Confianza científica: Muy alto
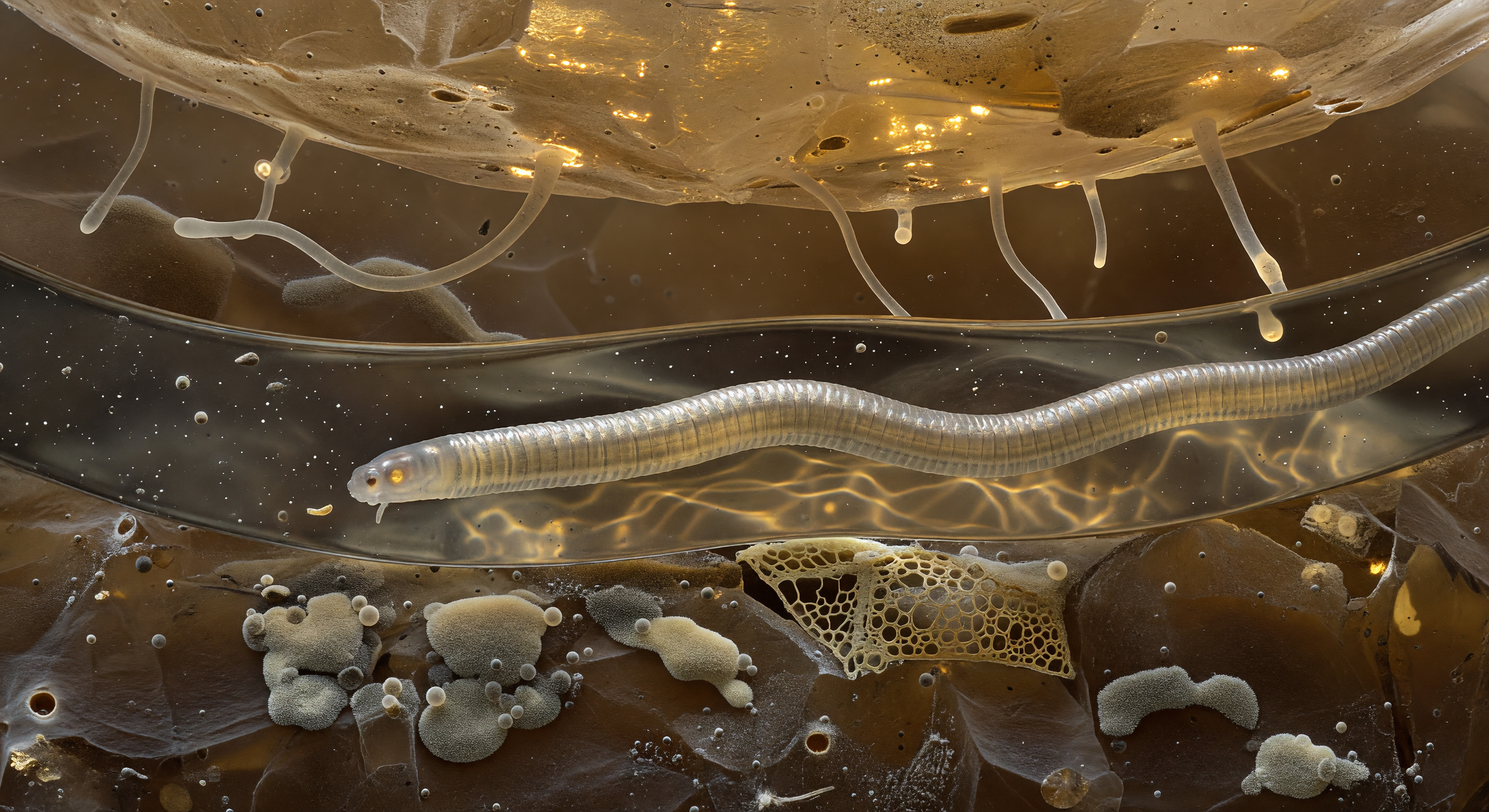
Desde el interior mismo del corredor acuoso, el mundo nematodo se revela como una catedral de física y biología entrelazadas: dos colosos de cuarzo transparente flanquean el túnel como acantilados cristalinos, sus superficies vivas con una laca iridiscente de biofilm bacteriano que destella en bronce, violeta y aguamarina bajo la luz refractada. La geometría del corredor no la dicta la roca sino la tensión superficial, esa fuerza que a esta escala domina con la autoridad de la gravedad, arqueando los meniscos en los extremos del túnel como espejos parabólicos que inundan el interior de una claridad fría y casi quirúrgica. Sobre el vacío, hifas fúngicas translúcidas se tienden de pared a pared en arcos tensos como cables de suspensión, mientras el propio cuerpo del gusano —un cilindro prismático de cutícula anulada, lleno de gránulos intestinales ámbar-amarillos autofluorescentes que tiñen el agua circundante de un oro difuso— presiona suavemente contra el suelo de película líquida donde la inercia no existe y cada ondulación muscular debe negociarse con la viscosidad a un número de Reynolds cercano a cero. Más allá de los meniscos ardientes se extiende la verdadera oscuridad del suelo, una penumbra ocre-marrón de masas minerales vagamente intuidas, recordatorio de que este corredor luminoso y vivo es un mundo completo encerrado entre dos granos de arena.

Te encuentras suspendido en el corazón geométrico del bulbo faríngeo terminal de un nematodo, una cámara de apenas cuarenta micrómetros de diámetro que, a esta escala percibida, se despliega como una catedral abovedada donde las paredes musculares se curvan en todas direcciones como las costillas de una nave románica. Tres placas trituradoras de cutícula electrodensa, del color del cuerno ambarino, convergen rítmicamente a lo largo de sus suturas radiales, comprimiendo el lumen en estrella con una violencia que desplaza el fluido faríngeo en lentas láminas viscosas que se doblan como vidrio caliente. Los haces musculares estriados que te rodean muestran, en su citoplasma casi translúcido de tonos salmón, la alternancia pálido-oscura de cada sarcómero, con un brillo azul-plata fugaz donde las redes de actina y miosina se alinean antes de relajarse de nuevo al rosa cálido. Fragmentos bacterianos destrozados —fantasmas traslúcidos con membranas fosfolipídicas aún iridiscentes en sus bordes— derivan perezosamente en el fluido amarillento y proteico, mientras esferas perfectas de lípidos dorados flotan como pequeñas lentes, proyectando imágenes invertidas de las placas trituradoras sobre su propia pared curva. Todo el interior, bañado por una luminiscencia ambarína difusa que se filtra a través del músculo y la cutícula laminada, permanece suspendido entre un aplastamiento y el siguiente, un mundo de presión hidrostática y deriva browniana donde la gravedad no existe y la viscosidad lo gobierna todo.

