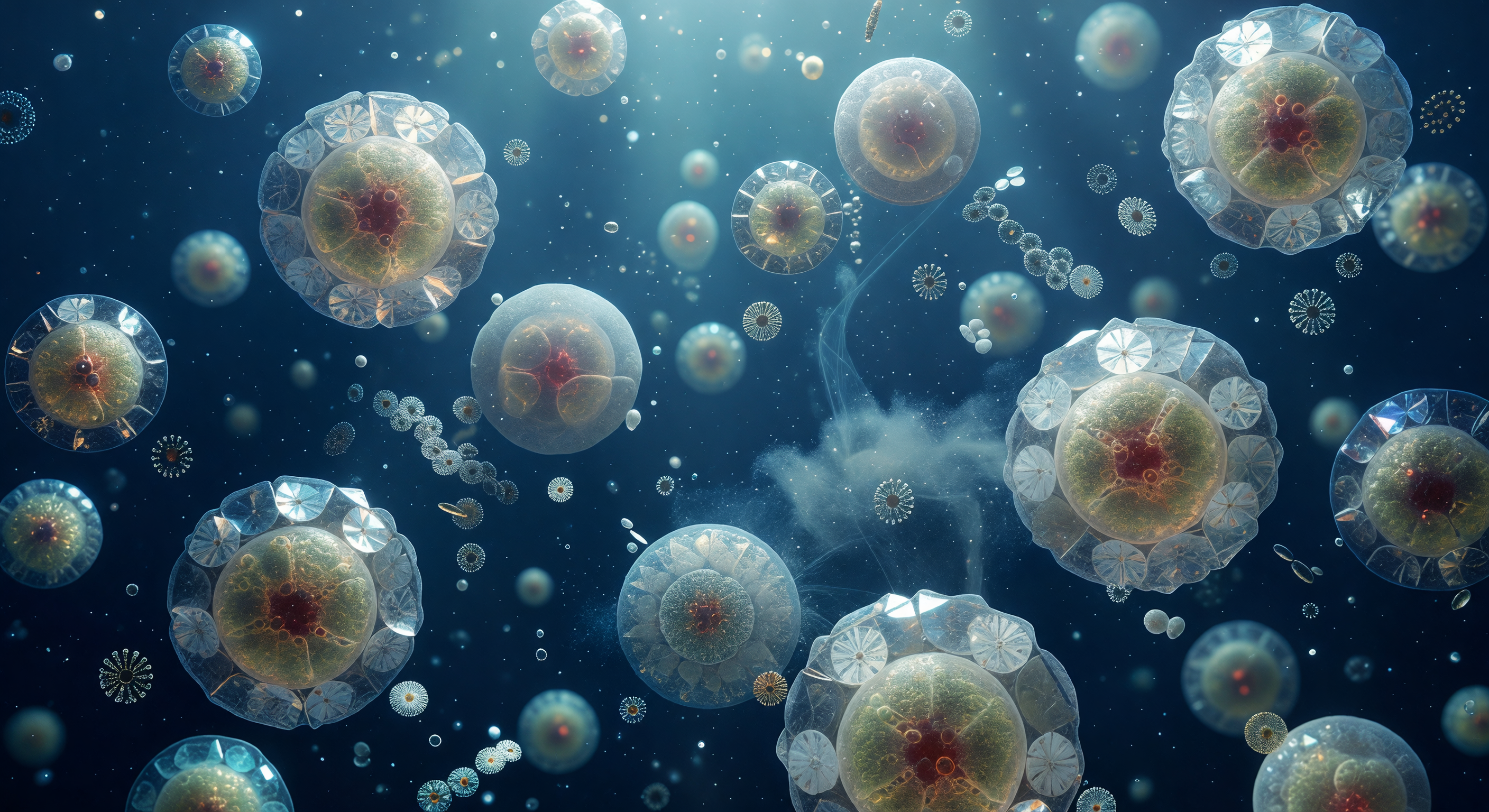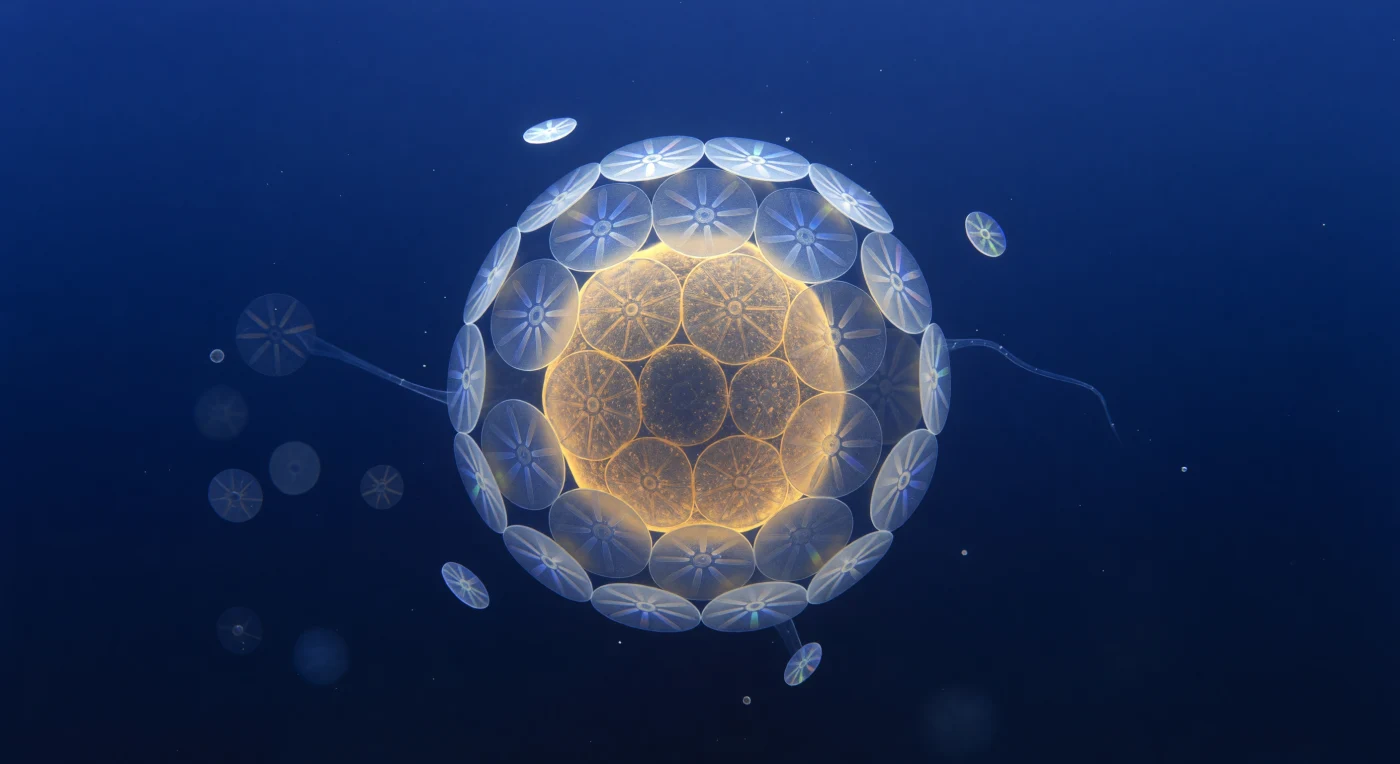
Você flutua a distância quase nula de uma única célula de *Emiliania huxleyi*, que ocupa todo o seu campo visual como uma lua âmbar suspensa no azul ultramarine mais profundo que qualquer oceano à superfície jamais exibiu — uma esfera translúcida de oito micrómetros irradiando uma luminescência interna dourada e açafranada, onde os cloroplastos comprimidos filtram a luz como vitrais de mel. Em torno desse núcleo vivo, vinte cocólitos de calcite entrelaçam-se com precisão de relojoaria, cada placa uma roda de raios cristalinos que converge num centro miniaturizado, e em cada junção a luz descendente de 480 nanómetros — já filtrada por metros de coluna de água — fragmenta-se em estilhaços espectrais fugazes: azul gelo, violeta, um lampejo de verde-menta na aresta de um cristal e depois o silêncio. Esta armadura de carbonato de cálcio, chamada cocosfera, não é apenas proteção: é a prova mineral de um organismo que regula o ciclo global do carbono, bombeando calcite para o fundo do oceano ao longo de milénios e acumulando, comprimido, nas falésias brancas que hoje emergem do continente. À sua periferia, cocólitos soltos giram imperceptivelmente no ruído térmico browniano, e filamentos de exopolímero esticam-se como vidro fiado através do campo médio — tudo envolto num índigo que se aprofunda abaixo, denso e antigo, como se a própria água guardasse memória da luz que nunca chegou até aqui.
