Confianza científica: Alto

Desde el fondo de una repisa de roca calcárea del tamaño de un plato, la mirada asciende a lo largo de treinta columnas de jade translúcido que se alzan desde una superficie blanquecina y craterada, casi lunar en su palidez, donde cada organismo no es una planta ni una colonia sino una sola célula gigante que ha construido por sí misma esta arquitectura de tallo cilíndrico y sombrilla radiada. Cada disco terminal —una ventana de roseta gótica dividida en segmentos gametangiales con precisión geométrica— se convierte en una pequeña linterna cuando la luz oblicua de la mañana mediterránea lo atraviesa desde atrás, encendiéndolo en chartreuse cálido y amarillo paja que se profundiza hacia un verde lima intenso en el centro, toda la textura celular revelada en transmisión como vitral iluminado. La pared del tallo más cercano, a pocos centímetros del ojo, muestra un ligero estriado en espiral y, en el flanco orientado al sol, el fantasma de una migración cloroplástica —un oscurecimiento casi imperceptible del verde que delata el desplazamiento lento del citoplasma fotosensible dentro de este ser unicelular de varios centímetros de alto. El agua por encima de los discos es un cerúleo profundo cargado de partículas en suspensión que atrapan el haz oblicuo y crean una neblina volumétrica suave, con la superficie lejana visible como un espejo ondulante de luz plateada. La escena oscila entre catedral y pradera submarina —las cápsulas como óculos de vidrio emplomado, el suelo blanco como losa de piedra caliza, el turquesa overhead como cielo— todo ello contenido en un fragmento de arrecife que cabe en la palma de la mano.

Suspendido apenas cuatro centímetros sobre la superficie inmóvil de la poza mareal, el ojo se convierte en instrumento óptico: el agua actúa como lente perfecta que amplifica y congela el mundo bajo ella, revelando una alfombra viva de *Caulerpa racemosa* que se extiende sobre basalto negro como un continente de esferas jade apretadas, cada rámulo un cúmulo de glóbulos de uno a dos milímetros cubiertos por una cera glauca que atrapa el sol cenital en destellos blancos y duros —mil estrellas en miniatura sobre un manto verde azulado. Lo que el ojo percibe como colinas, valles y horizontes es en realidad la topografía de un organismo unicelular gigante: toda la colonia de *Caulerpa*, con sus estolones, sus holdfasts y sus frondas erizadas, constituye una única célula cenocítica, un solo citoplasma continuo rodeado por una sola membrana plasmática que puede extenderse varios metros sin una sola tabique interno. Las redes cáusticas del sol —haces de luz refractados en la superficie— se mueven y disuelven sobre la alfombra algal con lentísima cadencia, mientras en los márgenes de la poza la esponja incrustante naranja ofrece un contraste cálido y poroso frente a la regularidad botánica de los rámulos esféricos. La paradoja de escala es total: lo que la biología define como célula, el ojo lo experimenta como paisaje habitable, con su propia gravedad, sus propias sombras de cobalto y su horizonte teal que se desvanece hacia las profundidades templadas de la cuenca.

Nos encontramos a pocos centímetros del fondo marino mediterráneo, a doce metros de profundidad, con la mirada al ras de una alfombra monoespecífica de *Caulerpa taxifolia* que se extiende hasta el límite visual en todas direcciones, cada fronda un único organismo cenocítico —una sola célula sin tabiques internos— cuyo citoplasma continuo recorre el talo entero desde los rizoides anclados en la arena hasta las últimas pínulas. La geometría pinnada de cada fronda se repite con precisión mecánica: el raquis, no más grueso que una aguja de tejer, despliega docenas de pínulas planas y emparejadas capaces de interceptar la luz azul difusa que desciende desde la superficie en columnas cáusticas que barren diagonalmente los ápices y revelan, por transparencia, la vacuola interna y el córtex granular verde oscuro. La saturación cromática del conjunto —un jade eléctrico que se disuelve en teal y luego en ultramar a medida que el agua absorbe las longitudes de onda más largas— convierte la escena en algo próximo a una catedral acuática de vidrio tintado, interrumpida únicamente por una concha de gasterópodo blanqueada que descansa sobre la arena entre los estolones, su superficie calcítica devolviendo un pequeño hemisferio del mundo circundante como una perla apagada, testimonio mudo de la uniformidad alienante que la rodea.

