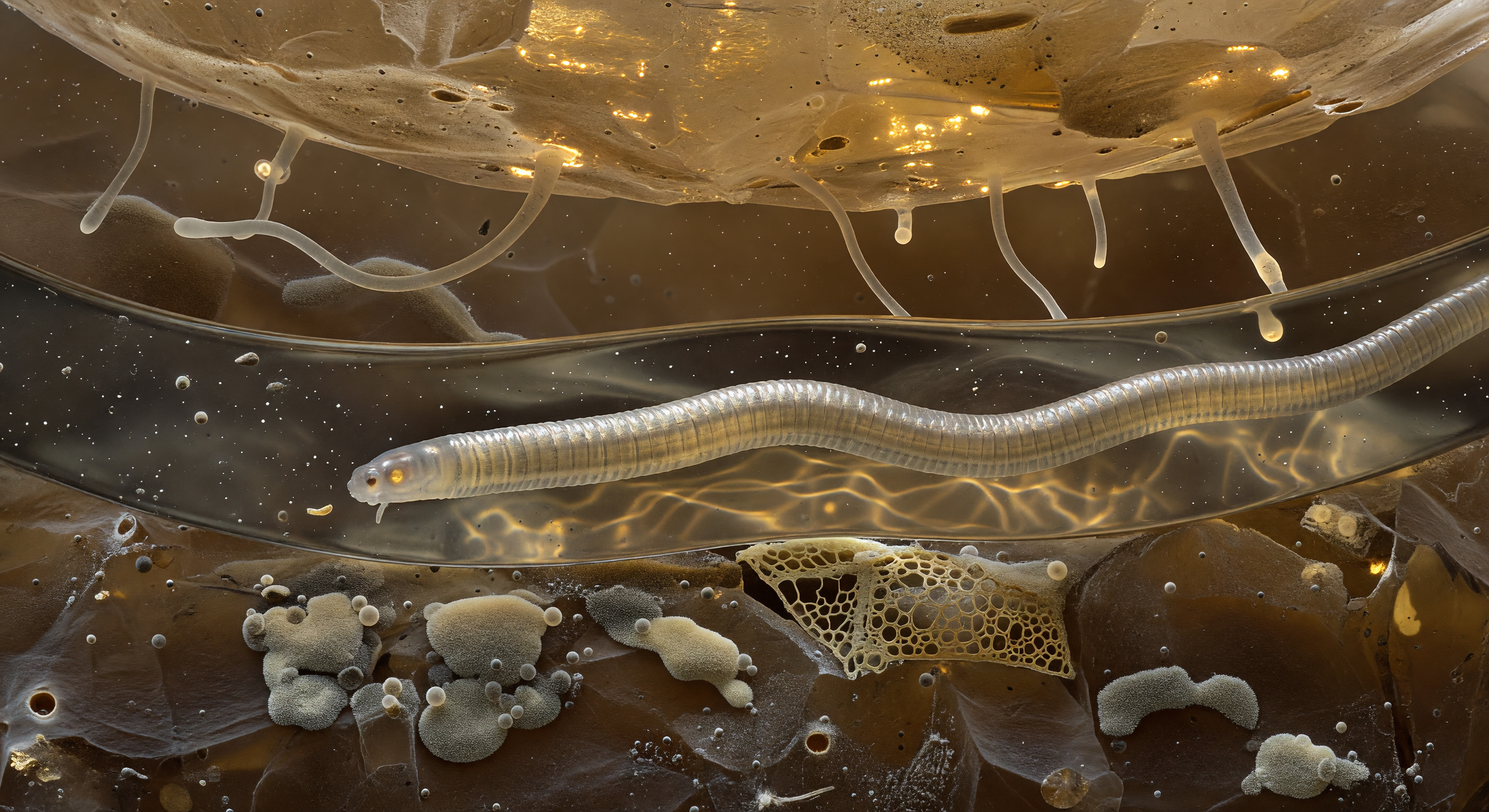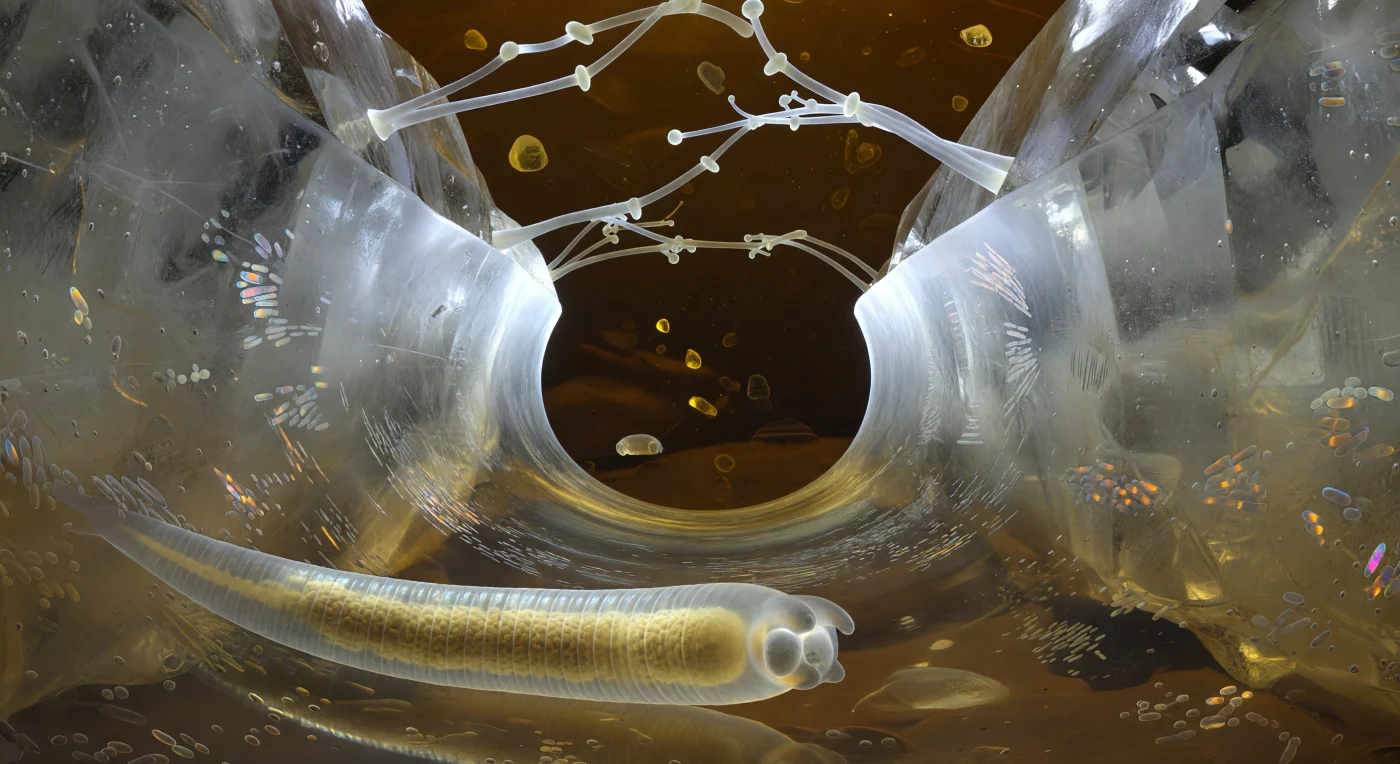
Der Blick führt durch einen kapillaren Wassertunnel, der von zwei massiven Quarzblöcken geformt wird — ihre Oberflächen ragen wie transluzente Klippen aus wasserklarem Kristall auf, durchzogen von blassen Mineraleinschlüssen, die das gestreute Licht in prismatische Splitter brechen. Die Tunnel-Geometrie ist vollständig von der Oberflächenspannung bestimmt: An beiden Enden wölben sich die Menisken wie blendend weiße parabolische Spiegel nach außen, ihre Krümmungskräfte stark genug, um das Wasser als zusammenhängende Linse zu halten und den Korridor mit einem fast chirurgischen Glanz zu fluten — während die Wände selbst von lebenden Bakterienbiofilmen überzogen sind, die in Bronze, Violett und Aquamarin schimmern, und darüber transluzente Pilzhyphen wie Hängebrückenkabel von Quarz zu Quarz spannen. Im Vordergrund liegt der eigene Körper als gebogener, durchscheinender Zylinder auf dem Wasserfilm, seine Kutikula-Annulierungen fangen das laterale Irisierlicht in kurzen Silberbögen, und durch die Körperwand hindurch leuchtet der Darm warm und bernsteinfarben — ein Kern aus gelb-braunen Granula in den Intestinalzellen, deren Autofluoreszenz sich durch das umgebende Wasser streut und den unteren Korridor in ein gedämpftes Gold taucht. Jenseits der weißglühenden Meniskusgrenzen löst sich die Welt in tiefes Ockerdunkel auf — die eigentliche Bodenfinsternis — und macht begreifbar, dass dieser Korridor aus Wasser, Mineral und Biomasse ein vollständiges Universum ist, in dem Schwerkraft bedeutungslos ist und jede Bewegung im viskosen Medium anhaltende Muskelarbeit erfordert.