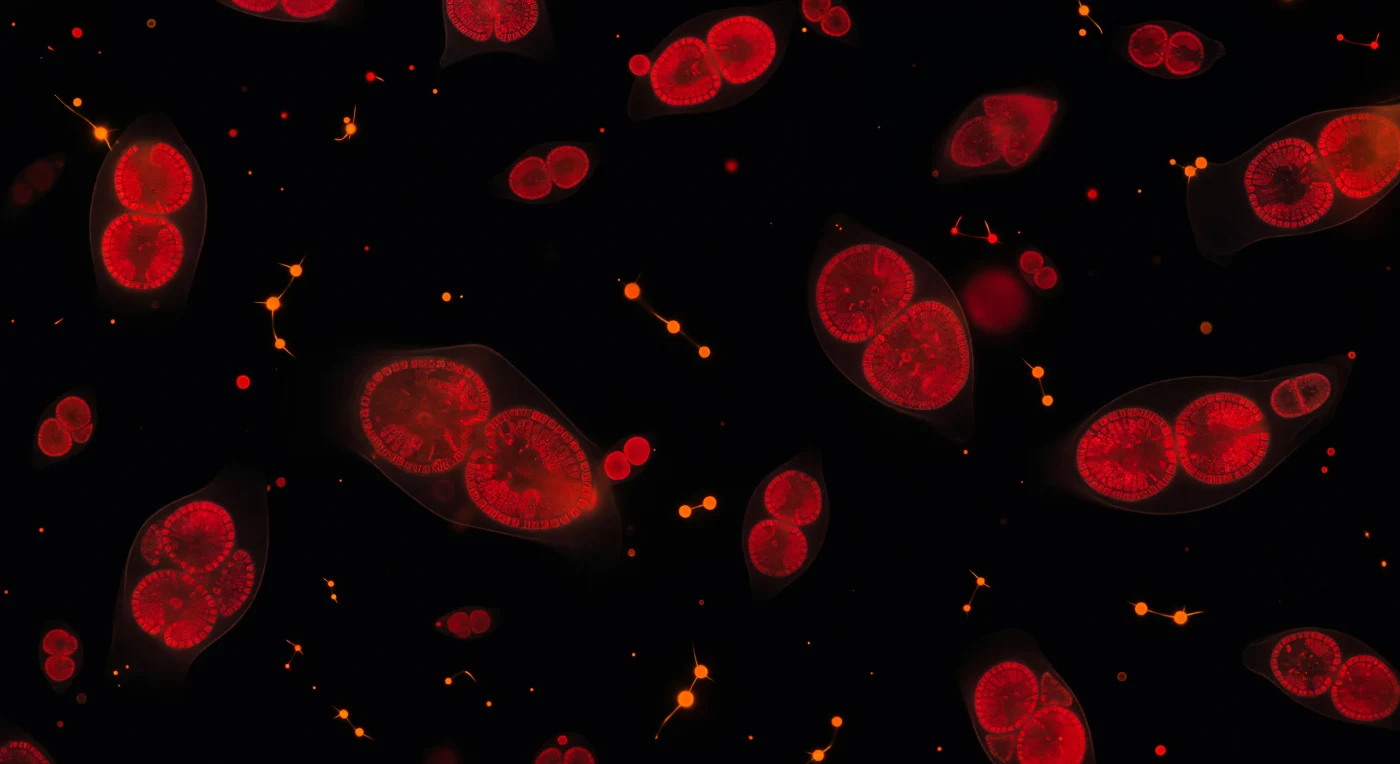
Flutuamos numa escuridão absoluta onde cada fonte de luz é gerada de dentro para fora: as células de fitoplâncton ardem em carmesim profundo à nossa volta, a sua autofluorescência de clorofila emitindo a 680 nm como brasas concentradas suspensas no vazio, sem horizonte nem gravidade percetível, apenas a deriva térmica lenta do movimento browniano a transportar cada organismo numa trajetória silenciosa e sem atrito. No interior de cada célula semitransparente, os cloroplastos bilobados resolvem-se em formas reniformes escarlates, densamente empacotadas com membranas tilacóides empilhadas em folhas lamelares cujas dobras mais espessas aprofundam a cor até ao bordô — estruturas que convertem fotões em energia química com uma eficiência que nenhuma tecnologia humana igualou. Entre estas nebulosas vermelhas, as cianobactérias punctuam a escuridão como faíscas laranja-tangerina, a sua ficoeritrina a queimar numa frequência distinta e mais quente, algumas alinhadas em cadeias que atravessam o meio líquido como pequenas constelações prokariotas. O fluido em si permanece invisível mas presente: halos difusos de matéria orgânica dissolvida e vesículas de membrana captam a luz celular e dispersam-na em coroas avermelhadas de apenas um micron de espessura, confirmando que habitamos uma suspensão aquosa e não o vácuo que a escuridão sugere. Neste campo visual medido em millionésimos de metro, cada célula é uma estrela autoluminosa, e a profundidade tridimensional entre elas — real, mensurável, vasta à sua escala — replica com fidelidade assombrosa a estrutura de um campo estelar observado em plena noite.

Você está até o peito naquelas águas do Atlântico Norte durante o auge de uma floração de *Emiliania huxleyi*, e o oceano ao redor deixou de ser água — tornou-se uma suspensão mineral viva, opaca como giz diluído, iluminada de dentro pelo espalhamento de dezenas de milhões de rodas de calcita por mililitro. A superfície se estende em todas as direções como uma extensão plana e ofuscante de jade-branco e creme-turquesa, um meio tão carregado de geometria microscópica que a luz não consegue penetrar mais do que a espessura de uma mão antes de ser espalhada de volta ao céu. Fileiras de Langmuir riscam a superfície em fitas paralelas cor de leite condensado, cada uma delas a assinatura visível de vórtices contra-rotativos subsuperficiais que concentram as partículas mais leves em linhas espessas orientadas pelo vento, espaçadas de dois a quatro metros. Na linha do horizonte, o limite da floração traça uma fronteira impossível: à esquerda, o branco calcário; à direita, o azul cobalto escuro do oceano aberto, nítido como tinta, como se dois planetas distintos tivessem sido pressionados lado a lado. Olhando para baixo, através de apenas cinco ou seis centímetros de relativa clareza, seu próprio corpo desaparece numa névoa luminosa verde-branca — abaixo desse limiar, cada fóton perdeu toda a memória da direção de onde veio.