Te ciernesobre una grieta de caliza a cinco metros de profundidad, y los quince globos de *Valonia ventricosa* que la habitan te miran de vuelta como canicas de vidrio soplado bajo una presión imposible: cada esfera de dos a cuatro centímetros de diámetro es, en su totalidad, una sola célula viva, una arquitectura coenocítica en la que millones de núcleos flotan en un citoplasma continuo contenido por una pared de celulosa y manano enrollada en capas de fibras cruzadas que le dan a la superficie ese lustre sedoso apenas distinto del vidrio puro. La red de cáusticos —filamentos de luz caribeña refractada desde la lámina de mercurio que tiembla sobre tu cabeza— recorre las superficies esmeraldas en reorganización continua, cada nodo una estrella que florece y se disuelve en menos de un segundo, mientras la translucidez de la pared deja entrever la vacuola central como una linterna de jade autoluminosa, el cloroplasto ectoplásmico presionado desde dentro como una piel viva de un milímetro. Un gusano de cerdas —cobre iridiscente, sus parapodios destellando como nácar— serpentea entre dos de las esferas mayores en la esquina inferior de la grieta, y su cuerpo, que en cualquier otro contexto sería el protagonista de la escena, aquí queda eclipsado por la masa de lo que no es más que una célula: la escala se invierte, la biología se vuelve absurda, y la certeza de lo que constituye un organismo se disuelve con la misma fluidez con que la luz baila sobre el coral coralino rosa.
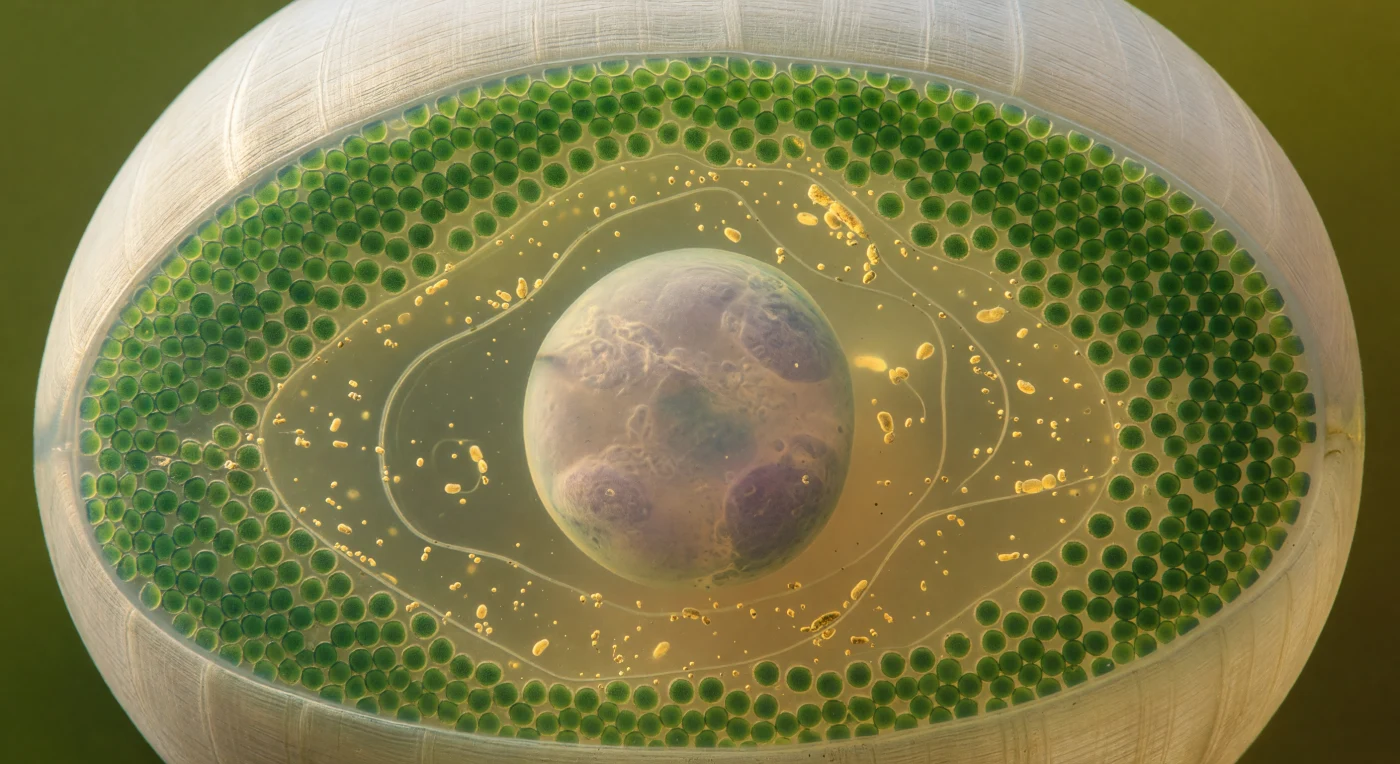
Suspendidos en el interior de la cámara rizoidal de una célula de *Acetabularia*, nos encontramos inmersos en un medio ámbar-verdoso que llena el espacio en todas direcciones como agua somera bañada por luz antigua, atravesada por la claridad del océano que filtra desde el exterior a través de la pared celular. Esa pared se curva a nuestro alrededor como el interior de un pequeño globo translúcido —crema-blanca, finamente estriada por las microfibrillas de celulosa cruzadas— y sobre su cara interna se aprieta un mosaico compacto e ininterrumpido de cloroplastos, discos de jade esmeralda que forman una vidriera viva y saturada, cada uno irradiando su propio verde profundo contra la contraluza cálida. En el centro de este mundo cerrado flota el núcleo