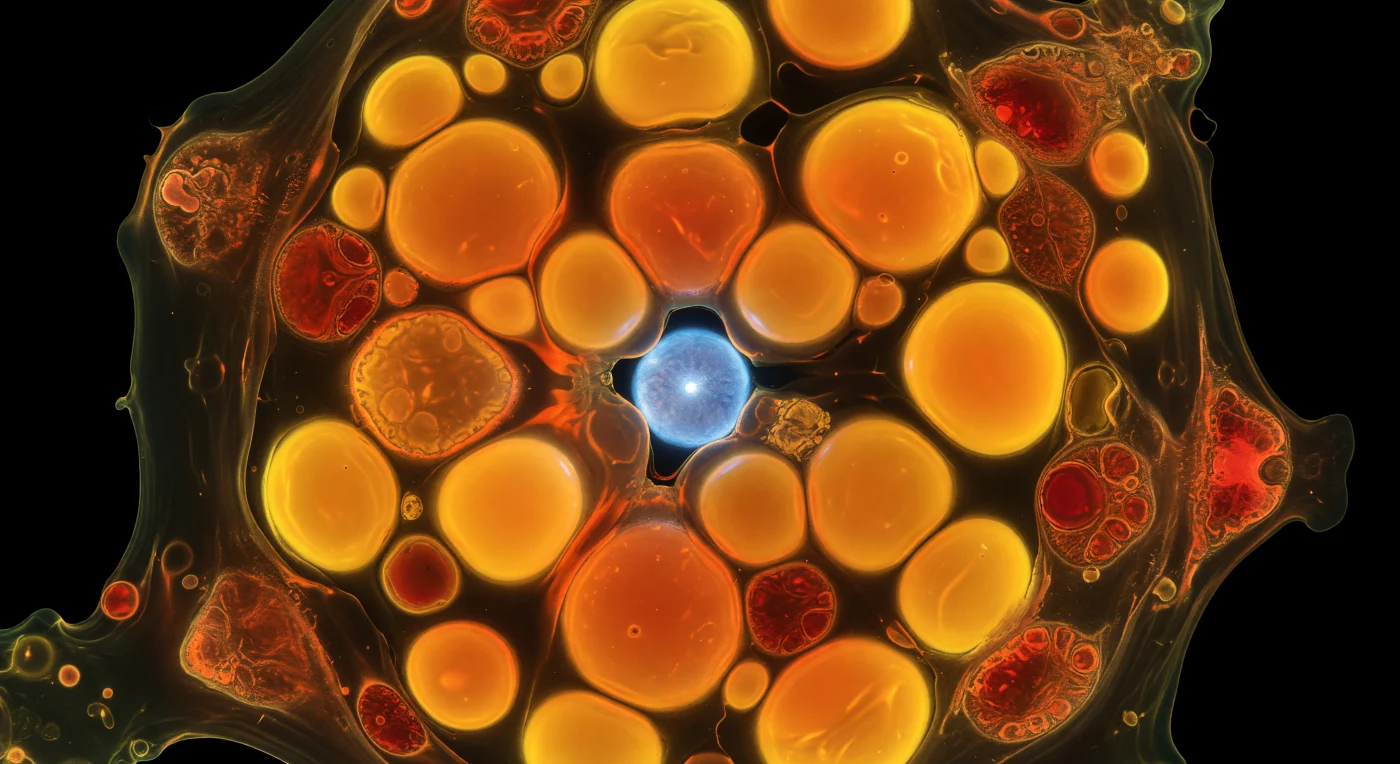
En el interior de una célula de *Emiliania huxleyi* sometida a inanición de nitrógeno, el espacio citoplasmático aparece dominado por esferas inmensas de lípidos que brillan con una intensidad anaranjada y dorada casi violenta: son gotas de grasa teñidas con Nilo Rojo, cuya fluorescencia no refleja luz exterior sino que la emite desde dentro, convirtiendo cada glóbulo en un pequeño sol autónomo que presiona contra sus vecinos y los deforma levemente en los puntos de contacto. A esta escala, el interior celular resulta claustrofóbico y denso, con estrechos canales de citoplasma viscoso serpenteando entre las esferas como ríos ambarinos atrapados entre planetas de lava, mientras los cloroplastos, empujados hacia la periferia por la marea lipídica, emiten una autofluorescencia roja pálida y menguante que apenas recuerda que esta célula fue, en otro momento, una máquina fotosintética activa. Este fenómeno de acumulación de lípidos es una respuesta adaptativa bien documentada: privada de nitrógeno, la célula redirige el carbono fijado por la fotosíntesis hacia la síntesis de lípidos neutros en lugar de proteínas y clorofila, almacenando energía densa en anticipación de condiciones aún más adversas. En algún punto entre los glóbulos ardientes, apenas visible a través de un corredor de luz ámbar, el núcleo resplandece con un azul eléctrico frío por la tinción DAPI sobre su ADN compactado, una luna cerúlea casi sepultada por la marea de aceite que la rodea en la oscuridad absoluta del exterior celular.