Im Innern des terminalen Pharyngealbulbus hängt man wie in einer gewaltigen Kathedrale, deren geschwungene Wände sich ringsum zu dem dreiarmigen Lumen zusammenziehen — einem sternförmigen, im Querschnitt Y-förmigen Kanal, der von konzentrierter Pharyngealflüssigkeit honigfarben durchleuchtet wird. Drei keilartige Mahlplatten aus elektronendichter Kutikula — hornfarben, an ihren abgeschrägten Kanten mit schwachen Interferenzstreifen wie poliertes Schildpatt — schlagen unter hydraulischem Druck radial gegeneinander: jeder Schlag dauert eine Bruchteilsekunde und verdrängt die viskose Flüssigkeit in trägen, glasartig wogenden Bögen nach außen, während zertrümmerte Bakterienfragmente und goldene Lipidtröpfchen in langsamer Brownscher Drift um einen herumschweben. Die Muskelwände des Bulbus sind streng geordnete Architektur — Sarkomerbänder verlaufen in blassrosa, nahezu transparentem Zytoplasma wie die Rippen eines romanischen Gewölbes, mit einem feinen blausilbernen Schimmer dort, wo Aktin- und Myosinfilamente in exakter Registeranordnung das Licht brechen. Das Licht selbst hat keine Quelle: Es streut diffus durch die transluzente Kutikula und das Muskelgewebe herein und taucht den gesamten Innenraum in ein gleichmäßiges, warmes Bernsteinleuchten, das den zähflüssigen, proteindurchwirkten Inhalt dieses mikroskopischen Zermalmungsapparats wie in flüssigem Harz konserviert erscheinen lässt.

Der Blick richtet sich von unterhalb eines massiven Quarzkorns nach oben, dessen muschelförmig gebrochene Flächen das diffuse Licht in kalte blau-weiße Aufblitzungen zersplittern – ein Gebirge aus glasklarem Mineral, das zwischen Kliffkante und Kathedralpfeiler changiert. Auf dem Scheitelpunkt dieses kristallinen Kolosses balanciert eine Dauer-Larve auf ihrer Schwanzspitze: ein bernsteinfarbener Turm aus verdickter Kutikula, deren feine Ringanulierungen im Streiflicht wie zarte topografische Wellen hervortreten. Durch die halbtransparente Körperwand hindurch schweben cremeweisse Fetttröpfchen wie eingefrorene Schneeflocken im Bernstein – gespeicherte Energiereserven für eine ungewisse Zukunft, denn das Dauer-Stadium ist ein biochemisch versiegelter Überlebenszustand, metabolisch heruntergefahren, die Mundöffnung durch einen dichten Buccalpropf verschlossen. Tief unter dem Quarzgipfel verliert sich die Welt in warmem Ocker und Umber: Mineralsplitter, Bodenaggreggate und blass-weiße Pilzhyphen spannen sich wie verwitterte Hängebrücken zwischen fernen Partikelwelten, während ein feiner Schleier aus Exopolymeren und gelöstem organischen Material die gesamte Szene in eine leicht komprimierte Tiefenschärfe taucht, die den senkrechten Stand der Larve gleichzeitig schwindelerregend und unverrückbar erscheinen lässt.
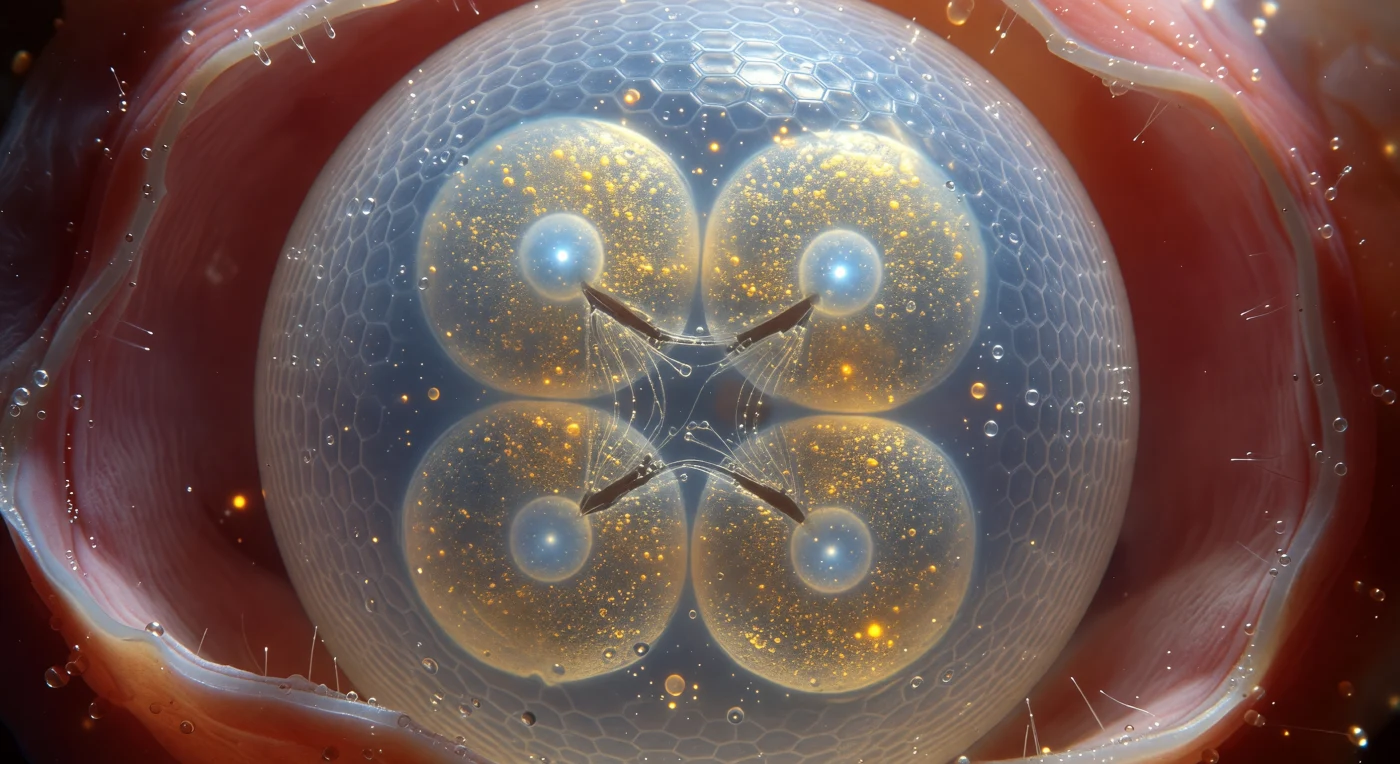
Du schwebst so nah an der Eischale, dass ihre gewölbte Oberfläche deinen gesamten Blick ausfüllt wie die Flanke eines Eisbergs: eine milchig-transluzente Chitinmembran, deren verflochtene Polysaccharidfasern einfallendes Licht in ein kühles Blau-Weiß zerstreuen und unter polarisiertem Licht verschiebende Indigo- und Silberbänder über ihre Oberfläche wandern lassen. Dahinter pressen sich vier Blastomere mit geometrischer Präzision gegeneinander – ihre abgeflachten Kontaktflächen bilden dunkle Furchenlinien wie zwischen gefangenen Seifenblasen –, jede Zelle prall gefüllt mit goldgelben Dottergranula, die ein warmes Bernstein-Orange durch die Schale bluten lassen. Im Zentrum jeder Blastomere hängt ein arktisch-blauer Zellkern, sein Nukleolus leuchtend wie ein kleiner kalter Stern, während zwischen zwei Zellen die Geistergestalt einer