primario: una esfera de casi ochenta micrómetros, gris pálido con matices lavanda, cuya membrana no impone un límite rígido sino una frontera luminosa y suave que dobla el resplandor ámbar en un halo que se enfría hasta el gris-plata en su superficie, mientras que en su interior el nucléolo reposa como una piedra oscura en agua turbia. Entre la pared y el núcleo, el citoplasma está recorrido por la ciclosis: gránulos dorados y ámbar, junto a filamentos del retículo endoplasmático, trazan arcos lentos y deliberados alrededor del núcleo en corrientes tan pausadas que parecen deriva geológica antes que flujo, recordando que esta célula única —sin tabiques internos, gobernada por un solo núcleo gigante— orquesta toda su fisiología desde este espacio que ahora habitamos.

Te encuentras dentro del corredor hueco de un ser vivo que desafía toda intuición sobre lo que puede ser una sola célula: el lumen de un estolón de *Caulerpa*, apenas dos milímetros de diámetro, se abre ante ti como la nave de una catedral gótica sumergida en luz esmeralda, sus paredes curvas encendidas desde dentro por miles de cloroplastos apretados que emiten una luminiscencia verde oscura con destellos dorados y aguamarina allí donde la densidad del pigmento se adelgaza. Cruzando el espacio vacuolar en todas direcciones, las trabéculas —hebras citoplásmicas traslúcidas de unas pocas decenas de micrómetros de grosor— avanzan en perspectiva hacia la oscuridad profunda del fondo, formando una sucesión de arcos apuntados y arbotantes orgánicos que recuerdan la geometría del gótico tardío pero que aquí son tensiones vivas, estructuras de refuerzo que sostienen el lumen abierto contra la presión de turgencia de la célula. Este organismo es un cenocito: un ser sin tabiques internos en el que miles de núcleos comparten un mismo citoplasma continuo que se extiende desde los rizoides hasta las frondas, unificado por la corriente lenta de la ciclosis, ese río viscoso que arrastra hacia ti los gránulos de almidón y los orgánulos como motes de ámbar suspendidos en la quietud luminosa del vacuolo. Una sola célula construyó esta arquitectura, la mantiene viva y la recorre entera en una continuidad sin fronteras internas, contradiciendo con su existencia todo lo que el tamaño suele significar en biología.