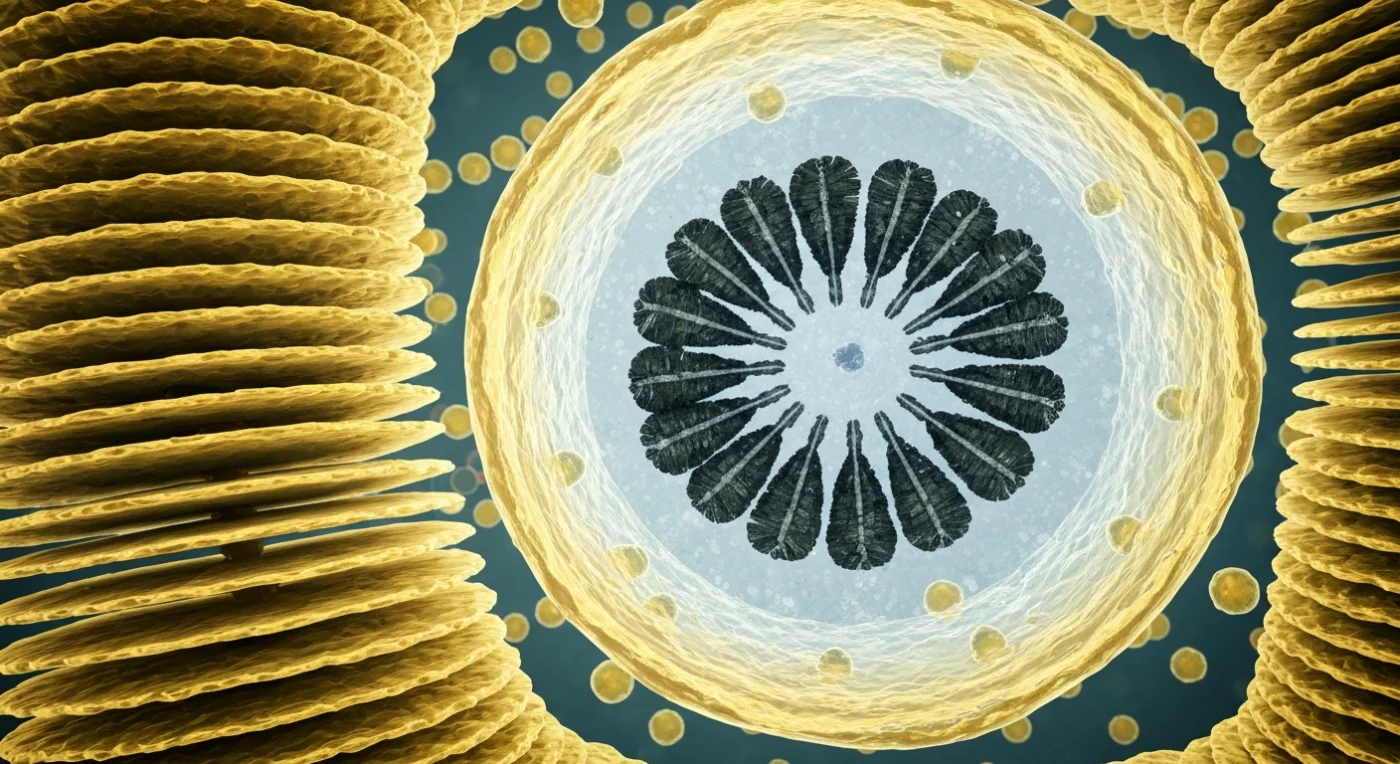
Ao seu redor ergue-se uma catedral de membranas douradas empilhadas em folhas paralelas e precisas — as cisternas do Golgi de *Emiliania huxleyi*, cada uma com apenas algumas dezenas de nanómetros de espessura, translúcidas e levemente luminescentes com o âmbar quente das bicamadas lipídicas, curvando-se suavemente nas extremidades onde a face trans incha em vesículas que se desprendem e derivam pela citoplasma como lanternas flutuantes, transportando precursores de calcite para a câmara de montagem adiante. Dominando o campo visual como uma cúpula industrial colossal, a vesícula do cocólito ocupa o primeiro plano — um único compartimento membranoso de parede âmbar translúcida, cujo interior aquoso emite uma fria luminescência azul-acinzentada sobre uma placa base de polissacárido, fina e levemente iridescente como seda congelada. Contra essa estrutura orgânica, unidades de calcite em cinzento-carvão dispõem-se em anel radial — trinta prismas com facetas internas que captam a luz em reflexos oblíquos prateados, interlocking com uma precisão que parece engenharia, embora seja apenas química aquosa; um quadrante permanece aberto, os cristais ali mais curtos e rugosos nos seus topos onde a mineralização continua em tempo real, iões de cálcio chegando em pulsos invisíveis através da membrana. O espaço entre as cisternas está densamente ocupado por ribossomas que pontuam as superfícies do retículo endoplasmático em constelações repetidas, dando a cada superfície uma textura aveludada e áspera, enquanto mitocôndrias alongadas pulsam em laranja-vermelho no penumbra granular do fundo — nada aqui está vazio, cada nanómetro cúbico de citoplasma é território, e a fábrica opera em silêncio absoluto, construindo pedra a partir do mar.

Diante de você, suspenso na escuridão absoluta, uma cocosfera arde como um sol capturado: quinze rodas de calcita entrelaçadas formam uma esfera quase perfeita, e cada cocólito flameja sob a luz polarizada cruzada como um mandala radiante, com o padrão de extinção em cruz de Malta varrendo cada placa em negro aveludado enquanto os domínios cristalinos adjacentes incandescentes em branco de primeira ordem parecem ter pressão interna. Nas bordas espessadas dos raios e nos escudos distais elevados, a interferência de filme fino eleva a cor para um dourado pálido e um marfim alaranjado quente, como luz de vela contra a neve, revelando onde as diferenças de percurso óptico empurram a retardância ligeiramente além do branco puro. Ao redor desta esfera central, cocólitos individuais desprendidos derivam em todas as direções numa lenta translação browniana, cada um uma roda elíptica de calcita com apenas 3 µm de diâmetro — plana como folha martelada, porém arquitetonicamente ornada — projetando sua própria explosão estelar de birrefringência contra o campo negro, alguns de perfil como finas luas crescentes luminosas, outros de frente revelando a simetria perfeita de uma catedral cristalográfica em miniatura. Esta geometria intrincada não é obra de engenharia, mas de biomineralização: a célula viva secreta íon a íon cada prisma de calcita dentro de uma vesícula intracelular, construindo no escuro e no frio das águas oceânicas uma armadura que acaba por sedimentar-se no fundo marinho e, ao longo de milhões de anos, compor os grandes depósitos de calcário e giz que afloram hoje em continentes.