mitotischen Spindel noch sichtbar ist: silberweiße Mikrotubuli, straff wie Drähte zwischen Chromosomenmassen gespannt, bevor sie sich endgültig auflösen. Das periviltelline Fluid, das Ei und umgebende Uteruswand trennt, ist von makelloser optischer Klarheit – ein biologisches Immersionsmedium, das jeden inneren Detail in scharfer Schärfe hält, während das rosafarbene, von Mikrovilli besetzte Gewebe dahinter in warmes Licht verschwimmt. Dieses versiegelte, druckbeaufschlagte Universum von kaum einem Zehntel Millimeter ist vollständig autark: ein Entwicklungsprogramm, das in weniger als einer Stunde eine weitere Zellteilung vollziehen wird.

Die Welt, die sich vor dir ausbreitet, ist eine endlose Ebene aus geriffeltem Gold und Schiefer-Grau: perfekt parallele Annulationsrücken ziehen sich Horizont zu Horizont wie versteinerte Dünen, ihre abgerundeten Kämme von einem harten, schräg einfallenden Licht erfasst, das jeden Talboden in absoluten Schatten taucht. Diese Strukturen sind keine passiven Oberflächenornamente — sie sind biomechanische Notwendigkeit, das Ergebnis einer mehrschichtigen Kutikula aus vernetztem Kollagen und Cuticlinen, die unter dem Innendruck des hydrostatischen Körperzylinders unter ständiger Spannung steht und dem Nematoden seine charakteristische Schlängelbewegung erst ermöglicht. Links erhebt sich die laterale Ala wie ein kontinuierliches Gebirgsmassiv über die annulierten Ebenen — eine longitudinale Strukturfalte der Hypodermis, deren flachere Transversalstreifung im Streiflicht anders glänzt und die vermutlich sensorischen oder adhäsiven Funktionen dient. Aus dem vorderen Horizont ragen kuppelförmige Papillae der Kopfsensilla wie monolithische Halbkugeln hervor, jede mit einem einzigen zentralen Mikroporus an der Spitze — Eintrittspforten mechanosensorischer und chemosensorischer Nervenendigungen, die diesem Tier sein vollständiges Bild der Welt liefern. Die gesamte Oberfläche ist unbeweglich und zugleich lebendig gespannt, ein unter Druck stehendes, atmendes Material, das in seiner goldgrauen Stille eher an Mineral als an Fleisch erinnert.

Wer hier durch die Welt gleitet, befindet sich an der Spitze eines Fadenwurms, die Körperachse dicht über dem Oberfläche eines Nähragar-Gels gedrückt, das in alle Richtungen bis an die Grenzen der Wahrnehmung reicht — ein weich gewelltes, transluzentes Terrain, von unten durch intensives kaltweiß-es Durchlicht erleuchtet, das die gesamte Umgebung in ein leuchtendes Diorama verwandelt. Direkt voraus erhebt sich der bakterielle Rasen wie ein undurchdringlicher Urwald: dicht gedrängte *E. coli*-Stäbchen als dunkle, leicht irisierende Silhouetten gegen das Gegenlicht, Schulter an Schulter zu einem fast schwarzen Blätterdach aus lebender Biomasse verschmolzen, das sich in jede laterale Richtung bis zum Horizont erstreckt. Hinter dem Wurm öffnet sich im Kontrast dazu die sinusförmige Fraßspur wie ein Kathedralen-Schiff aus geräumtem Gel, die Wände des Korridors aus abgescherten Bakterienzellen glänzen im Durchlicht mit einem bläulich-weißen Rand, während ältere Spuren weiter hinten bereits wieder von neu einwandernden Zellen bewölkt werden. Im unmittelbaren Vorderfeld arbeitet der Pharynx als hochfrequente Pumpe: der Terminalbulbus kontrahiert und entspannt sich in rascher Folge — rund 250 Mal pro Minute — und zieht dabei einzelne Bakterienstäbchen in den Mahlapparat, während sich der Darm dahinter fortschreitend orange-braun färbt, die Darmgranula als Punkte fainter Autofluoreszenz aufblitzen und die gesamte Verdauungsachse als leuchtende Röhre unter der korrugiert-metallisch schimmernden Kutikula sichtbar wird.