El vehículo no tripulado se mantiene suspendido a apenas veinte centímetros del fondo del mundo, a cinco kilómetros de profundidad, donde la presión del agua convierte cada movimiento en un acto de precisión mecánica y la temperatura roza el punto de congelación del mar. Directamente delante, iluminado por el cono duro y azulado de los paneles LED del ROV, se alza *Syringammina fragilissima*: un montículo irregular de quince centímetros de diámetro construido grano a grano por una única célula colosal, una xenofiofor que ha aglutiando fragmentos de foraminíferos y partículas minerales en una arquitectura rugosa y piteada que imita la escala de una cadena montañosa en miniatura. La luz oblicua talla sombras de bordes afilados en cada cavidad de la superficie, tiñendo de carbón y ámbar oscuro la textura extraordinary del organismo y convirtiendo sus oquedades más profundas en pozos de negrura absoluta, mientras los fragmentos de concha capturan destellos especulares fugaces como astillas de porcelana rota. El sedimento que se extiende más allá — un gris ceniciento y finísimo, surcado por trazas de gusanos y microóndulas labradas por corrientes abisales imperceptibles — se disuelve en oscuridad total a menos de medio metro, una oscuridad que no es ausencia de luz sino presencia física de todo el océano apilado sobre este instante. Partículas de nieve marina derivan en silencio a través del cono iluminado, casi inmóviles, suspendidas sobre esta célula única que lleva tiempo geológico ensamblando pacientemente su propia geología en uno de los rincones más solitarios de la biosfera.

De pie en el centro exacto de esta esfera viviente, el mundo se curva en todas las direcciones al mismo tiempo: una catedral sellada cuyas paredes son la propia célula, una única *Valonia ventricosa* de tres centímetros de diámetro que encierra un universo completo en la lógica perfecta de una burbuja. La superficie interior de la membrana vacuolar forma un horizonte continuo que envuelve todo el campo visual, un mosaico denso e ininterrumpido de cloroplastos aplastados contra el ectoplasma cortical que tiñe la esfera entera de verde esmeralda y verde botella, una alfombra luminosa y ligeramente granulada de pigmento fotosintético que brilla con intensidad diferente según la exposición a la luz exterior, brillante en el ecuador, más oscura y jade en los polos. Más allá de ese manto vivo, apenas entrevistas a través de él, las capas cruzadas de microfibrillas de celulosa dibujan un enrejado de espiga en crema y marfil, bandas diagonales dispuestas en ángulos precisos y opuestos que confieren a la pared transparente la profundidad estructural de un tejido fino tensado sobre un vacío luminoso. El fluido vacuolar que llena todo este espacio interior no es agua corriente sino un líquido de precisión química extraordinaria, amarillo pálido como paja con un matiz de miel, cuya altísima concentración de potasio es invisible al ojo pero se percibe en la calidad del medio: más denso, más quieto, más silencioso que cualquier océano abierto, transmitiendo la luz verde del manto clorofílico sin apenas dispersión hasta convertir la iluminación en una luminiscencia omnidireccional sin sombras que envuelve cada objeto desde todos los ángulos simultáneamente. De vez en cuando, pequeñísimas vesículas o cristales iónicos derivan con lentitud glacial por la savia, captando la luz ambiental como motas de oro y plata que apenas se mueven, suspendidas en la quietud monumental de una arquitectura biológica mantenida en equilibrio perfecto por la presión de turgencia.