En la oscuridad absoluta, el océano ha dejado de tener superficie: agua negra y cielo negro se funden en un mismo vacío sin horizonte, y el cuerpo del observador existe solo como una perturbación en un medio vivo. Cada movimiento de las manos desencadena una descarga en cascada de millones de células de *Lingulodinium polyedra*, organismos unicelulares de seis a doce micrómetros cuyas proteínas luciferasa y luciferina, encapsuladas en organelos llamados escintilones, liberan fotones a exactamente 460 nanómetros, un azul eléctrico frío que los dedos dejan grabado en el agua como líneas de fuego que tardan dos segundos en desvanecerse, cartografiando vórtices y remolinos en luz viva. Las crestas de cada ola mantienen un pulso sostenido mientras el movimiento orbital del agua agita continuamente las células superficiales, revelando la geometría oculta del océano abierto en una topografía luminosa, y a cincuenta metros de distancia la estela de un barco ya desaparecido persiste como un río de incandescencia azul-blanca que parece casi sólido, trazado por un casco ausente sobre el agua negra. Este espectáculo no requiere luna ni estrellas: es la firma bioquímica de una reacción cuya antigüedad supera la de los animales, repetida simultáneamente en seres incontables que no distinguen entre la presión de una ola y el peso de un cuerpo humano que respira.
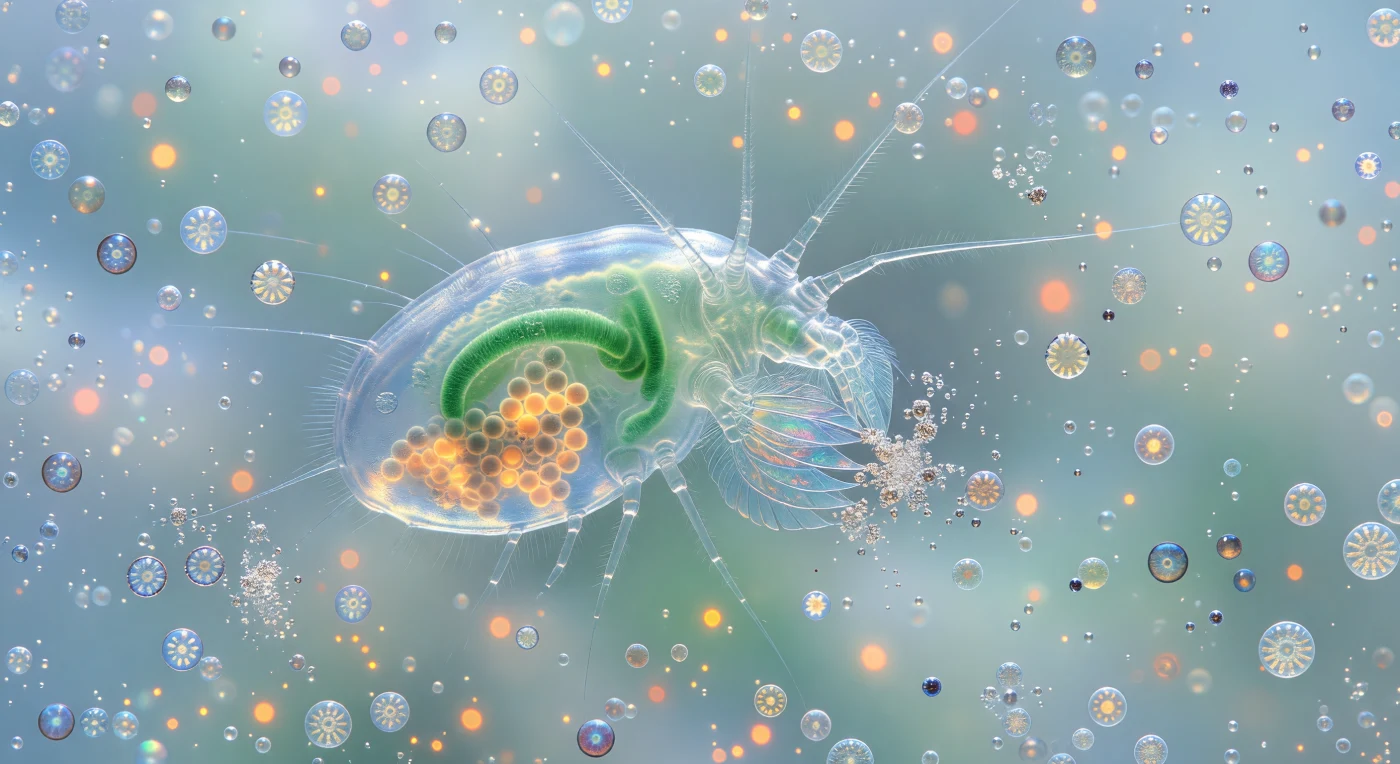
Flotas a dos milímetros de una arquitectura viviente: el copépodo *Calanus* se despliega ante ti como una linterna de quitina transparente, su intestino color jade y sus racimos de huevos ámbar ardiendo a través de tres capas de cutícula como brasas vistas a través de cristal esmerilado, cada órgano iluminado desde adentro por la luz oceánica transmitida. Sus maxilípedos —plumas de vidrio soplado armadas con setas del grosor de un cabello— vibran en el borde del campo visual, atrapando células de cocolitóforos individuales con la precisión de una telaraña que captura rocío, mientras la corriente de alimentación que genera el animal arrastra coccósferas intactas en arcos orbitales lentos, haciéndolas girar y destellar de plata a oro pálido según el ángulo entre sus placas de calcita y la luz difusa. Donde las puntas de los apéndices hacen contacto, las esferas calcáreas detonan en explosiones de purpurina blanca —cada *Emiliania huxleyi* destruida libera una lluvia de discos de calcita de dos micrómetros que tumban y relampaguean antes de ser barridos por la corriente—, mientras las células intactas que aún derivan exhiben una autofluorescencia naranja-rojiza desde sus cloroplastos, pequeñas constelaciones de calor metabólico moviéndose a través del medio turquesa frío. El agua circundante no es vacío sino una suspensión de leche luminosa —diez millones de cocolitos sueltos por mililitro dispersando la luz disponible en una neblina azul-blanca que disuelve toda perspectiva a tres largos de cuerpo de distancia—, convirtiendo la profundidad detrás del copépodo en una atmósfera infinita y gloriosa donde la densidad de la floración misma funciona como un cosmos de escala invertida: no hay horizonte, no hay superficie, solo la vasta presencia del bloom presionando desde todas direcciones mientras el animal organiza toda esa luz dispersa en vectores de aproximación y destrucción reluciente.