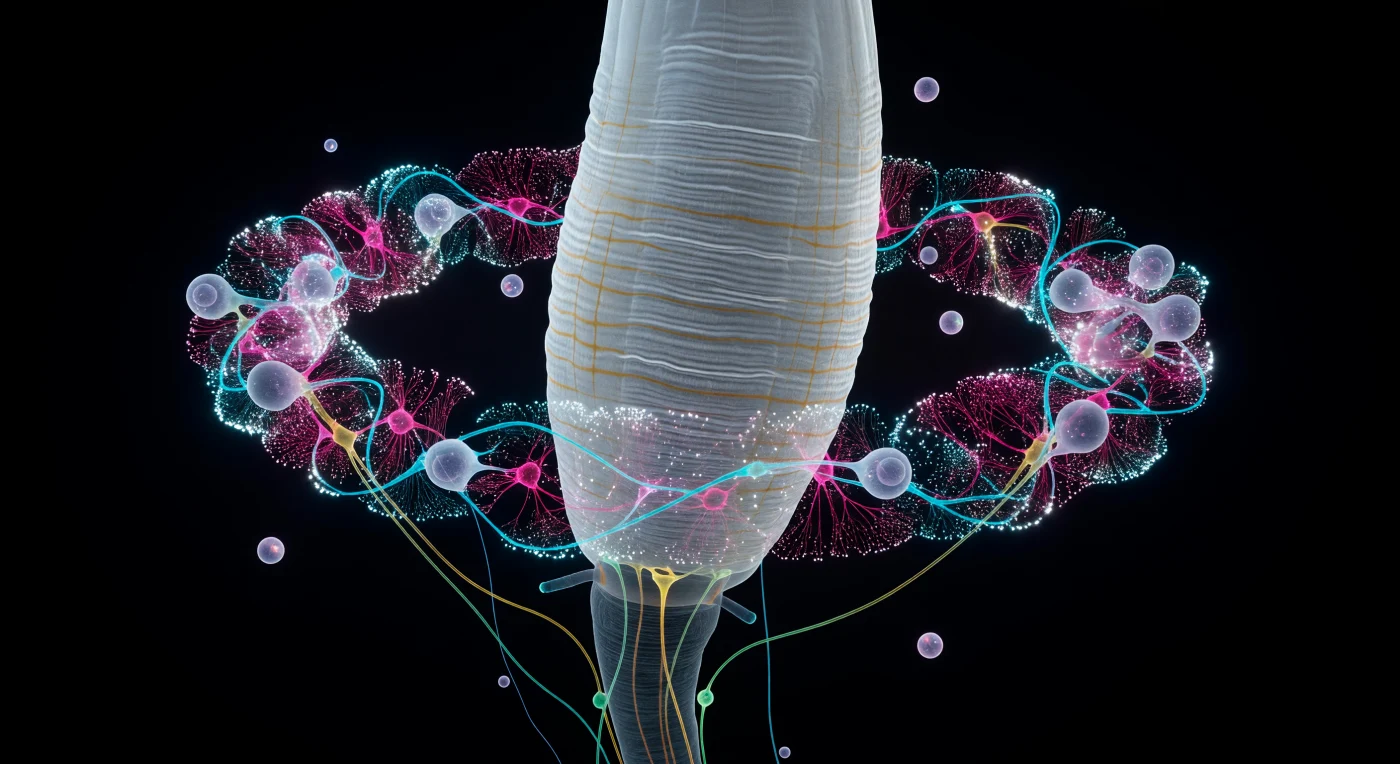
Im Inneren des Nervensystems eines der häufigsten Tiere der Erde schwebt der Betrachter vor dem bleich gestreiften Pharynxpfeiler, dessen Längsrillen und Querrunzeln das Licht des Nervenrings von allen Seiten einfangen – einer kolossalen Säule aus Streifen muskel, die wie eine Gletscherflanke aus dem pseudocoeloem Dunkel emporsteigt. Um diesen zentralen Anker herum lodert der circumösophageale Nervenring, ein kaum zwanzig Mikrometer breiter Kranz aus Neuropil, in dem sich Zehntausende von synaptischen Kontakten in einem Volumen verdichten, das man in einem einzigen langsamen Gleiten durchqueren könnte: kühlcyane Axonbündel amphider Sinnesneuronen weben sich von außen nach innen, heißmagentafarbene Interneuron fasern pulsieren darunter, und gelbgrüne Motoneuronkommissuren schwingen bogenförmig in die Schwärze des Bauchstrangs hinab. Synapsencluster blitzen als winzige weiße Supernovae durch das Geflecht, Ausdruck eines elektro chemischen Feuers, das keine externe Lichtquelle benötigt, weil jede Neuronenklasse ihre eigene Spektral signatur trägt wie ein Mineral seine Kristallfarbe. Diese lebendige Krone ist das gesamte Zentralnervensystem eines Tieres, dessen vollständige Verschaltung – 302 Neuronen, rund 7000 Synapsen – als einziges Mehrzeller-Konnektom vollständig kartiert ist, ein Kontinent neuronaler Komplexität, der in permanenter Dunkelheit brennt, während der Wurm darüber mit einem Körper länge pro Sekunde seinen bakteriellen Kontinent durchpflügt.
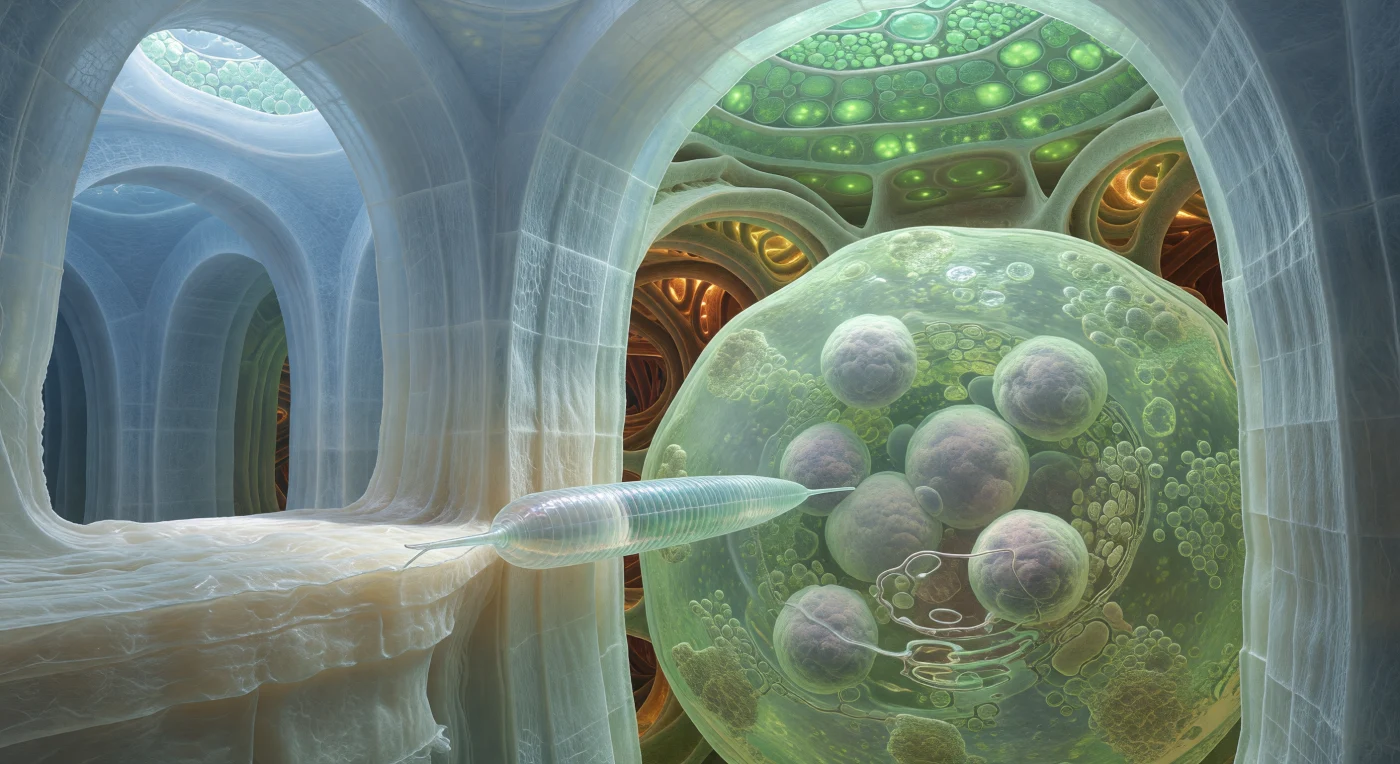
Wir treiben reglos an der Kreuzung zweier gewaltiger Zellwände, eingeklemmt zwischen gewölbten Platten aus blassblau-weißem Zellstoff, die wie überflutete Kathedralsäulen um uns aufragen – jede Lage ein komprimiertes Geflecht aus Cellulosemikrofibrillen, deren Oberfläche ein feines Gittermuster trägt, das das kühle Jadegrün des von weit oben hereinfilternden Lichts einfängt und bricht. Dicht vor uns presst ein Nematodenjuveniler seinen nadelfeinen Stilett gegen diesen Wandknoten: ein straffes, glasklares Druckrohr aus annulierter Kutikula, das in der schimmernden Diffuslicht in einem schwachen Irisierschein aufleuchtet – das gesamte Tier ein biologischer Hydraulikkolben, der gezielte Kraft auf eine einzige mikroskopische Schwachstelle konzentriert, um sie zur Riesenzelle zu formen. Dahinter öffnet sich jene transformierte Wirtszelle wie ein weites, milchig-grünes Hallenschiff: ihr Zytoplasma so dicht mit Ribosomen durchsetzt, dass es wie von innen beleuchtetes Mattglas wirkt, während mehrere riesige Zellkerne – blasslavendelgrau, ihre Kernhüllen leicht gefaltet – darin treiben wie aufgedunsene Monde in einer stummen Umlaufbahn. Im Tiefenfeld glühen spiralverdichtete Xylемgefäße in warmem Bernstein-Rostbraun, als pulsierte durch ihre lignifizierten Wände ein uralter hydraulischer Rhythmus, und das gesamte Szenario ist in eine apoplastische Stille getaucht, in der jede Oberfläche mit dem feinen Glanz wässriger Filme überzogen ist und kein Schatten hart fällt.