En la penumbra cálida y sedimentada del fondo costero, la pared de sedimento aparece como una sección geológica perfectamente revelada, dominada por el estolón de *Caulerpa* — un cilindro tenso y lacado de cuatro milímetros de diámetro que brilla en un verde esmeralda profundo, tan geométricamente liso y presurizado desde su interior que parece vidrio soplado más que tejido vivo, y cuya particularidad más extraordinaria es que todo este organismo visible — estolón, rizoides y frondes — no es sino una sola célula gigante, un coenocito continuo sin tabiques internos que puede extenderse metros en el sustrato sin interrumpir jamás su citoplasma unificado. Descendiendo desde su cara inferior, la red de rizoides se despliega en ramificaciones fractales de ámbar translúcido, cada filamento primario de apenas cien micras de ancho, dividiéndose simétricamente en rootlets cada vez más finos que abrazan granos individuales de cuarzo y feldespato como raíces de un árbol alienígena que coloniza un paisaje mineral a escala de bolígrafo. La turgor interna de toda esta arquitectura coenocítica mantiene la forma estructural sin esqueleto celular convencional, mientras los cloroplastos migran dentro del citoplasma común respondiendo a gradientes de luz — abundantes y densamente comprimidos contra la pared en la zona iluminada del estolón, dispersos y escasos en los rizoides más profundos donde la luz ya llega convertida en un último fulgor ámbar-marrón antes de extinguirse en la oscuridad granular. Suspendido a la altura de un grano de arena, el espectador percibe el gradiente lumínico del sedimento como una cronología comprimida: luz costera difusa y ligeramente verdosa en la interfaz superior, seguida por una penumbra cada vez más cálida y densa que convierte los últimos filamentos rizoidales visibles en siluetas fantasmales atrapadas en una oscuridad suave e infinita.

Ante ti se elevan tres pilares translúcidos desde un fondo de aragonita blanquecina, cada uno una única célula viva que ha construido su propio cuerpo centímetro a centímetro sin dividirse jamás: la más joven luce una corona de pelos citoplasmáticos que dispersan la luz lateral en filamentos celádón luminosos, la intermedia muestra un proto-casquete hinchado cuyos primordia radiales aún negocian su forma definitiva, y la madura despliega un disco perfecto cuyo borde upturneado arde como vitral esmeralda contra el azul oceánico. Cada tallo está impregnado de carbonato cálcico secretado por la propia célula, una biomineralización que convierte tejido vivo en arquitectura mineral de color menta pálida, mientras la pared celular sostiene presiones de turgencia capaces de mantener erguida toda esa altura sin ningún sistema vascular ni esqueleto multicelular. El sol raseante de la tarde atraviesa los tejidos desde la derecha y revela, en gradaciones de jade y oro, la distribución de cloroplastos que tapizan el córtex como un mosaico fotosintético continuo, mientras el núcleo gigante —único y relegado al rizoides enterrado bajo la arena— gobierna a distancia toda esta complejidad morfogenética mediante gradientes de ARN mensajero que ascienden por el citoplasma como instrucciones enviadas desde una sala de máquinas invisible. La sombra circular que el casquete maduro proyecta sobre el sustrato blanco es quizás el detalle más perturbador: una geometría tan precisa, tan resuelta, ejecutada por una sola célula que lleva semanas construyéndose a sí misma.

Suspendido a la altura de una canica junto al punto de ramificación más alto de un arbusto de *Halimeda*, el observador se encuentra cara a cara con discos de aragonita del tamaño de su palma abierta: superficies mate y granulosas como caliza erosionada, empolvadas de diatomeas ocre que centellean como brasas ámbar en la luz azul-verdosa filtrada por ocho metros de columna de agua. Cada segmento calcificado es, en realidad, parte de una sola célula gigante ceonócita —sin tabiques internos, con miles de núcleos flotando en un citoplasma continuo— cuya pared exterior ha mineralizado aragonita en una costra rígida mientras los nodos de articulación permanecen sin cristalizar, intensamente verdes y flexibles, llenos de cloroplastos que emiten un suave resplandor esmeralda cuando la luz de fondo los atraviesa. La arquitectura fractal se despliega en todas direcciones: cadenas de discos que se bifurcan y rebifurcan en curvas lentas impulsadas por el pulso residual del oleaje, desprendiendo una fina harina de carbonato que deriva como motas plateadas y efímeras en la penumbra cerúlea. Más allá, las cintas de *Posidonia* se disuelven en un bokeh de jade y oro pálido, una pared viva y abstracta que convierte este minúsculo universo mineral-orgánico en un espacio de precisión estructural extraordinaria, suspendido en silencio dentro de la pradería submarina.