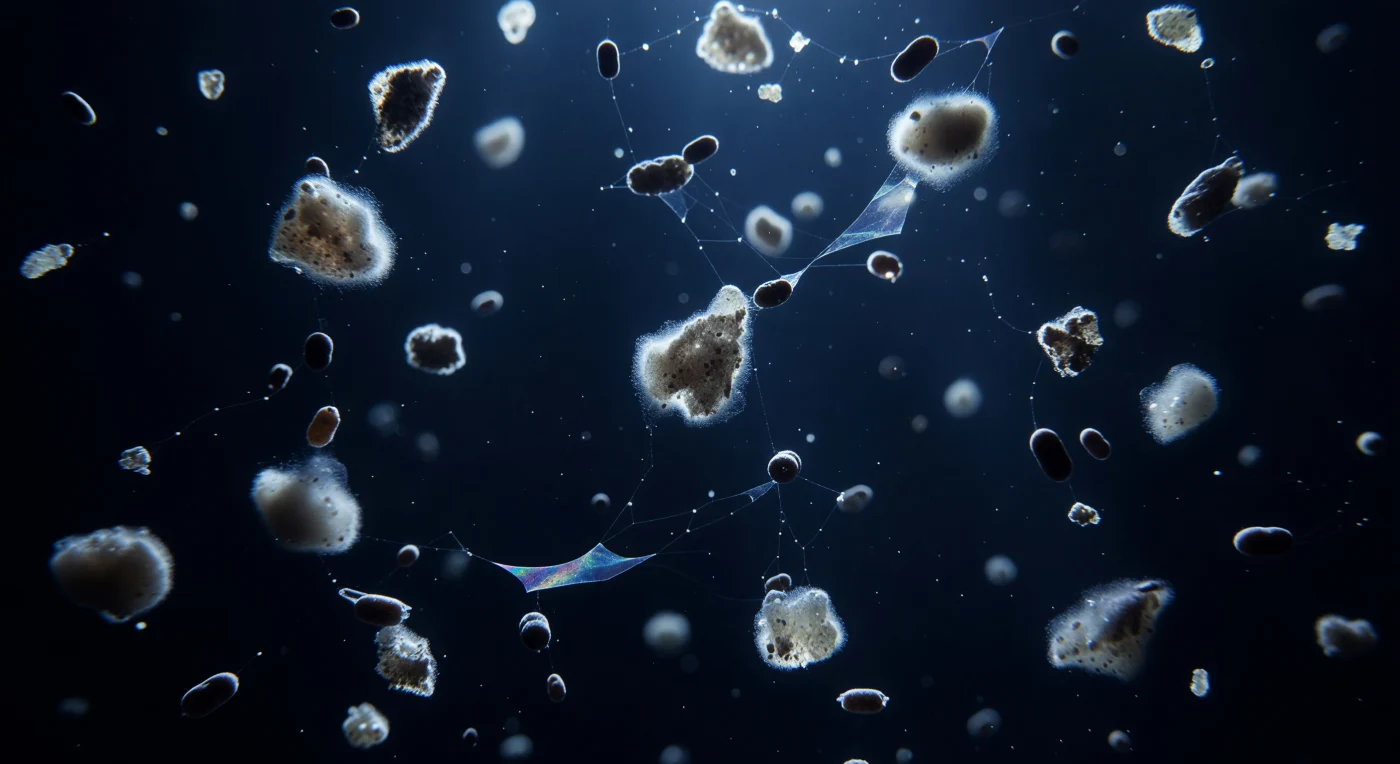
Suspenso a duzentos metros de profundidade, o observador existe dentro de uma coluna de água índigo-escura quase absoluta, atravessada por uma procissão vertical interminável de flocos brancos-acinzentados que descem em câmara lenta extrema — agregados de neve marinha entre um e quatro milímetros, construídos ao longo de dias a partir de cocolitos calcários, frústulas de diatomáceas mortas, muco e detritos celulares compactados em matéria biológica em colapso lento. Os últimos fótons que filtram da zona eufótica, atenuados entre 450 e 490 nanómetros, retroiluminam cada floco num halo azul-branco fantasmagórico, fazendo cintilar os fragmentos de calcite dispersos no seu interior como lanternas geladas que se apagam antes de chegar ao fundo. Entre os flocos, pellets fecais cilíndricos de cor castanha-escura descem três a dez vezes mais depressa, perfurando o campo de neve como pequenos projéteis densos e lisos, enquanto filamentos de partículas exopoliméricas transparentes — TEP — esticam-se entre os agregados em véus iridescentes quase invisíveis, revelando-se apenas pelo modo como refratam a escassa luz nas suas margens. Esta tempestade de matéria orgânica, silenciosa e sem testemunhas, representa a bomba biológica de carbono do oceano em funcionamento: o arquivo sedimentar da vida à superfície, escrito floco a floco numa descida que não pára há centenas de milhões de anos.
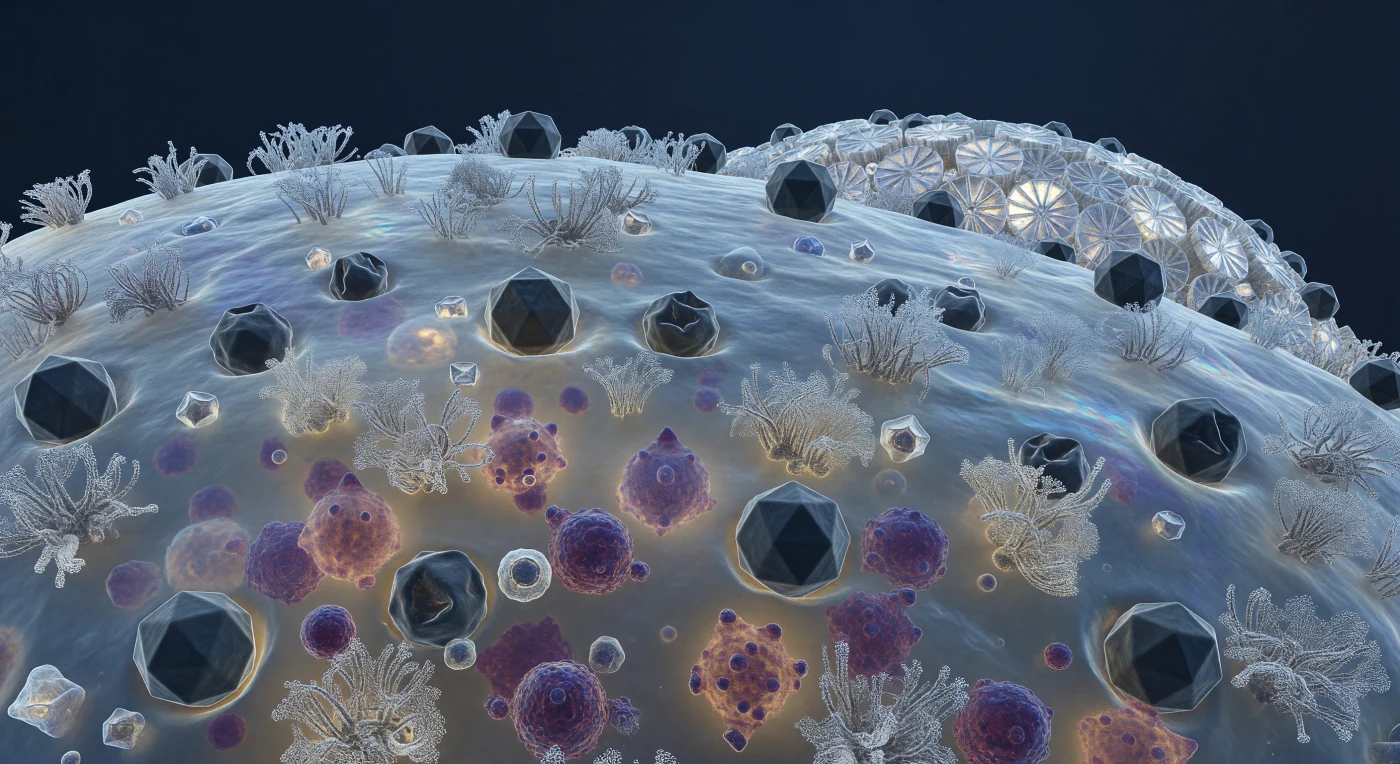
Diante de nós estende-se uma paisagem de escala continental — embora meça apenas micrómetros — a superfície curvada de uma coccolitófora sob assalto viral, onde dezenas de cápsides icosaédricas do vírus EhV-86, cada uma com cerca de 200 nanómetros de diâmetro, pressionam os agrupamentos de recetores glicoproteicos como pedregulhos assentes numa costa viva. Algumas cápsides preservam a sua geometria perfeita de vinte faces triangulares, as arestas captando a luz fria e difusa do oceano profundo em flashes de cinzento-ardósia; outras jaz colapsadas e amachucadas, a simetria rompida após a injeção do genoma na célula hospedeira, a membrana lipídica por baixo arqueada em pequenas depressões de rendição molecular. Através da pele translúcida azul-acinzentada da membrana — que tremula com a agitação browniana e exibe manchas iridescentes onde os domínios de radelas lipídicas se concentram — massas densas de púrpura e magenta iluminam o interior citoplasmático como âmbar embaciado: novos viriões em fase de montagem que preenchem compartimento após compartimento, empurrando a membrana para fora em bolsas subtis onde a replicação atinge maior intensidade. No horizonte desta superfície biológica, as placas de calcite cristalino das cocólitas erguem-se como contrafortes cerâmicos brancos, as suas geometrias de raio e aro dispersando a escassa luz em lampejos prismáticos — beleza geométrica extraordinária a encobrir uma catástrofe silenciosa que se desenrola molécula a molécula.