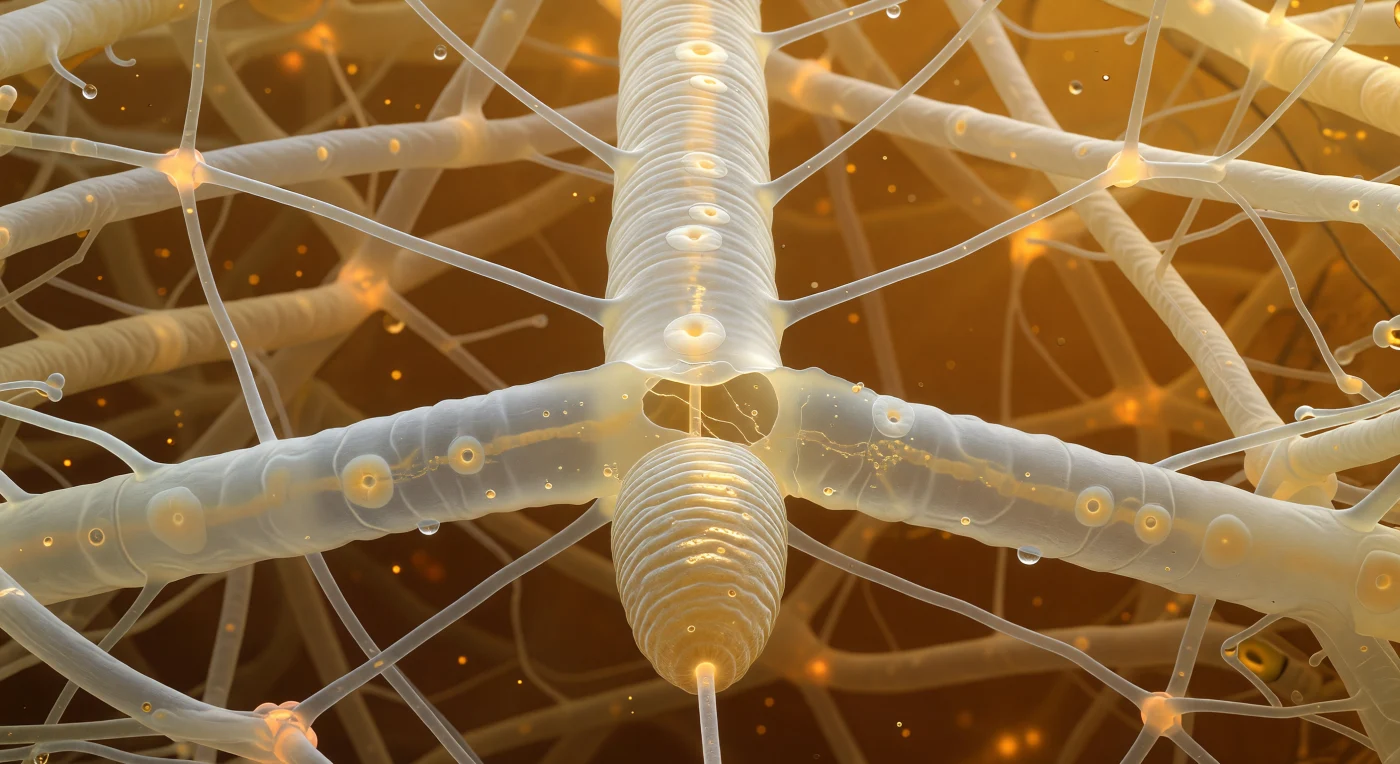
Im Inneren des Bodens hängt man in einem Geflecht lebender Fäden: Vor einem dehnt sich eine mächtige, weiß-transluzente Hyphe aus wie ein unter Druck stehendes Rohr, durch das in bernsteinfarbenem Strom Organellen und Zytoplasma fließen, die Septen als leicht opake Scheiben mit zentraler Pore sichtbar, das ganze Innere von einem warmen Tee-Gold durchleuchtet, das durch verrottendes Laub von oben hereinfiltriert. Unmittelbar davor — den Vordergrund ausfüllend — drängt sich das Vorderende eines fungivoren Nematoden gegen die Hyphen-Wand: ein gerippter, leicht kegelförmiger Zylinder aus Kutikula, dessen Stylet bereits durch die Hyphenwan gestochen hat, der Pharyngeal-Pumpe rhythmisch folgend, während rund um den Einstich eine Retrakt-Welle des Zytoplasmas sichtbar nach außen weicht und winzige Tröpfchen extradierten Materials das Bernsteinlicht in warme Spiegelreflexe zersplittern. Das Nematod bewegt sich in einem Regime verschwindender Trägheit — an Stelle von Schwerkraft regieren Oberflächenspannung und Viskosität jede Regung — und so klebt ein feiner Wasserfilm zwischen Zellulosefasern und Hyphenwänden als helle, gebogene Meniskus-Linien an allen Oberflächen, die gesamte architektronische Tiefe des Bildes aus raumem Bernstein, durchscheinendem Elfenbein und dem einzigen kühlen, metallischen Reflex des Stylets aufgebaut.