En el borde superior de un grano de cuarzo fracturado que se alza como un acantilado de cristal, una larva dauer se mantiene erguida sobre la punta de su cola en un equilibrio improbable, su cuerpo ámbar translúcido elevándose como una torre solitaria contra la penumbra ocre del suelo. La cutícula engrosada de la larva —una adaptación de resistencia que puede sobrevivir meses sin alimentarse ni moverse— brilla con anillos finos bajo una luz difusa y envolvente, mientras que a través de su pared corporal semitransparente se distinguen grandes gotas de grasa cremosas, reservas energéticas esféricas suspendidas como perlas en resina, testimonio silencioso de un organismo que ha pausado su biología entera para esperar condiciones mejores. La boca está sellada por un tapón bucal opaco, el extremo anterior curvado ligeramente en el aire quieto, sin apertura visible, como un capullo que guarda su forma pero ha renunciado por ahora al mundo. Muy abajo, al pie del grano que desde aquí parece una catedral mineral, se adivina entre la oscuridad cálida un laberinto de fragmentos de suelo, hifas fúngicas pálidas tendidas como cables entre masas de partículas lejanas, y una bruma tenue de materia orgánica disuelta que comprime la perspectiva y hace de este diminuto pilar ámbar —de apenas un milímetro de altura— el eje vertical de un mundo antiguo y fracturado.
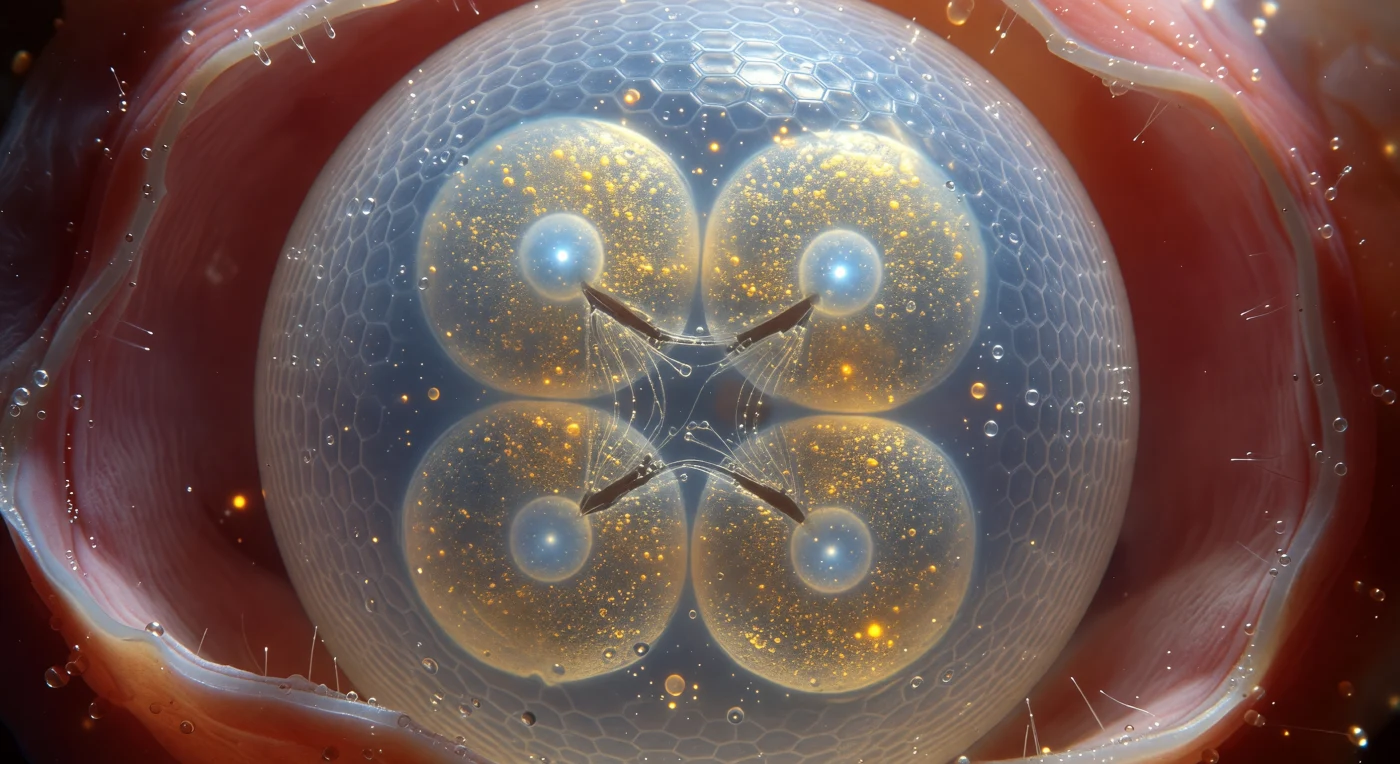
Hovering a fracción de milímetro de distancia, la pared curvada del huevo de nematodo llena todo el campo visual como la superficie de una luna helada vista desde un módulo de aterrizaje: una membrana translúcida de quitina —polisacáridos entrecruzados en capas comprimidas— dispersa la luz polarizada en bandas ondulantes de índigo y plata que recorren su domo con cada microdesplazamiento de presión interna. Al otro lado de esa barrera, cuatro blastómeros recién divididos se comprimen entre sí en disposición tetraédrica, sus superficies de contacto aplanadas en surcos de segmentación oscuros y geométricos, mientras sus interiores refulgen en oro cálido por los gránulos de vitelo —reservas lipoproteicas que alimentarán cada división futura— y cada núcleo flota como una esfera de hielo azul pálido con su nucléolo brillando en su interior como una perla fría. Entre dos de los blastómeros persiste el fantasma de un huso mitótico: cables de microtúbulos de plata viva tensados entre masas cromosómicas que aún no han terminado de segregarse, destello efímero de la maquinaria molecular más precisa que existe antes de que se desmonte en segundos. Todo este universo sellado y autosuficiente —con su fluido peivitelino perfectamente diáfano actuando como lente de inmersión natural y el tejido uterino rosa coral cerrándose en suave penumbra alrededor— lleva ya programada en esas cuatro células la totalidad del destino celular del organismo: cada una de las 959 células somáticas que conformarán el adulto está ya decidida, inscrita en un linaje tan determinístico como un plano arquitectónico.