Suspenso a cem metros de profundidade no oceano aberto subtropical, o observador existe à escala de uma única célula de dinoflagelado — cerca de trinta micrómetros de altura — imerso numa penumbra de azul cobalto espectralmente puro, onde apenas dois por cento da irradiância superficial consegue penetrar como um brilho difuso e quase intangível que parece emanar da própria água em todas as direções. Neste crepúsculo índigo, dezenas de células fitoplânctónicas derivam em constelação lenta: dinoflagelados que ardem em dourado-azeite quente contra o frio da coluna de água, as suas membranas anfiesomais levemente prismáticas a refratar os fotões disponíveis, enquanto cadeias de diatomáceas em filamentos espiralados reluzem com as suas paredes de sílica transparente e conteúdos castanho-dourados comprimidos como brasas. Esta é a camada de clorofila profunda — uma zona de acumulação biológica onde o fitoplâncton explora o limite inferior da luz utilizável, capturando fotões de 475 nanómetros com pigmentos ajustados à escassez, num equilíbrio delicado entre fotossíntese e custo metabólico da manutenção em profundidade. Passando lentamente, um agregado de neve marinha do tamanho de um edifício na referência do observador desce em silêncio absoluto — uma teia translúcida de exopolímeros, placas de cocólitos ainda entrosadas nas suas formas geométricas de roda calcítica, peletes fecais compactos e células meio-vivas presas em fios de muco — enquanto mais abaixo o azul se extingue em gradiente suave para o índigo verdadeiro e depois para a escuridão total, onde apenas centelhas brancas de neve marinha distante capturam os últimos fotões antes de espiralar para o abismo.

Diante de vós, suspensa no milímetro mais superficial do oceano aberto antes do amanhecer, ergue-se uma única célula de *Ceratium tripos* como uma catedral de âmbar translúcido, os seus três cornos ocos de celulose e calcite a estenderem-se quase meio milímetro de ponta a ponta — à vossa escala, o comprimento de um autocarro urbano visto do outro lado de uma rua silenciosa. A superfície da célula é um mosaico de painéis alveolares iridescentes, cada costura a capturar os primeiros fotões do crepúsculo pré-auroral como um fio de fogo azul-branco, enquanto as colunas de luz rosa e aço-azulado que entram pela interface ar-água acima varrem o interior e fazem pulsar os cloroplastos dourados em ritmos lentos e prismáticos. No sulco cingular que circunda o equador da célula como um canyon esculpido, o flagelo transversal bate em fita helicoidal contínua, invisível como evento isolado mas colectivamente visível como um frémito prateado que aparece e desaparece no índigo da água — ao seu lado, o flagelo longitudinal desce para a escuridão como um único fio de vidro. No meio da cena, a três ou quatro centímetros reais de distância mas percepcionado aqui com a massa de um edifício de escritórios em vidro, um copépode paira em imobilidade — o corpo quase completamente transparente, as antenas abertas como remos de cristal, cada perna segmentada individualmente maior do que toda a célula de *Ceratium* à vossa frente — e a água entre vós dois guarda ainda um halo de bioluminescência azul-esverdeada, eco de células perturbadas momentos antes, que se dissolve lentamente na luz crescente que transforma toda a coluna de água de índigo profundo a um cobalto translúcido atravessado por fios de ouro.

Você está imerso numa névoa viva de tom jade, uma suspensão tão densa de células de *Ostreococcus tauri* — cada uma com apenas 0,8 micrómetros de diâmetro — que o próprio meio aquoso parece ter adquirido cor e substância, tingido coletivamente pela clorofila de bilhões de organismos em simultâneo. Estas células são os menores eucariotas fotossintéticos conhecidos, tão próximos do limite de resolução da luz visível que cada uma oscila na fronteira entre partícula e onda, a sua membrana lipídica de sete nanómetros produzindo halos espectrais iridescentes que tremem e desaparecem à medida que a célula roda na corrente térmica. O cloroplasto único de cada célula ocupa quase todo o seu interior como uma gema de jade escuro encerrada numa esfera de vidro, e quando a luz de excitação os atinge em ângulo favorável, liberam um breve rubor de autofluorescência vermelha — a assinatura da clorofila — antes de regressarem ao seu verde fresco e translúcido. O movimento browniano é absoluto e omnipresente: nada repousa, cada célula deriva em micro-correntes térmicas invisíveis, e a profundidade lê-se não como distância mas como saturação de cor, as células próximas nítidas como lanternas viridinas, as mais afastadas já fantasmas dissolvidos na luminescência coletiva do nevoeiro. Esta névoa viva estende-se em todas as direções sem horizonte nem chão, uma substância viva em suspensão que é simultaneamente organismo e oceano.