Das Sichtfeld öffnet sich auf einen Korridor aus riesigen, cremefarbenen Sandkörnern, deren von Meeresströmungen polierte Siliziumdioxidflächen wie gewölbte Mondlandschaften abfallen, während zwischen ihnen goldbraune Diatomeenschalen aus biogenem Opal eingekeilt sind — durchbrochene Glaskathedralen, deren Punktreihen und Striae prismatische Aureolen aus bernsteinfarbenem Licht ins blaue Halbdunkel streuen. Die Oberflächen der Körner sind von einer Schwefelbakterienmatte überzogen, die in Magenta und Pflaume schimmert und bei jeder Blickwinkeländerung langsam chromatisch aufzuglühen scheint, ein strukturfarbenes Membransystem aus gestapelten Bakterienzellschichten. Durch diesen Zwischenraum schiebt sich ein Meeresnematode — ein Tier von 0,3 bis 1,5 mm Länge, das sich durch sinusförmige Muskelwellen vorwärtsbewegt, im Niederreynolds-Regime, wo Trägheit bedeutungslos ist und das umgebende Meerwasser sich wie ein zähes Gel anfühlt — sein gerippter Chitinpanzer besetzt mit adhärierenden Diatomeenfragmenten und feinen Sedimentpartikeln wie eingelassenes Mosaik. Im rechten Hintergrund überragt ein cremefarbenes Foraminiferen-Gehäuse die Szene wie ein gotisches Gewölbe aus Kalzit, seine gekammerten Bögen in kühle Innenfinsternis verlaufend, während goldene Borsten eines Polychaeten von oben zwischen den Körnern herabragen und in blaugrauem Kolloidaldunst verschwinden.
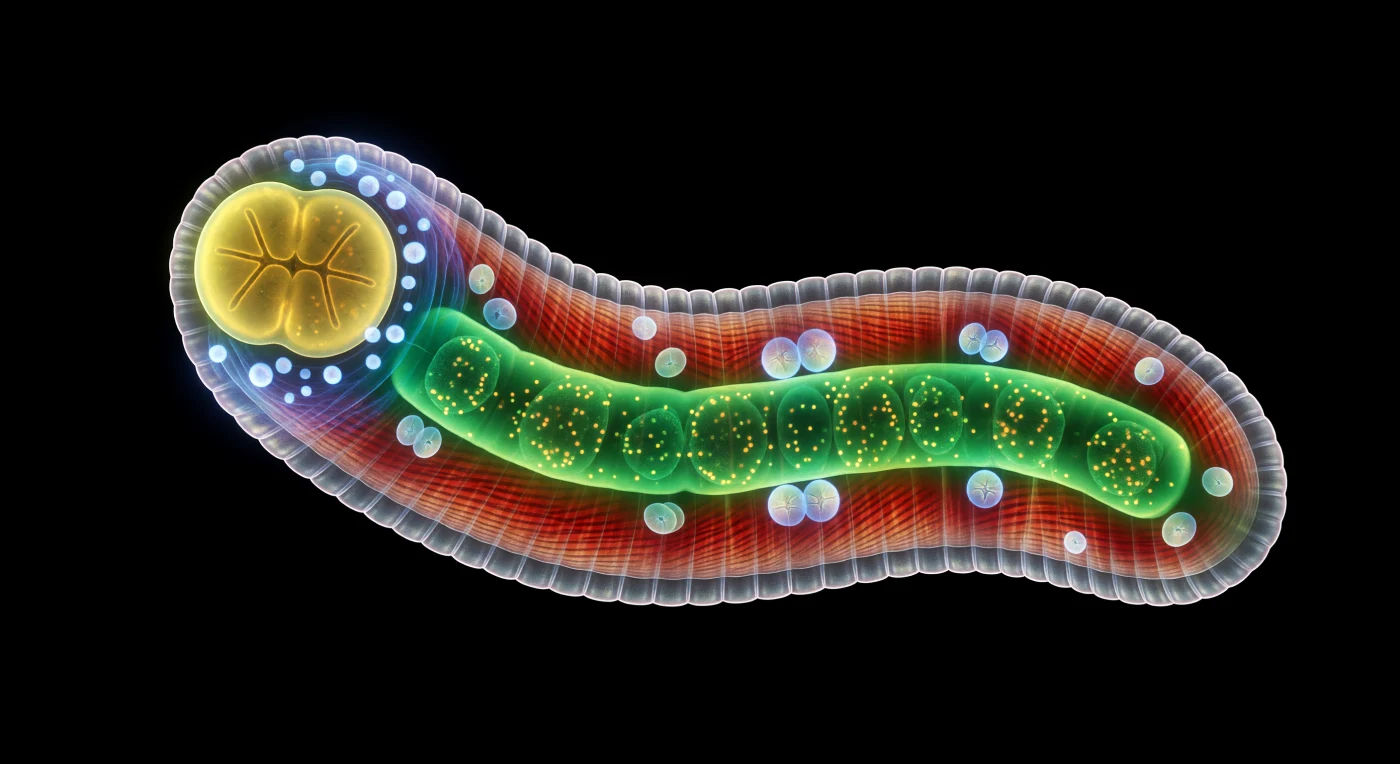
Vor Ihnen wölbt sich eine gewaltige, selbstleuchtende Gestalt in der charakteristischen S-Kurve eines adulten Hermaphroditen – ein lebendiger Schaltkreis aus überlagerten Farben, der sich wie eine nächtliche Stadtsilhouette aus absolutem Schwarz erhebt, jedes Photon ausschließlich aus dem Inneren des Körpers selbst stammend. Die äußere Kutikula schimmert perlweiß mit einem Hauch von Violett, wo ihre kollagenen Schichten das innere Licht brechen, während sich darunter vier mächtige Längsbänder der Körperwandmuskulatur in tiefem Karmesinrot erstrecken, deren diagonale Sarkomerstreifen wie die Hitzeschilde einer Raumsonde das Licht moiréartig modulieren. Der Darm zieht als sattgrüner, leuchtender Schlauch durch die gesamte Körperlänge, seine riesigen Epithelzellen prall gefüllt mit hunderten gelbgrüner Granula, die in einem langsamen, viskosen Strom durch das Zytoplasma treiben und das emeraldene Licht als einen Farbschein auf die benachbarten roten Muskelpartien werfen. Um den zweilappigen, citringelben Pharynx im Vorderkörper liegt wie eine Krone aus Saphiren der Nervenring – einzelne cyanblaue Neuronenzellkörper und ein kühler zeruleanfarbener Neuropilschleier, das einzige neuronale Koordinationszentrum eines Tieres mit exakt 302 Nervenzellen. Im Uterus dahinter treiben blass-opaleszierende Embryonen wie Miniaturjuwelen, ihre Furchungsstadien als blauweiße Kugelgeometrien sichtbar, jede Zellteilung ein Takt in einem Entwicklungsprogramm, das seit Jahrmillionen nahezu unverändert abläuft.

Vor dir öffnet sich ein Abgrund aus lebendem Bernstein — die Mundöhle eines räuberischen *Mononchus*, deren geschwungener Dorsalzahn wie ein monolithischer Bogen aus sklerotisiertem Kutikulargewebe in warmem Kastanienbraun und dunklem Ocker aus dem Dunkel herausragt, seine polierte Oberfläche in schimmernden Karamelltönen schillernd durch das diffus gestreute Bodenlicht. Die Innenwände der Buccalhöhle sind mit radial angeordneten Dentikeln ausgekleidet — elfenbeinfarben, nadelspitz, in konzentrischen Ringen in die pulsende Dunkelheit des pharyngealen Schlunds zurückweichend —, während der gesamte Raum wie ein gotisches Gewölbe aus Horn wirkt, dessen feine kutikullare Annulierungen das Licht in irisierenden Bändern brechen. Gegen die Kammerwand gepresst kollabiert ein kleinerer bakterivorer Nematode: Seine Kutikula — normalerweise ein unter hydraulischem Druck straff gespannter, perlgrau-transparenter Zylinder — faltet und kräuselt sich nun unter dem Sog des Räubers, diagonale Druckfalten propagieren von der Einzugsstelle zurück entlang des Körpers wie ein weicher Schlauch, der ausgequetscht wird. Durch die noch immer transparente, komprimierende Hülle hindurch sind die inneren Organe des Beutetiers sichtbar: der bilobäre Pharynx als dunklere Masse im Vorderkörper, der sinuöse Gonad als leuchtend-heller Gewebestrang unter Druck, und der Darm, dessen Zellen mit autofluoreszierenden Granula gefüllt ein letztes goldgelb-grünes Leuchten aus dem Körperinneren senden — ein erlöschendes Biolumineszenz-Glimmen im Angesicht der verschlingenden Schwärze.