La superficie que se extiende ante el observador es un mundo de llanuras corrugadas perfectamente paralelas, cada cresta de anulación curvándose de horizonte a horizonte como dunas de arena petrificadas en oro pálido y gris plateado, sus cimas suavemente redondeadas y casi vítreas bajo una iluminación oblicua que talla las depresiones entre crestas en sombras absolutamente negras. Esta arquitectura repetitiva no es ornamental sino estructural: la cutícula del nematodo es una matriz extracelular multilaminar secretada por la hipodermis subyacente, un cilindro hidrostático presurizado cuya integridad mecánica depende precisamente de esta geometría anular que distribuye la tensión circunferencial y permite la ondulación sinusoidal del cuerpo durante la locomoción. A la izquierda, las alae laterales se elevan como una cordillera montañosa continua, su meseta superior con estrías transversales más finas que brillan con un oro ligeramente más cálido, proyectando una sombra larga y precisa sobre el valle más cercano — estas estructuras longitudinales son engrosamientos cuticulares que sirven como guías mecánicas durante la locomoción en superficies sólidas. Cerca del horizonte anterior, varias papilas hemisféricas de las sensilas cefálicas emergen de la llanura corrugada como formaciones silenciosas y monolíticas, cada una coronada por un único microporo apical que atrapa un destello especular brillante, revelando su función como orgánulos mecanosensoriales y quimiosensoriales que conectan el entorno exterior con el sistema nervioso del animal. Todo el paisaje existe a una escala donde la gravedad es irrelevante, la tensión superficial gobierna cada interacción y este material — orgánico pero casi mineralizado, cálido y presurizado, silenciosamente vivo — constituye la frontera completa entre el animal y su mundo.

Presionado contra la superficie del agar, el mundo entero se despliega como un diorama retroiluminado: una alfombra densa e impenetrable de bastones de *E. coli* se extiende hasta cada horizonte, sus siluetas oscuras y ligeramente iridiscentes recortadas contra la luz blanca fría que sube desde abajo, y detrás de ti, una nave catedralicia de agar limpio se curva en la firma sinusoidal de tu propio paso, con paredes verticales de células bacterianas cortadas a ras que centellean con un filo azul-blanco en sus caras expuestas. En el plano medio, otros rastros de sinuosidad distinta cruzan el campo en distintas orientaciones, sus arcos superpuestos formando un mosaico complejo de terreno despejado y terreno en vías de recolonización, donde las bacterias ya comienzan a invadir los bordes más antiguos con una neblina punteada. Justo ante ti, el bulbo terminal de la faringe late en contracciones rítmicas de cuatro golpes por segundo, succionando bastones individuales hacia el interior de la cavidad bucal mientras el intestino se va tiñendo progresivamente de un cálido marrón anaranjado, los gránulos de sus células emitiendo destellos autofluorescentes de un amarillo-verdoso fugaz bajo la luz. Tu cuerpo entero —luminoso, arquitectónico, corrugado por las anulaciones de la cutícula que captan reflejos especulares del suelo brillante— es a la vez máquina y paisaje, una criatura transparente suspendida en un universo regido no por la gravedad sino por la tensión superficial y la viscosidad de un mundo de película de agua donde cada movimiento exige esfuerzo muscular continuo.