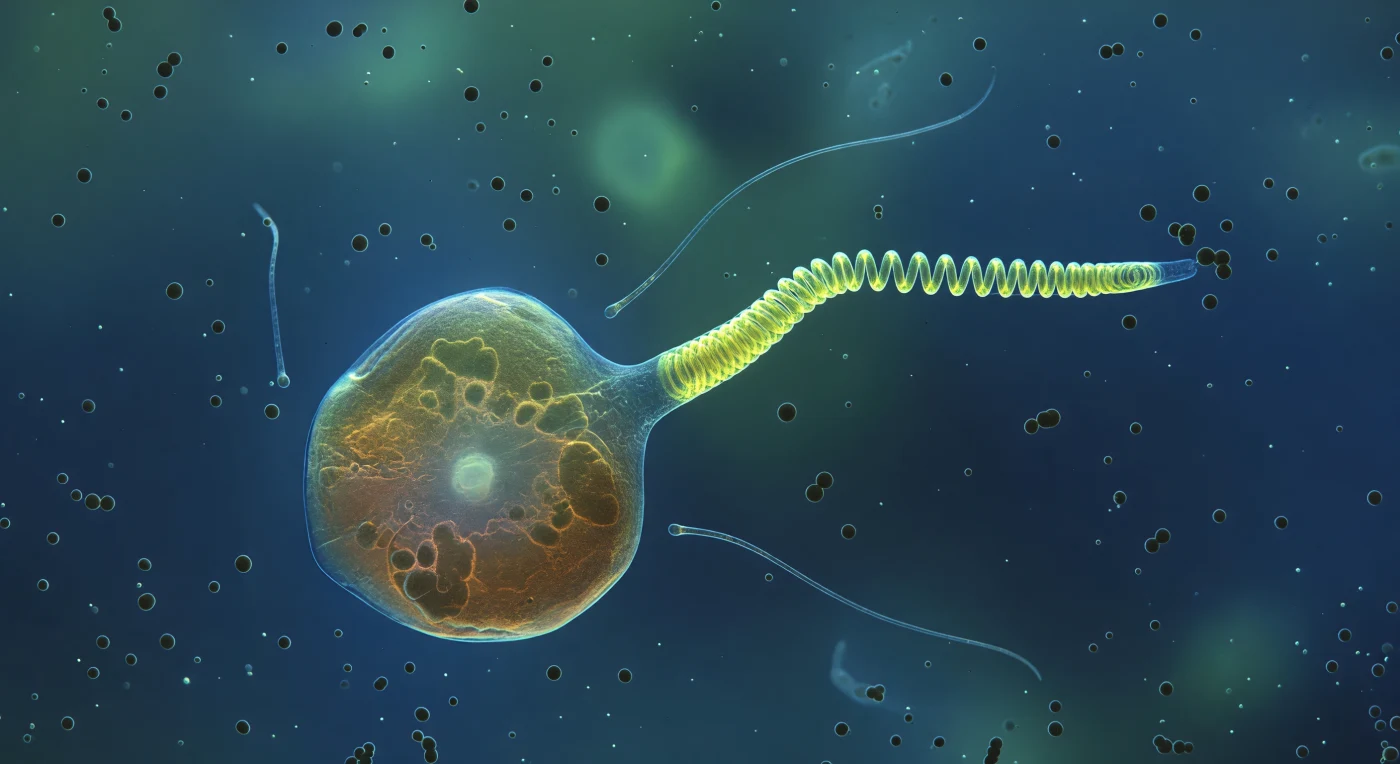
Diante de você abre-se um universo translúcido banhado por uma luz azul-esverdeada e difusa, fria e ligeiramente fosforescente, como se o próprio oceano emanasse claridade de todas as direções ao mesmo tempo. O centro desta visão é ocupado por uma célula de *Chrysochromulina* de proporções monumentais a esta escala — um globo âmbar-dourado e semitransparente, com cloroplastos escuros pressionando contra a membrana interna e um núcleo pálido brilhando no centro, a superfície celular iridescente como uma película de sabão sob pressão browniana constante. De um dos polos da célula, o haptonema irrompe em plena transformação violenta: o terço proximal já colapsou numa espiral helicoidal densa e incandescente de amarelo-verde fluorescente, enquanto a porção distal permanece como uma haste rígida em vias de flectir na zona de transição, toda a estrutura suspensa no instante exato em que a energia biofísica acumulada se liberta numa fracção de segundo — um mecanismo de recuo ativado por alterações do citoesqueleto interno que transforma microtúbulos organizados numa mola comprimida. Ao redor da ponta em movimento, esferas escuras e opacas de cerca de um micrómetro derivam lentamente no gradiente químico que emana da célula maior, bactérias atraídas pelos compostos orgânicos dissolvidos que ali se concentram, enquanto dois flagelos quase transparentes flutuam frouxamente de cada lado do corpo celular, captando a luz ambiente como fios de prata que se dissolvem na escuridão cobalt do fundo infinito.

Da proa do navio de investigação, o olhar desce até à fronteira quase cirúrgica que separa o oceano aberto de um azul-índigo profundo e transparente de uma massa de água opaca, calcária, de um turquesa-branco que evoca calcário pulverizado suspenso em água glaciar — abaixo, invisíveis mas omnipresentes, dezenas de milhares de milhões de células de *Emiliania huxleyi*, cada uma com cinco a dez micrómetros de diâmetro, revestidas por placas de calcite geometricamente perfeitas que dispersam a luz solar em todas as direções com uma isotropia quase lambertiana, conferindo à água uma luminescência interior sem fonte única. A circulação de Langmuir organizou a espuma superficial em fileiras paralelas que correm com o vento e se adensam precisamente na linha de fronteira, onde as duas massas de água se atritam em lentes horizontais lentas e invisíveis. Um ganso-patola fecha as asas e mergulha em seta exatamente nessa orla produtiva, levantando um géiser de água branca-giz que capta o sol de quarenta e cinco graus antes de se desmoronar sobre a superfície turquesa-pálida, enquanto o ar traz um leve aroma sulfuroso-adocicado — dimetilsulfureto exalado pelas células de cocolitóforos sob stresse fisiológico. No horizonte, onde a floração se estende até ao limite da visão, a distinção entre oceano e céu dissolve-se num halo luminoso difuso produzido coletivamente por biliões de prismas de calcite em rotação, cada um com dois a quatro micrómetros, sedimentando lentamente para um fundo que, em milénios, se tornará calcário.

Você existe à escala de um único cocólito — cerca de três microns de largura — encravado numa matriz sedimentar que se estende em todas as direções como um continente pálido e comprimido, cada vazio preenchido por um agregado ceroso de membranas celulares e andaimes de polissacáridos esmagados por dez milhões de anos de sobrecarga geológica. À sua volta ergue-se um labirinto denso de rodas de calcite entrelaçadas, cada uma um prodígio geométrico de raios cristalinos fundidos num disco de precisão extraordinária, as suas superfícies captando a luz lateral âmbar que atravessa a coluna de sedimento como o sol da tarde filtrado por pedra fosca, lançando micro-sombras nítidas sobre cada bordo elevado e sobre o padrão de raios centrais. Ao seguir o horizonte laminar com o olhar, é possível distinguir a estratificação — bandas alternadas de assemblagens densas de cocólitos e zonas ligeiramente mais escuras onde minerais argilosos se infiltraram durante períodos glaciares de produtividade biológica reduzida, cada banda com apenas dezenas de microns de espessura, representando séculos de chuva lenta a partir da zona fótica acima. Os cocólitos mais próximos conservam as suas unidades cristalinas trigonais perfeitamente interligadas, as suas áreas centrais abertas como pequenas rosáceas no pavimento de uma catedral vistas de baixo, enquanto os mais antigos mostram os primeiros sinais de dissolução — bordos suavizados, a definição dos raios a esbater-se numa uniformidade calcária à medida que a diagénese reclama a sua geometria. Este é o pós-vida de um bloom: dez milhões de coccolitóforos comprimidos num centímetro de sedimento, a sua armadura de calcite sobrevivendo a todo o resto da sua biologia, a geometria persistindo, obsessiva e magnífica, através do tempo profundo.