Suspendido en la columna de agua, el espectador es testigo de una explosión biológica que se despliega en cámara lenta: el cuerpo desgarrado de una *Ventricaria ventricosa*, una sola célula del tamaño de una canica, yace como una tienda de campaña derrumbada sobre el arrecife, su pared celular de microfibrilas de celulosa —gruesa, correosa, levemente iridiscente— todavía rezumando savia mientras sus bordes se curvan hacia adentro. De su interior surge una constelación de protoplastos hijos, esferas casi perfectas de entre una décima y un milímetro de diámetro, cada una cargada en su córtex con cloroplastos dispuestos como adoquines bajo una membrana translúcida, que las tiñen de un verde esmeralda profundo en las sombras y de un dorado citrino encendido donde los rayos tropicales las atraviesan en ángulo rasante. Este evento —denominado segregación espontánea o fisión múltiple— es el mecanismo reproductivo de esta gigantesca unicelular: la célula madre libera decenas o cientos de propágulos capaces de asentarse y desarrollarse en nuevos individuos autónomos, cada uno con su propia bioquímica completa encerrada en una membrana que apenas supera el grosor de una pompa de jabón. La distorsión refractiva del sap celular difundido alrededor de cada esfera curva suavemente los colores rosados y malvas del coralino que recede en el fondo, convirtiendo todo el volumen de agua en un sistema óptico vivo donde la física del vidrio y la del mar se funden en una respiración silenciosa.
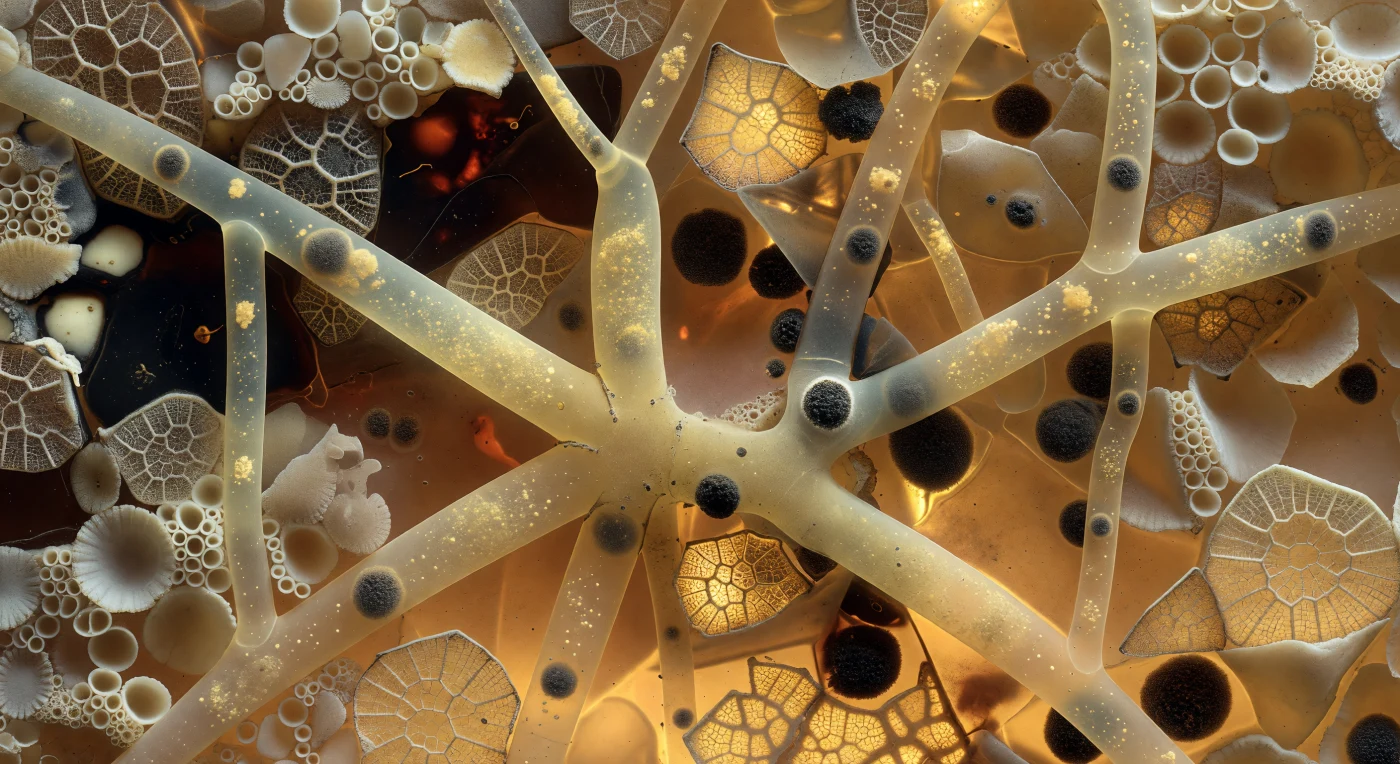
Suspendido en el interior iluminado del esqueleto mineral de una xenofiópora, el observador se encuentra rodeado por un mosaico de fragmentos conquiliológicos —paredes de foraminíferos compactas como losas de marfil y retículas de radiolarios cuyos finos armazones de sílice capturan la luz ámbar transmitida desde abajo, todo cementado por una matriz orgánica oscura y ligeramente lustrosa que une el conjunto como resina antigua solidificada. Esta arquitectura no es fósil sino la envoltura viva de un ser unicelular de