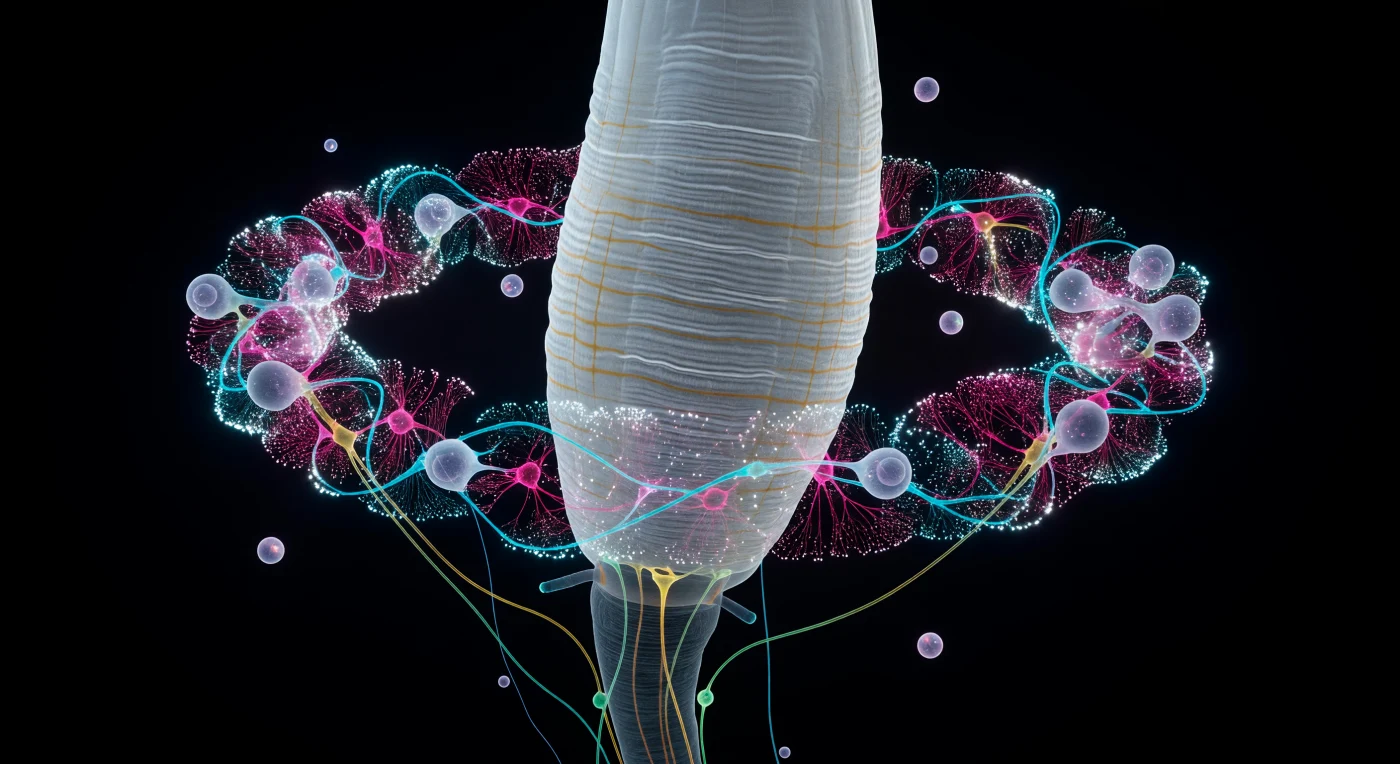
El observador flota suspendido en el vacío pseudocelómico a menos de diez micrómetros de la pared faríngea: ante él se eleva una vasta columna de músculo estriado, translúcida como marfil envejecido, sus crestas longitudinales y corrugaciones transversales talladas con una precisión que recuerda la arquitectura de un glaciar a escala continental. A su alrededor, el anillo nervioso circunesofágico ciñe la faringe como una corona de fuego electroquímico, apenas veinte micrómetros de diámetro pero empaquetado con una complejidad que desafía la comprensión: fascículos de axones presionados entre sí con separaciones de nanómetros, neuronas sensoriales anfidiales en cian glacial tejiendo caminos desde la periferia, interneuronas en magenta profundo pulsando bajo la superficie del neuropilo, y comisuras motoras en verde-amarillo que arquean hacia la oscuridad donde comenzará el cordón nervioso ventral. Cada cuerpo celular de tres a ocho micrómetros orbita el conjunto como un pequeño planeta de citoplasma translúcido, y entre ellos, los cúmulos de vesículas sinápticas estallan como supernovas blancas, instantes de fusión de membrana saturada en calcio congelados en la oscuridad permanente del interior del suelo. Este anillo —el centro de integración sensorial y motora de un animal con exactamente 302 neuronas y más de 7 000 conexiones sinápticas completamente cartografiadas— arde en silencio dentro de un organismo que navega su mundo bacteriano a un largo de cuerpo por segundo, sin ojos, sin cerebro convencional, guiado únicamente por esta diadema de luz intrínseca y señal química.
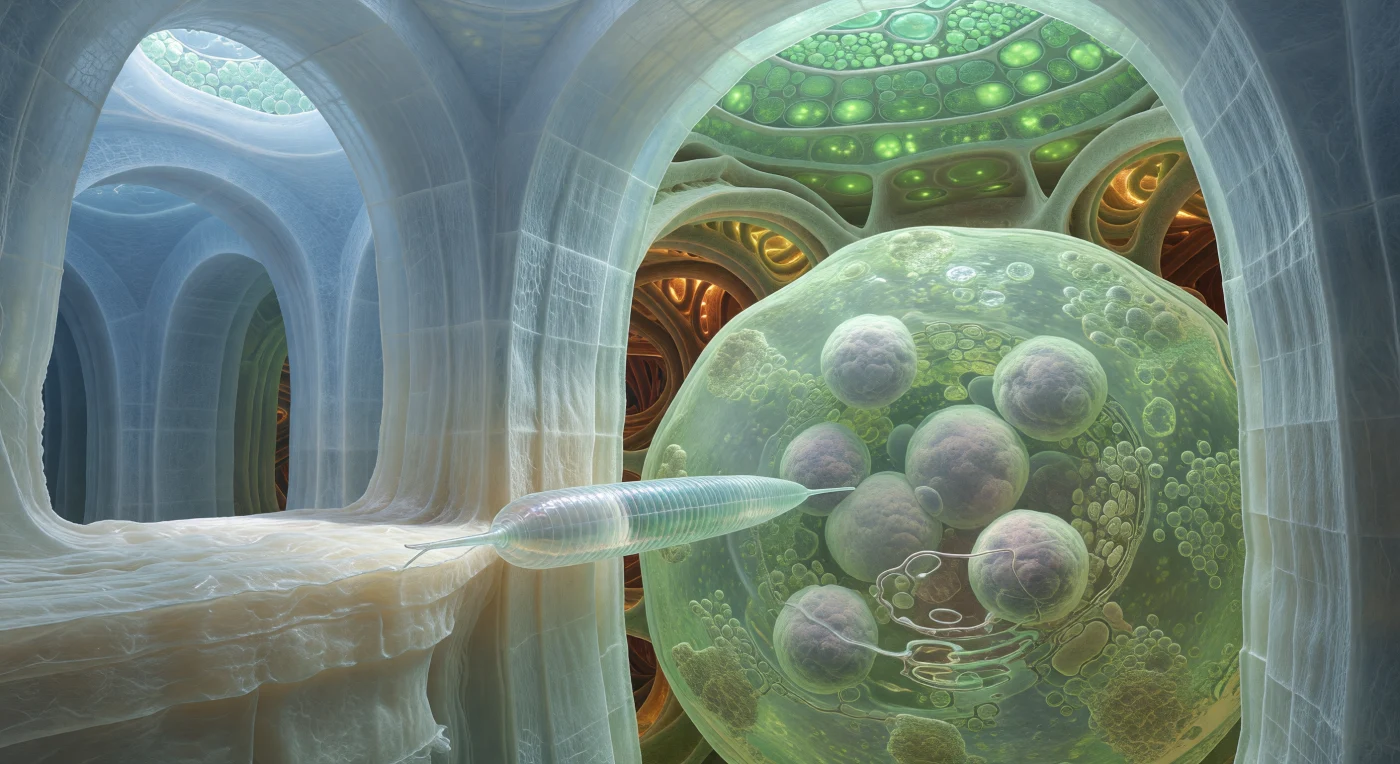
Nos encontramos suspendidos en la juntura de dos paredes celulósicas colosales, estructuras laminadas de microfibrillas comprimidas que se elevan como los pilares interiores de una catedral inundada, su superficie entretejida con una textura de retícula fina que capta la luz jade filtrada desde la epidermis lejana. Contra esta muralla de marfil traslúcido avanza el juvenil del nematodo *Meloidogyne*, un cilindro presurizado de cutícula anulada que empuja su estilete —una aguja hueca de nanómetros de apertura pero de fuerza mecánica suficiente para perforar esta pared— en busca del control metabólico que transformará una célula ordinaria en una factoría de alimentación permanente. Más allá de la pared, la célula gigante ya transformada se despliega como una sala inundada de citoplasma verde-limón turbio de ribosomas, donde múltiples núcleos hipertrofiados flotan como lunas de lavanda pálida, su contenido genético amplificado por rondas sucesivas de endorreduplicación que elevan la ploidía hasta 64 veces el valor normal. En el fondo profundo, los vasos del xilema de paredes espiralizadas y lignificadas irradian un fulgor ámbar-siena como circuitos de alta tensión biológica, canales hidráulicos que el parásito interceptará para drenar el flujo continuo de azúcares y aminoácidos hacia su propio cuerpo. Todo el espacio intersticial está bañado en el apoplasto acuoso, una película viscosa que conecta cada superficie en un continuo húmedo sin sombras nítidas, solo gradaciones cromáticas que van del azul-blanco glacial de la celulosa más próxima al verde difuso del citoplasma hasta el oro profundo que pulsa en los corredores vasculares más distantes.