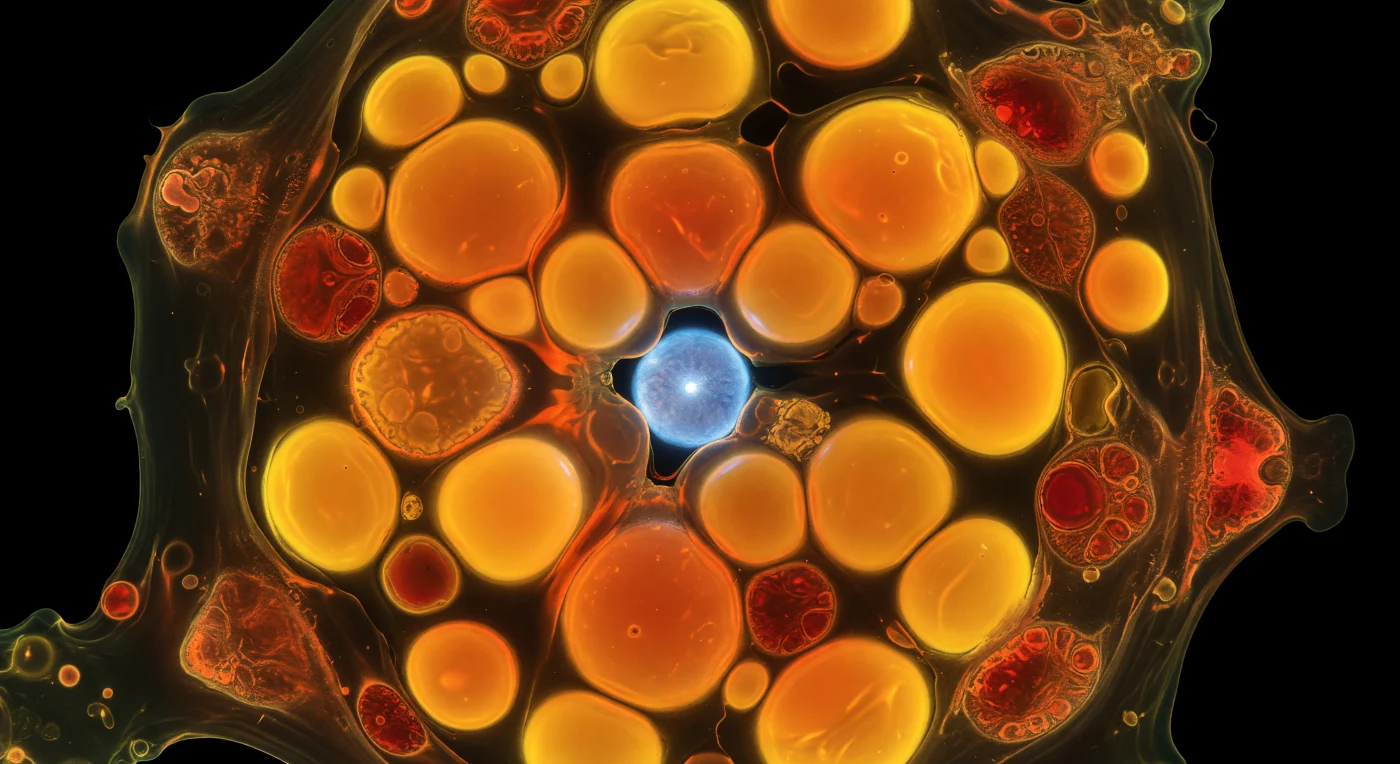
Dentro desta célula única de *Emiliania huxleyi* privada de azoto, o observador é envolvido por uma multidão de esferas perfeitas a arder em laranja-dourado intenso — gotículas lipídicas carregadas de vermelho do Nilo, cada uma delas emitindo luz de dentro para fora como lanternas de âmbar comprimidas no espaço citoplasmático. Estas reservas de lípidos neutros acumulam-se quando a célula, incapaz de sintetizar proteínas por falta de azoto, redireciona o carbono fixado pela fotossíntese para a produção de óleos de armazenamento, transformando o seu interior numa refinaria bioquímica à escala dos micrómetros. Nas margens desta paisagem incandescente, os cloroplastos foram empurrados para a periferia, a sua autofluorescência vermelha de clorofila apagando-se como brasas a arrefecer junto a um fogo maior, as membranas tilacoides ainda visíveis como laminações concêntricas mas a funcionar a meio gás, a maquinaria fotossintética desacelerada e reorientada. Em algum ponto do interior congestionado, o núcleo — marcado pelo DAPI num azul cerúleo elétrico — espreia-se através de uma fresta entre esferas de óleo, a sua superfície lisa e tensa, o nucléolo um ponto branco mais brilhante no centro, quase perdido na enchente laranja. Fora da membrana plasmática não existe nada: apenas escuridão absoluta, tornando este mundo interior completamente autorreferencial, cada fotão nascido no interior, refletido entre superfícies oleosas, reabsorvido e reemitido, até que a célula inteira arde com uma luz que não tem origem nem saída exterior.

As mãos movem-se pela água e o oceano responde com fogo frio: cada dedo deixa um rasto de azul elétrico a 460 nanómetros que persiste dois segundos inteiros antes de se dissolver em teal e depois em nada, o turbilhão de cada vórtice mapeado com precisão em luz viva produzida por milhões de células de *Lingulodinium polyedra* que disparam os seus escintilosomas em cascata mecânica, luciferase encontrando luciferina numa reação bioquímica mais antiga do que qualquer animal. A crista de cada vaga transporta um pulso sustentado em vez de um clarão, porque o movimento orbital das partículas de água agita continuamente as células à superfície, revelando de repente a geometria oculta do mar aberto numa topografia escrita em luz — e cinquenta metros mais longe, a esteira de um barco desaparecido forma um rio de incandescência azul-branca tão denso que parece quase sólido, uma estrada luminosa traçada através da água negra por um casco que já não existe no campo de visão. O corpo do observador define o seu próprio limite: onde o torso rompe a superfície, um halo de luminescência de dois comprimentos de braço mapeia cada micro-ondulação gerada pela respiração, cada batimento do coração transformado em nova cascata de células sensíveis, de modo que simplesmente existir na água é suficiente para produzir uma carta viva dos menores movimentos do corpo. Abaixo da superfície, o brilho penetra meio metro antes de ser absorvido pela coluna de água, criando um volume luminoso pouco profundo e infinito em simultâneo — a pele iluminada apenas por esta luz surge azul-acinzentada, como se mergulhada em algo diferente de água do mar, enquanto o céu negro acima e o oceano negro abaixo se tornam indistinguíveis, e só a fronteira entre ambos, marcada em fogo frio, confirma que existe um mundo.