Estás suspendido en una columna de agua oceánica profunda, a la deriva en el azul frío y saturado del océano abierto filtrado por decenas de metros de columna, donde la gravedad apenas existe y el movimiento más pequeño es obra del azar molecular. A tu izquierda, la célula en fase C de *Emiliania huxleyi* domina tu campo visual como una joya arquitectónica: su cocosfera de placas de calcita blanco-crema se ensambla en una simetría casi imposible, cada cocólito un engranaje mineral de cuchillas cristalinas cuya geometría trigonal fractura la luz descendente en destellos fríos de blanco y oro pálido, toda la estructura brillando con una luminosidad difusa propia de la calcita dispersando fotones. A tu derecha, separada apenas por unos pocos diámetros celulares de columna abierta, su gemela haploide en fase N revela una materialidad radicalmente opuesta: sin armadura alguna, su membrana translúcida de verde-crema expone el calor dorado-ámbar de los cloroplastos en su interior, y sus dos flagelos —cables biológicos de fracción de micra capturados en el instante preciso de máximo arco— se curvan como filamentos de vidrio mojado contra el azul infinito del fondo. Esta yuxtaposición de dos estados del mismo organismo —uno mineralizado y reflectante, el otro desnudo y fotosintéticamente cálido— es la expresión visible de una alternancia de generaciones que conecta la bioquímica del carbonato oceánico con el clima global, pues las coccosferas que estas células producen y eventualmente liberan se hundirán durante siglos hasta formar sedimentos calcáreos en el fondo del océano.

Ante ti se eleva una pared casi vertical de creta cretácica que se pierde hacia el cielo gris-azul del Atlántico norte, su superficie encendida por una luz rasante de tarde que hace arder la calcita blanca hasta el límite del dolor visual — este acantilado es, en realidad, un archivo biogénico comprimido, construido grano a grano por centenares de millones de años de floraciones de cocolitóforos que vivieron, calcificaron y se hundieron hacia el fondo de antiguos mares subtropicales poco profundos, sus minúsculas placas de calcita acumulándose en una lluvia microscópica ininterrumpida hasta formar cientos de metros de roca biogénica. Las bandas oscuras de sílex que cortan la cara blanca en líneas horizontales casi perfectas son fantasmas químicos de la diagénesis del fondo marino, momentos de tiempo profundo incrustados en la estratigrafía como tinta sobre una página, su violento contraste tonal marcando el pulso de millones de años de sedimentación y transformación mineral. A mitad de altura, apenas un punto vivo sobre la masa geológica muerta, un fulmar inmóvil sobre una cornisa estrecha revela de golpe la verdadera escala del acantilado y lo vuelve vertiginoso e incomprensible: lo que parece textura porosa al alcance de la mano es, visto desde otra escala, la huella acumulada de organismos que midieron menos de una décima parte del ancho de un cabello humano, cuya geometría cristalina —ruedas de calcita de dos a cuatro micrómetros— se perdió en la compresión pero cuya masa colectiva sostiene ahora este monumento blanco e incandescente frente al mar.