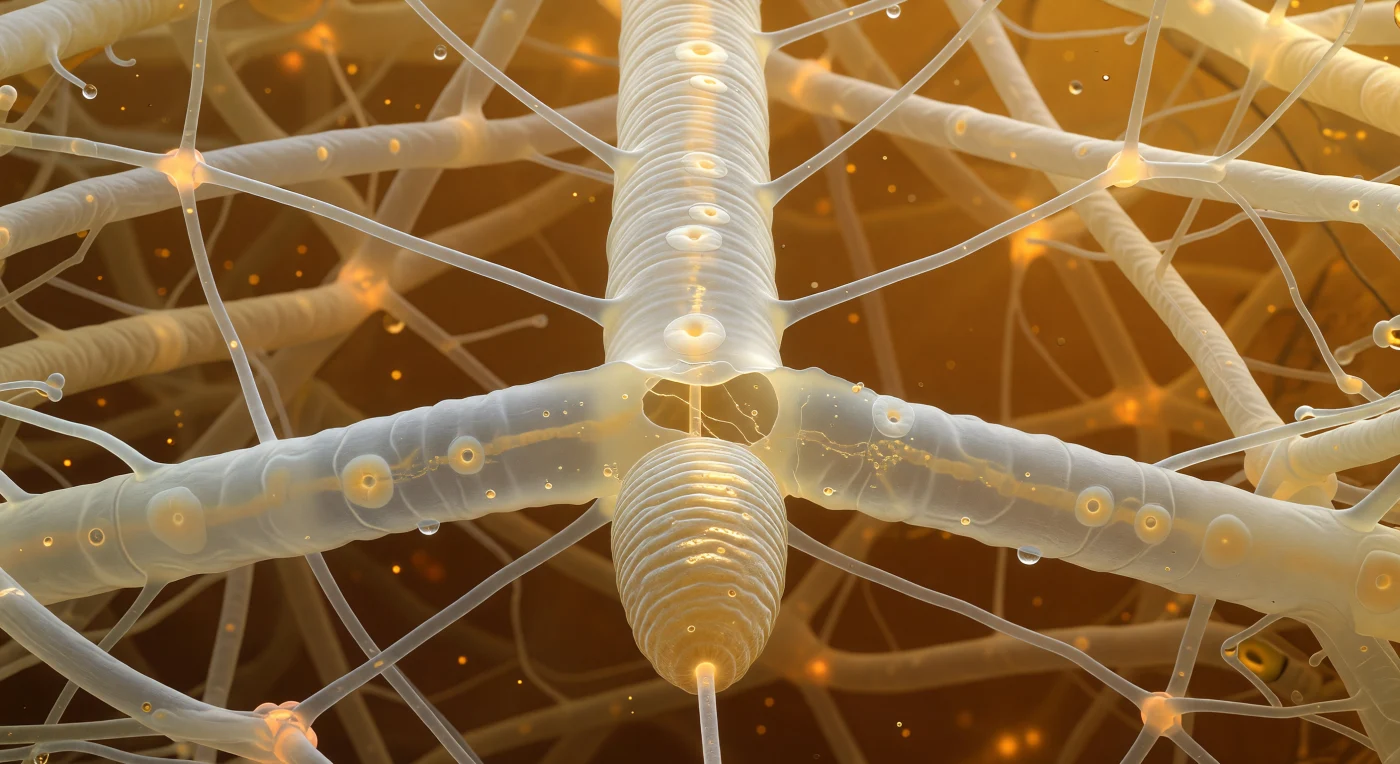
Dentro de este laberinto de filamentos vivos, te encuentras suspendido frente a una hifa que ocupa tu campo visual como una tubería presurizada de marfil translúcido, sus paredes levemente corrugadas por los septos que aparecen como discos opalinos a intervalos irregulares, mientras el citoplasma teñido de ámbar fluye en ríos lentos de orgánulos granulares hacia el interior. La red micelial se ramifica y anastomosa en todas direcciones a tu alrededor, sus uniones intercilares emitiendo un calor anaranjado donde los puentes citoplasmáticos pulsan con actividad metabólica compartida, y todo ello descansa sobre una celosía estructural de fibras de celulosa en descomposición que ancla el conjunto como los arbotantes de una catedral orgánica. En el primer plano, la cabeza del nematodo fungivoro se apoya contra la hifa: un cilindro de cutícula anillada cuyas crestas capturan la luz ámbar con un brillo casi iridiscente, mientras el estilete —una aguja rígida y refractante de quitina— ya ha perforado la pared hiphal y la bomba faríngea del animal pulsa a más de doscientas veces por minuto para extraer el contenido celular. Desde el punto de punción se expande una ola de retracción pálida: el citoplasma huyendo de la herida, los orgánulos desviándose alrededor de la lesión como un río que rodea un obstáculo, dejando a su paso una zona de menor densidad óptica y diminutas gotas de material extraído que refractan la luz ámbar en puntos cálidos y especulares. Todo el mundo visible está construido con una paleta estrecha —ámbar quemado, marfil, crema cálida— en la que el destello frío y casi metálico del estilete es el único elemento inorgánico en un universo enteramente vivo.