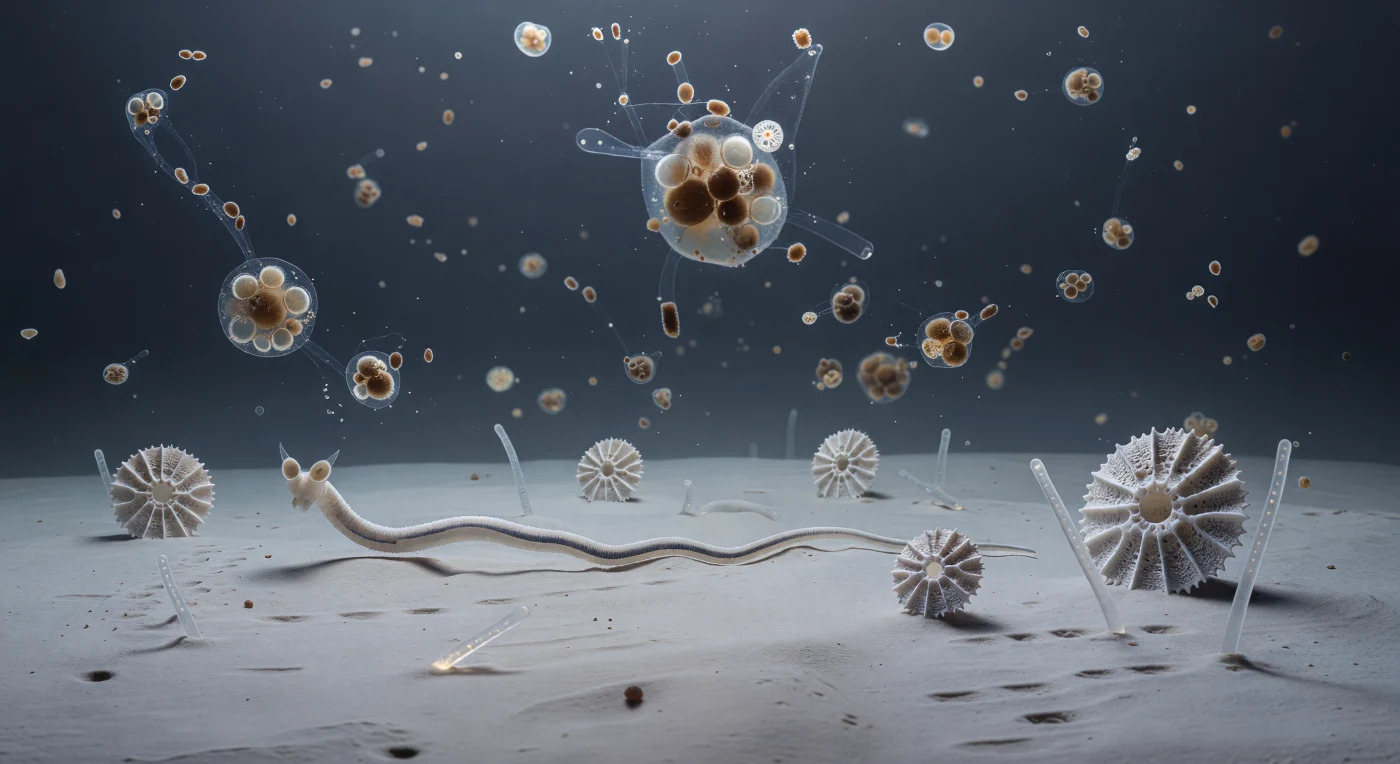
Die Welt, die sich hier auffaltet, besitzt keine Grenze, keinen Horizont, keine erkennbare Tiefe — nur eine schier endlose Fläche aus bleichem, knochenweißem Tiefseeton, dessen Textur so gleichförmig und dicht ist, dass die Oberfläche unter dem Körpergewicht eines Fadenwurms kaum nachgibt, als wäre der Meeresgrund aus erstarrtem Atem geformt. Foraminiferen-Gehäuse — geometrisch filigrane Kalzitkammern, jede in Relation zu den hier lebenden Nematoden so groß wie ein verlassener Dom — ragen vereinzelt aus dieser mattergrauen Ebene, ihre Poren von anoxischem Bodenwasser durchdrungen, während Kieselsäure-Spicula von Schwämmen wie gestürzte Sendemasten quer über den Sedimentgrund liegen und das diffuse Umgebungslicht in zarte prismatische Reflexe brechen. Von irgendwo weit oberhalb — in einer Wassersäule, die sich über zwei Kilometer bis zur lichtdurchfluteten Oberfläche erstreckt — sinken marine Schneeflocken mit der Langsamkeit von Objekten, die durch ein Medium fallen, das kaum weniger dicht ist als sie selbst: jede Aggregation ein loses Gefüge aus leeren Diatomeen-Frustreln, verdichteten Kotpillen, Schleimstofffäden und mineralischem Staub, braun-amber leuchtend gegen das kalte Blauschwarz des Freiwassers. Dazwischen bewegen sich die Nematoden in sinusförmigen Wellenbewegungen, deren einzelne Körperwellen so langsam ablaufen, dass ihre Fortbewegung erst nach Sekunden erkennbar wird — ihre überproportional großen Amphid-Organe vorwärts gerichtet, chemische Signale aus den niedersinkenden Aggregaten abtastend, während der Tiefseeschlick hinter ihnen die kaum einen Körperdurchmesser tiefen Spurrinnen sofort wieder mit nasser Talkum-Feinheit auffüllt.

Innerhalb einer haarfeinen Wasserbrücke zwischen zwei Bodenpartikeln hängt der Blick reglos im Bernstein: Über uns wölbt sich die Meniskusoberfläche wie ein konvexes Fischauge aus reinem Oberflächenspannungsglas, darin das gesamte Bodengefüge zu einer einzigen verzerrten Panoramakugel zusammengepresst – Quarzflächen, rostbraune Feldspatschichten, dunkle organische Filamente, alles kreisförmig in die spiegelnde Kuppel gefaltet und von jedem kleinsten Brownschen Zittern der schwebenden Kaolinitplättchen zum Flimmern gebracht. Das Wasser selbst ist kein neutrales Medium, sondern ein warmes Bernstaunlösungsgemisch aus Huminsäuren, die aus zerfallendem Blattmaterial gelöst wurden, und gibt dem Fluid eine fast biolumineszente Tiefe, während Tonteilchen von kaum einigen hundert Nanometern Breite in trägen, erratischen Bögen taumeln und dabei silbrig, dann dunkel, dann rostgold aufleuchten. Wo der eigene Kutikularmantel des Nematoden am Bildrand an den Meniskus stößt, brennt die Dreiphasenkontaktlinie als irisierender weißgoldener Saum – eine konzentrierte Brechungszone, an der Wasser, Mineraloberfläche und Luft in einem physikalisch präzisen Gleichgewicht aufeinandertreffen und die als kupfern-rosafarbener Halo um die gerippte Cuticula schwingt. Jenseits der Meniskuskuppel löst sich der Luftraum nicht in Klarheit auf, sondern verbleibt als trüber, amber-brauner Dunst, in dem keine Horizontlinie existiert – nur ein sanfter Übergang vom Spiegelglanz der Grenzfläche in jene diffuse organische Wärme, die erlischt, sobald das nächste Korn den Lichtweg abschneidet.