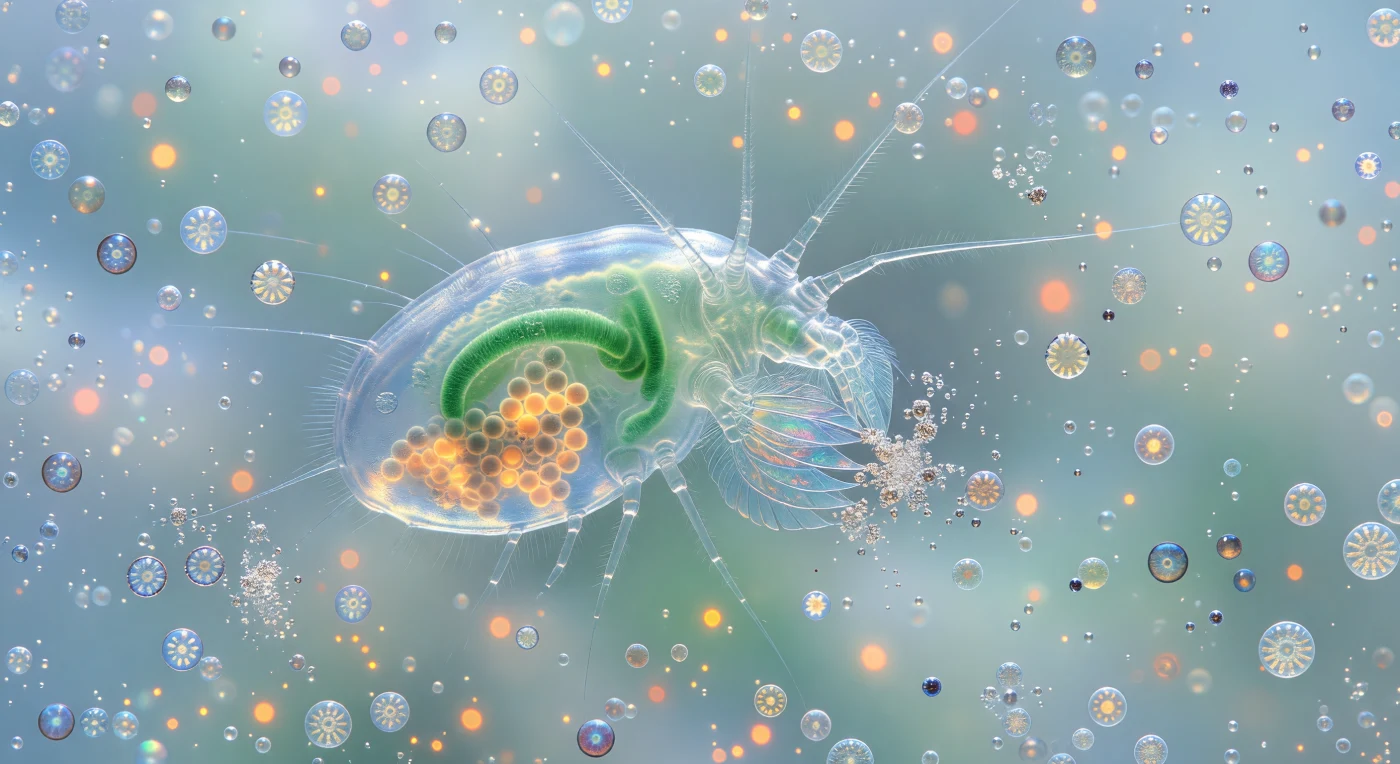
Você paira a dois milímetros de distância de um ser cujo corpo é uma janela viva: o copépode *Calanus* atravessa a coluna de água num azul-turquesa denso de cocólitos soltos, dez milhões por mililitro, que dispersam a luz filtrada do oceano numa névoa luminosa que dissolve qualquer horizonte. O corpo translúcido do crustáceo age como uma lanterna de vidro fosco — o tubo intestinal cor de jade e os agrupamentos de ovos âmbar-alaranjados ardem através de três camadas de cutícula quitinosa, órgãos iluminados de dentro para fora pela luz difusa que desce da superfície. As maxilípedes, dispostas como penas sobrepostas de vidro soprado, vibram no limite do campo visual e cada seta capta células individuais de cocolitóforos tal como uma teia de aranha apanha gotas de orvalho; no momento do contacto, as esferetas calcificadas explodem em rajadas de purpurina branca, cada coccosfera destruída libertando uma pulverização de discos de calcite de dois micrómetros que giram e cintilam antes de serem varridos pela corrente alimentar. Ao longo dos arcos lentos e quase imperceptíveis do vórtice, coccosferas intactas de *Emiliania huxleyi* giram enquanto derivam em direção à boca do copépode, as suas placas de calcite entrelaçadas refletindo prata, ouro pálido e azul-branco frio consoante o ângulo de iluminação, os cloroplastos no interior emitindo uma autofluorescência vermelho-alaranjada que aquece a névoa turquesa como pequenas constelações de calor metabólico.

Suspenso na coluna oceânica a esta escala diminuta, o observador encontra-se face a face com dois organismos separados por apenas alguns comprimentos de célula: à esquerda, a célula diploide em fase C da *Emiliania huxleyi* ergue-se como um pequeno lustre mineral, o seu cocosférico inteiramente revestido de placas de calcite creme-branco cujos esporos radiais e fenestras centrais captam a luz difusa que filtra lá de cima e a devolvem em clarões prismáticos frios, dourados e brancos; à direita, a sua gémea haploide em fase N apresenta uma nudez quase vulnerável — membrana translúcida de verde-creme, ligeiramente irregular, deixando adivinhar o calor âmbar-jade dos cloroplastos no interior. Os dois flagelos desta célula menor estão congelados no auge do batimento, curvados em sinusoides graciosíssimas contra o azul profundo, enquanto o haptonema se enrola discretamente entre eles como um resorte de vidro molhado quase invisível. Esta dualidade morfológica não é acidental: representa a alternância de gerações que define o ciclo de vida da espécie, sendo a fase C a responsável pela precipitação de carbonato de cálcio que, ao afundar após a morte celular, transporta carbono para o fundo oceânico há dezenas de milhões de anos, sedimentando lentamente nas faléisas de giz do mundo inteiro. Ao fundo, cocólitos soltos derivam como faíscas brancas a distâncias variadas, criando uma profundidade de campo que amplifica a vertigem desta escala, onde o Movimento Browniano é lei e a imobilidade uma negociação constante com o fluido viscoso que governa tudo.

Erguendo o olhar desde a base das Falésias Brancas de Dover, o observador contempla uma parede quase vertical de calcário cretáceo que se eleva a mais de cem metros, ardendo de brancura sob um sol rasante que faz a superfície parecer emitir luz própria contra o céu cinzento-azulado do Atlântico Norte. O que parece uma rocha uniforme é na verdade uma acumulação densa de bilhões de cocolitos — minúsculas placas de calcite segregadas por algas unicelulares durante o Cretáceo tardio, há 65 a 100 milhões de anos, quando um mar quente e raso cobria esta região e florações ciclicamente explodiam e morriam, chovendo lentamente sobre o fundo em camadas de lama biogénica que a compressão geológica transformou em carbonato sólido. As bandas escuras de sílex que cortam a face branca em linhas estratigráficas quase perfeitas são fantasmas químicos dessas antigas profundezas — sílica remobilizada durante a diagénese que migrou ao longo de planos preferenciais, registando em matéria dura os ritmos de sedimentação de oceanos desaparecidos. A meio da falésia, quase invisível mas inconfundivelmente vivo, um fulmar pousado numa cornija de calcário empresta escala humana a este monumento mineral, tornando súbita e vertiginosamente real a altura desta enciclopédia de organismos invisíveis convertidos em pedra.