En el pasillo intersticial que se abre entre los granos de arena —bóvedas silíceas pulidas a la escala de colinas suaves—, el agua marina lo envuelve todo como un cristal ligeramente turbio, cargado de sales y arcilla coloidal que difumina los bordes lejanos en una neblina azul sin origen preciso. Las frústulas de diatomeas se acuñan en cada fisura como catedrales de sílice biogénica, sus hileras de punctae y sus bandas de cíngulo capturando la poca luz disponible y devolviéndola en aureolas ámbares y doradas, mientras la biopelícula bacteriana de azufre —una membrana iridiscente de rosas y violetas que cambia de tono con cada variación de ángulo— reviste las superficies redondeadas de los granos como una piel que respira en pulsos cromáticos lentos. Por el centro de la escena avanza el nemátodo: un cilindro de marfil con los anillos cuticulares perfectamente visibles, cada annulus cubierto de partículas de sedimento y fragmentos de diatomea adheridos como incrustaciones de joyería, un organismo que no nada sino que sinúa con esfuerzo muscular constante a través de un medio tan viscoso que la inercia carece de sentido, donde la tensión superficial y la presión hidrostática interna de su propio cuerpo gobiernan cada movimiento. A la derecha, emergiendo entre dos granos-colinas, la concha de foraminífero se eleva en calcita crema como una nave gótica de cámaras apiladas, cada apertura un arco oscuro que conduce a sombras interiores, mientras desde el techo invisible de este mundo descienden las setas doradas de un poliqueto como columnas de una arquitectura que pertenece a otra escala completamente diferente.
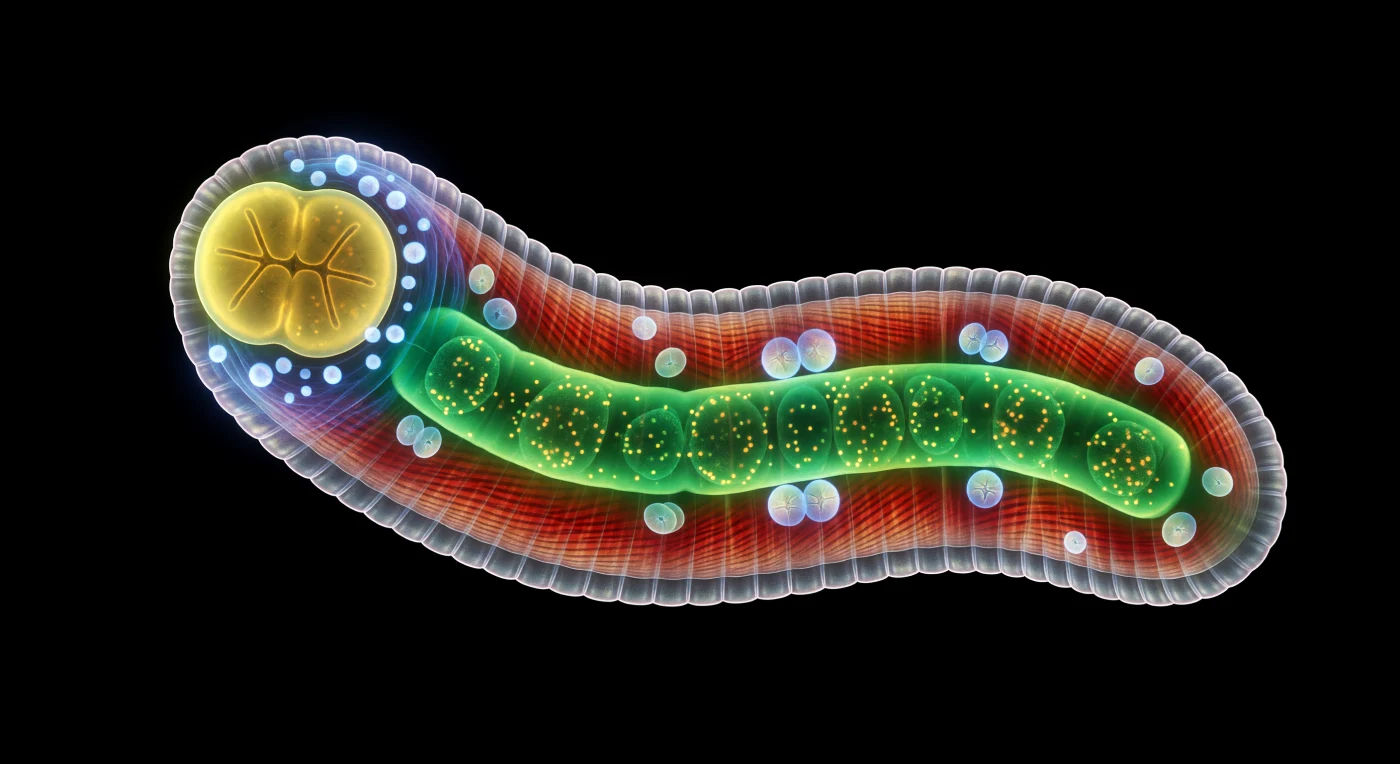
En la oscuridad absoluta, un cuerpo se curva ante ti como una metrópolis iluminada desde dentro: el hermafrodita adulto de *Caenorhabditis elegans* en su postura sinusoidal característica, con cada tejido emitiendo su propia longitud de onda como si la biología fuese una forma de arquitectura lumínica. Las cuatro bandas longitudinales de musculatura corporal arden en rojo carmesí atravesadas por estrías de sarcómeros diagonales, mientras el intestino central — un tubo de esmeralda saturada cargado de gránulos autofluorescentes en movimiento lento y viscoso — proyecta destellos jade sobre los tejidos circundantes, revelando la transparencia radical de un organismo que no oculta ninguno de sus procesos internos. Hacia el extremo anterior, el faríngeo bilobulado reluce en amarillo-verde citrino mientras un collar de puntos cian-blanco — los cuerpos celulares del anillo nervioso — corona la región cefálica con una aureola de luz helada, recordándonos que en este animal cada uno de sus 302 neuronas está cartografiada, cada sinapsis catalogada, cada destino celular conocido desde el primer cigoto. En el útero, embriones opalescentes en azul-blanco frío flotan como perlas en división, sus surcos de escisión trazando geometrías de blastómeros que reproducen, en miniatura, el mismo programa de desarrollo conservado a lo largo de cientos de millones de años de evolución. Todo existe contra un negro sin fondo que convierte al animal en la única fuente de luz en su universo, un circuito de joyería viva donde la física del bajo número de Reynolds hace de cada movimiento ondulatorio un acto de precisión mecánica continua.