Im gesamten Sichtfeld wölben sich drei weiche, gedrungene Lippen aus durchscheinendem Gewebe — ihr Oberfläche fein gerippt, in nassem Perlmutton schimmernd, und in ihrer Mitte öffnet sich ein Y-förmiger Schlund, der rhythmisch pulsiert und Licht wie Prismen bricht. Auf der Flanke, kaum breiter als ein paar Mikrometer, liegt die Amphidenpore eingesenkt in eine seichte Hautfalte: ein mondsichelförmiger Kanal, von polierten Hüllzellen gesäumt und gefüllt mit einer viskosen Flüssigkeit, aus der zwölf Zilien als feines Parallelgeflecht auftauchen — jede Spitze mit gefalteten Rezeptormembranen besetzt, die ein blassblau-weißes Glühen aufnehmen, als würde chemische Information direkt als Licht sichtbar. Das Amphid ist das wichtigste Sinnesorgan des Nematoden: ein hochspezialisiertes Chemorezeptor-Ensemble, das gelöste Moleküle aus der wässrigen Bodenfilmschicht abtastet und dem Tier erlaubt, Nahrungsquellen aufzufinden oder Schadstoffe zu meiden — alles mit einer Nervenbahn von gerade 302 Neuronen. Rund um den Vorderpol legt der dünne Wasserfilm, der hier alles bedeckt, die chemische Landschaft offen: tiefes Cyan quillt aus einer Attraktantquelle nahe am Bildrand, verläuft durch Grün in warmes Bernstein — eine atmosphärische Farbfläche, die molekulare Konzentrationsgefälle als Ombré aus Licht und Farbe verkörpert und zeigt, dass für dieses Tier Geruch keine abstrahierte Wahrnehmung, sondern das unmittelbar sichtbare Relief seiner Welt ist.
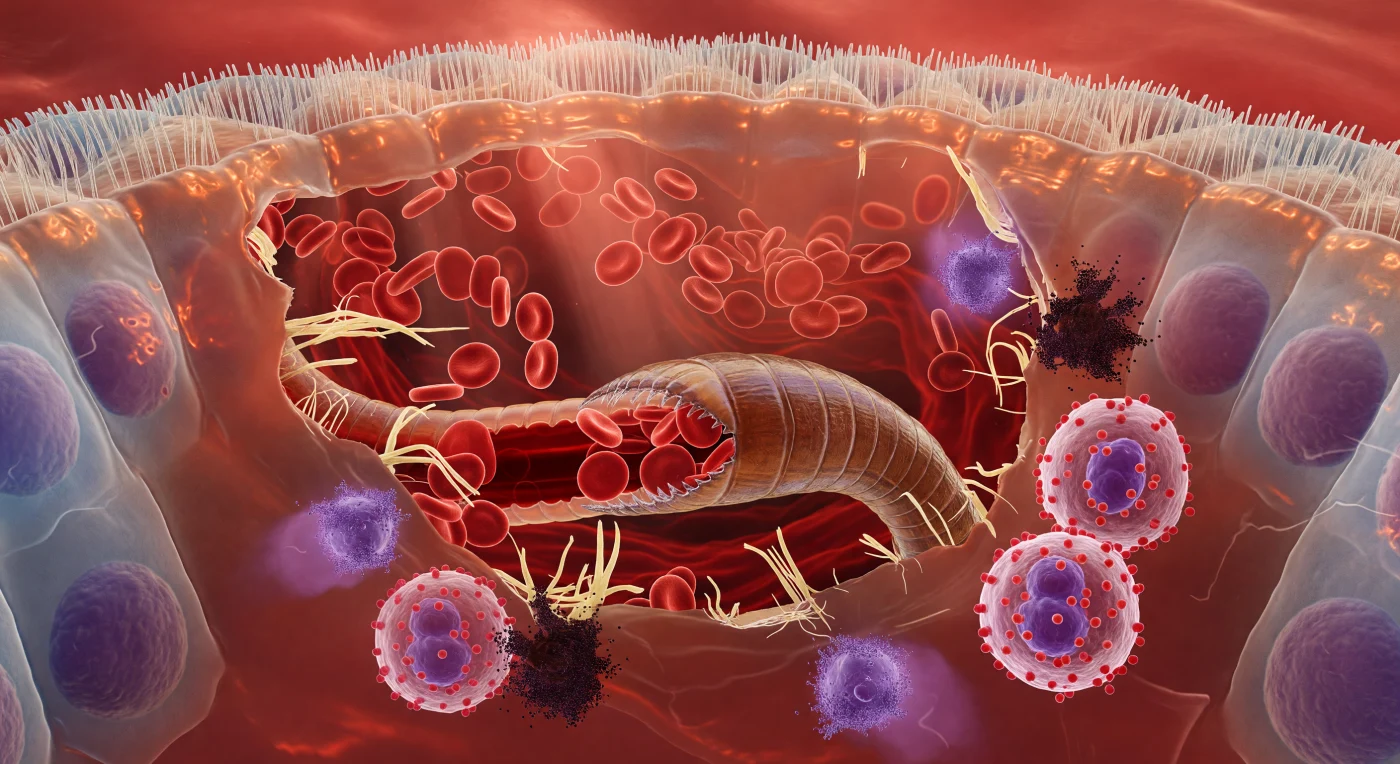
Du schwebst in der warmen, karmesinrot leuchtenden Tiefe einer Dünndarmzotte, umgeben von einer lebenden Architektur aus geschichteten Geweben, deren jede Oberfläche mit extrazellulärer Flüssigkeit überzogen ist wie Glas im Regen. Über dir bilden die dicht gepackten Mikrovilli des Bürstensaums ein unregelmäßiges, feuchtes Gewölbe, während die Hakenwurm-Bukkalkapsel – ein gehärtetes, bernsteinbraunes Chitingehäuse mit messerscharfen Schneidplatten – die zerrissene Submukosa wie ein in weiches Mauerwerk getriebener Rammbock auseinanderdrängt und zerrissene Kollagenfasern wie gebrochene Tragseile in den Wundspalt freisetzt. Aus den geplatzten Kapillaren strömen bikonkave Erythrozyten in dichten, langsam taumelnden Strömen hervor – jede Zelle eine transluzente Scheibe von leuchtendem Scharlachrot, die im durchfallenden Gewebslicht wie ein Glasmedaillon aufglüht und sich in pulsierendem Rhythmus in den Schlund des Wurms ergießt, dessen eigener Darm durch die irisierend-korrigierte Kutikula als dunkelroter, arteriell gefüllter Schlauch erkennbar ist. Am Wundrand löst sich die Immunantwort wie ein Wettergeschehen: Eosinophile drängen sich in das zerrissene Gewebe, ihre riesigen lachsfarbenen Granula vibrierende Reservoire komprimierter Biochemie, während Mastzellen in langsamen, tintenartigen Purpurwolken explodieren und basophile Granula sich in federartigen Schlieren durch die Gewebsflüssigkeit ausbreiten – alles durchdrungen von der roten, lebendigen Wärme eines Körpers, der noch nicht weiß, dass er von innen verzehrt wird.