proporciones extraordinarias: la *teca* o *test* de una xenofiópora, construida a lo largo de su vida por una única célula gigante que agrega y consolida detritos del fondo oceánico en una estructura que puede alcanzar varios centímetros de diámetro. Atravesando este laberinto mineral, los canales llamados *linelas* se ramifican en todas las direcciones como corredores de vidrio ámbar translúcido de cuarenta a ochenta micrómetros de anchura, transportando el citoplasma dorado pálido compartido de esa célula multinucleada, cuyas diminutas masas de cromatina grafito flotan a intervalos irregulares como piedras sumergidas en una corriente de miel. La luz cálida que atraviesa la preparación histológica convierte el campo entero en algo suspendido entre la geología y la biología, entre la catedral y la célula, revelando que lo mineral y lo vivo no son aquí categorías separadas sino capas superpuestas de una misma entidad.
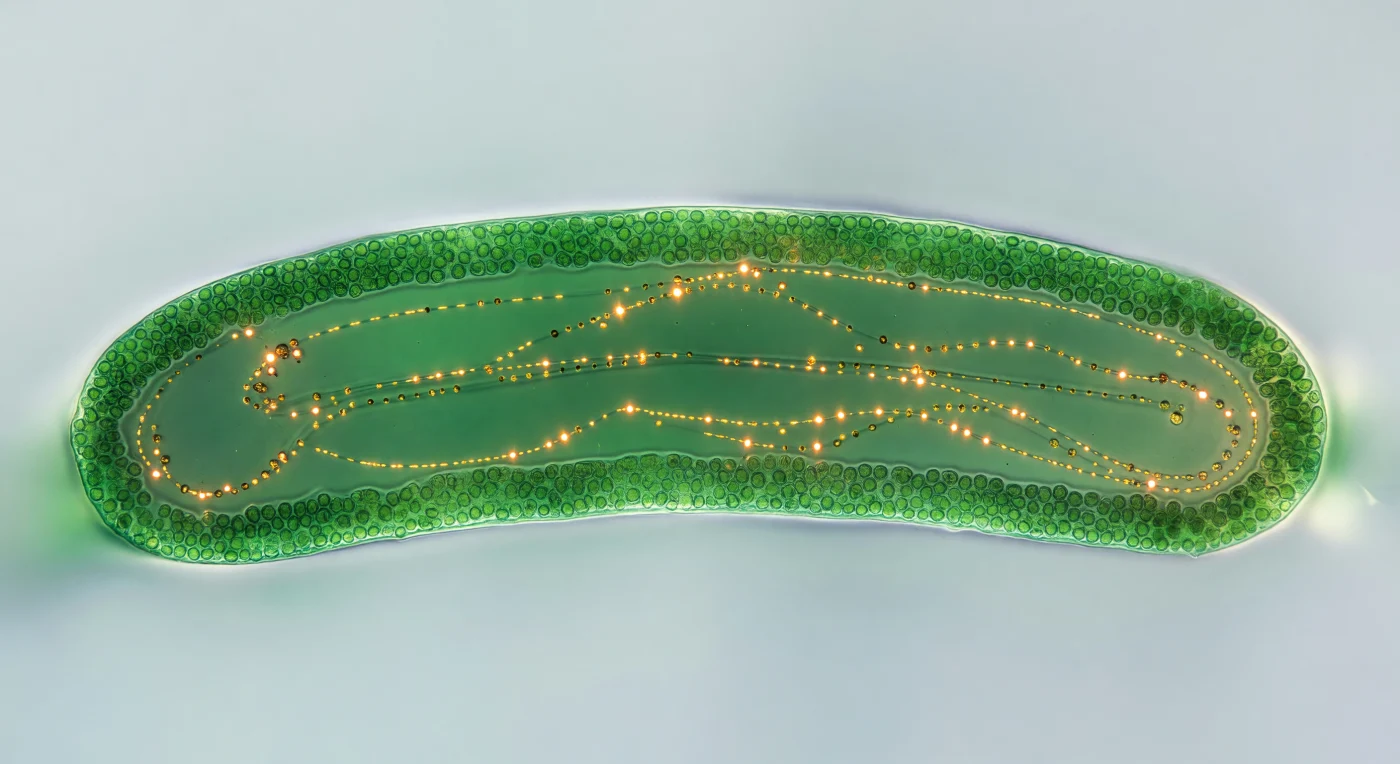
Suspendido a escasos centímetros sobre ella, el ojo abarca de borde a borde un mundo verde que ocupa todo el campo visual: un único segmento de estolón de *Caulerpa*, una célula solitaria de dos milímetros de anchura, que reposa en una película de agua marina sobre un cristal bañado por una luz blanca transmitida desde abajo. Esa iluminación convierte la célula en una linterna arquitectónica viva, cuya corteza exterior arde en esmeralda y malaquita saturados —un manto continuo de cloroplastos tan apretados que forma una muralla de jade— mientras el interior se abre en un corredor más oscuro y frío, la nave de una catedral tallada en vidrio biológico, todo ello contenido en una única célula sin tabiques internos, un citoplasma continuo que se extiende sin interrupciones a lo largo de todo el organismo. Dentro de ese canal endoplásmico, el movimiento existe pero no se anuncia: gránulos ámbar y oro avanzan en largas trayectorias arqueadas a la velocidad de una respiración lenta, deslizándose sobre cables de actina invisibles a apenas uno a diez micrómetros por segundo, algunos congelados