Te encuentras suspendido en el umbral de una oscuridad aniquiladora: la cavidad bucal de un *Mononchus* depredador que, a esta escala, se lee como la entrada a una catedral esculpida en ámbar vivo. El diente dorsal curvado se alza ante ti como un arco monolítico de cutícula esclerotizada, con una translucidez rojo-marrón profunda que refracta la luz difusa del suelo en gradientes de caramelo y ocre quemado a lo largo de su borde afilado, mientras los dentículos secundarios —dispuestos en anillos concéntricos de marfil pálido— se pierden en la oscuridad pulsante del lumen triradial faríngeo. Parcialmente engullida en este espacio, una nemátoda bacterívora más pequeña se comprime contra la pared de la cámara bajo la succión: su cutícula, normalmente un cilindro liso y presurizado de translucidez perlada, se arruga ahora en pliegues diagonales que se propagan desde el punto de ingestión, mientras a través de ella aún se distinguen el faringe bilobulado, la gónada sinuosa como un hilo luminoso, y el intestino cuyos gránulos brillan con un tenue verde-dorado, últimos destellos de un esqueleto hidráulico que cede. El entorno es suelo de ocre oscuro —granos de arena y limo comprimidos en paredes de acantilado rugoso, recubiertos de biopelícula bacteriana que iridisce con humedad opalescente— mientras la luz, llegada como una niebla cálida desde la izquierda, se desvanece en la negrura absoluta de la garganta depredadora, haciendo de esta escena algo simultáneamente íntimo e insondable.
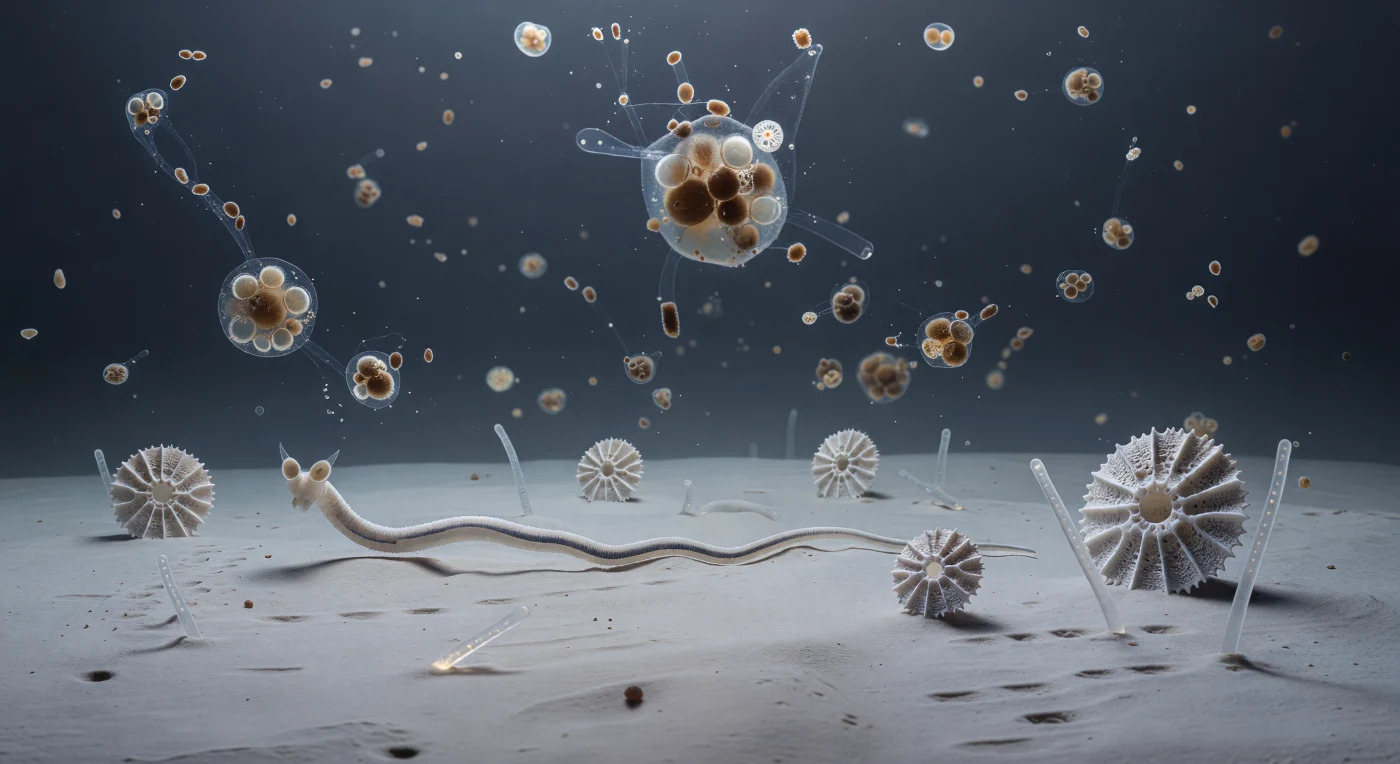
En este lugar donde la gravedad casi no importa y la presión del agua equivale a cientos de atmósferas, el suelo abismal se extiende como una llanura de arcilla pálida y hueso viejo, casi sin textura a esta escala, salpicada de carcasas blancas de foraminíferos que se alzan como catedrales calcáreas y espículas de esponja silíceas que yacen como torres derrumbadas. Desde arriba, a una altura incomprensible en el vacío azul-gris, descienden lentamente agregados de nieve marina —fragmentos de diatomeas, pellets fecales y hebras de mucopolisacáridos— girando con una suavidad que solo permite el agua casi tan densa como ellos mismos, cada uno una fuente concentrada de materia orgánica para un ecosistema que vive de la lluvia lentísima del mundo superior. Los nemátodos que se mueven sobre este sustrato son cilindros translúcidos y finamente anulados, de apenas una fracción de milímetro de diámetro, cuya locomoción sinusoidal resulta tan pausada que parece geológica: sus grandes órganos amfidiales —cavidades quimiorreceptoras laterales desproporcionadamente anchas respecto al cuerpo— escanean el agua intersticial en busca de las trazas moleculares que delatan la proximidad de un agregado orgánico. Todo aquí funciona en el régimen de bajo número de Reynolds, donde la viscosidad domina sobre la inercia y cada ondulación muscular debe sostenerse de manera continua, convirtiendo la quietud extrema de este desierto polar submarino no en ausencia de vida, sino en vida calibrada al tempo más austero del planeta.