como puntas de luz naranja, otros extendidos en arcos luminosos de ocre que trazan la geografía de las autopistas citoplasmáticas. La profundidad de campo es brutal en su estrechez: la superficie superior del estolón se disuelve en bokeh verde mientras el plano iluminado permanece afilado como un corte, de modo que la luz misma parece leer la arquitectura interior de la célula, revelando en un instante todo el abismo que separa lo visible de lo verdaderamente pequeño.

En la oscuridad casi absoluta del fondo rocoso mediterráneo, tres tallos de *Acetabularia* se elevan desde la costra calcárea como columnas de marfil traslúcido, cada uno una célula única e ininterrumpida de varios centímetros de altura, emanando una fosforescencia residual de clorofila del color del fuego fatuo visto a través de cristal esmerilado. Cada tallo es un ser unicelular gigante que desafía toda intuición sobre lo que puede ser una célula: su interior, apenas visible como una niebla granular de cloroplastos suspendidos en gel citoplasmático, contiene un único núcleo colosal que reposa como una esfera pálida en el rizoide anclado a la roca, gobernando la arquitectura de todo el organismo a distancia. Cuando una cascada de dinoflagelados perturbados libera destellos de luz azul-blanca que descienden por la columna de agua como grietas químicas en movimiento lento, la geometría radial de los sombreros apicales queda revelada en toda su precisión arquitectónica: membranas apenas más gruesas que una pompa de jabón, sus bordes festoneados lanzando sombras duras y rotantes sobre los tallos mientras la luz dispersada penetra brevemente las paredes celulares y expone la organización viva en su interior. Entre destello y destello, el agua misma tiene densidad y textura —partículas de materia orgánica en suspensión capturan cada pulso luminoso como una galaxia lenta de chispas individuales— y los tres tallos regresan a ser lo que son en la oscuridad: linternas biológicas solitarias, cuyo resplandor esmeralda apenas alcanza a calentar el vacío azul-negro que los rodea.