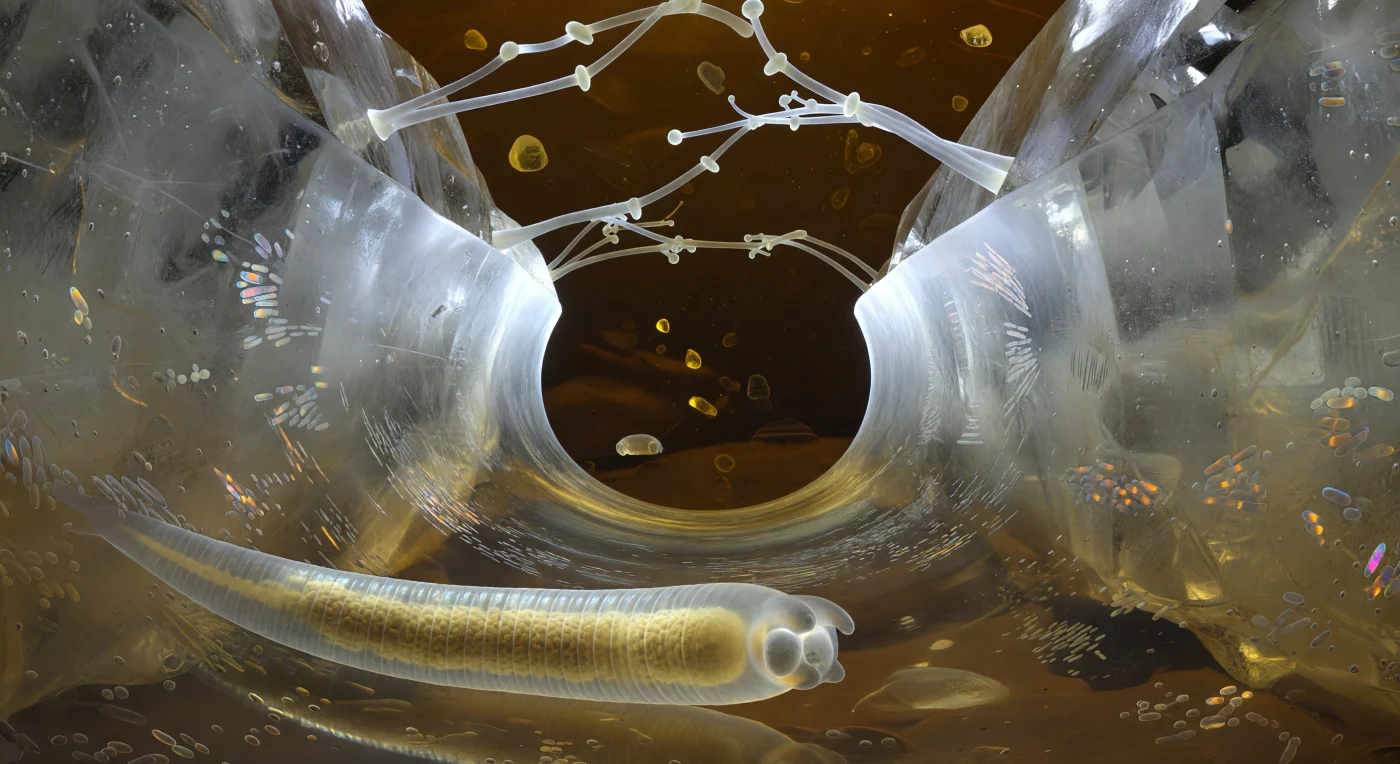
Suspensos en el interior de un puente capilar no más ancho que un cabello humano, el ojo se alza hacia una cúpula de menisco que se curva como el techo de una burbuja de jabón prensada contra el vidrio, su superficie tensada por las fuerzas capilares en una geometría convexa que actúa como un espejo ojo de pez de extraordinaria precisión óptica, reflejando y aplastando radialmente todo el paisaje circundante —granos de cuarzo como acantilados de vidrio esmerilado, filamentos orgánicos oscuros como cuerdas de brea, corredores turbios de agua intersticial viscosa— en una sola panorámica distorsionada que tiembla con cada vibración transmitida por el fluido. La línea de contacto trifásica donde el agua, el aire y el mineral convergen arde como un aro iridiscente de oro blanco que se curva hacia el cobre y el rosa, concentrando la refracción en un halo continuo que envuelve también el borde visible de la cutícula del nematodo, con sus anillos circumferenciales captando destellos ámbar en cada cresta y dejando caer sombra fría entre los surcos. El agua que nos rodea no es transparente sino de un marrón ambarino profundo, el color del té viejo ante la luz de la tarde, teñida por ácidos húmicos disueltos de la hojarasca en descomposición, mientras plaquetas de caolinita e ilita de apenas cientos de nanómetros derivan en arcos erráticos a través del medio, cada una captando la luz dispersa de forma distinta al girar —plata, luego oscuro, luego oro oxidado— en el movimiento Browniano perpetuo de este mundo donde la gravedad carece de sentido y la tensión superficial lo gobierna todo. Más allá del domo del menisco, el espacio aéreo no ofrece un cielo limpio sino una neblina ambar-marrón difusa, turbio de partículas suspendidas y vapores orgánicos dispersos, sin horizonte ni aristas nítidas, solo un gradiente desde la superficie especular del menisco hacia la oscuridad densa donde el próximo grano mineral presiona y corta por completo la luz dispersa.

Te encuentras a pocos centímetros del extremo anterior de un nematodo, suspendido exactamente a su misma escala, y los tres labios romos que flanquean la boca ocupan todo tu campo visual como la entrada a una gruta submarina tallada en alabastro translúcido, sus anillos cuticulares capturando la luz dispersa en destellos nacarados que van del marfil al oro pálido. En el flanco lateral, hundida en una discreta ranura epidérmica, la apertura del anfidio se abre como una hendidura en media luna: un canal relleno de fluido viscoso y bordeado por células vaina de membrana lisa, dentro del cual se distinguen doce terminaciones dendríticas ciliadas dispuestas en fascículo apretado, cada filamento rematado en membranas receptoras plegadas que absorben los fotones disponibles como brasas frías de luz azul-blanca. Los anfidios son los principales órganos quimiosensoriales de los nematodos, capaces de detectar gradientes de moléculas disueltas a concentraciones nanomolares a través de los cilios del par de neuronas AWA y AWC, cuya integración de señales dirige comportamientos de atracción o aversión codificados en un sistema nervioso de exactamente 302 neuronas. El entorno químico no permanece invisible a esta escala: la película acuosa que recubre la cutícula se estratifica en capas cromáticas — cian glacial concentrado cerca de la fuente atractiva, que sangra a través de verdes acuosos hasta un halo ámbar cálido en los márgenes de aversión — y dentro del canal anfidial, el trazador fluorescente acumulado en las dendritas dibuja un resplandor lima fantasmal que es evidencia visible de una señal química siendo internalizada y transformada en comportamiento.
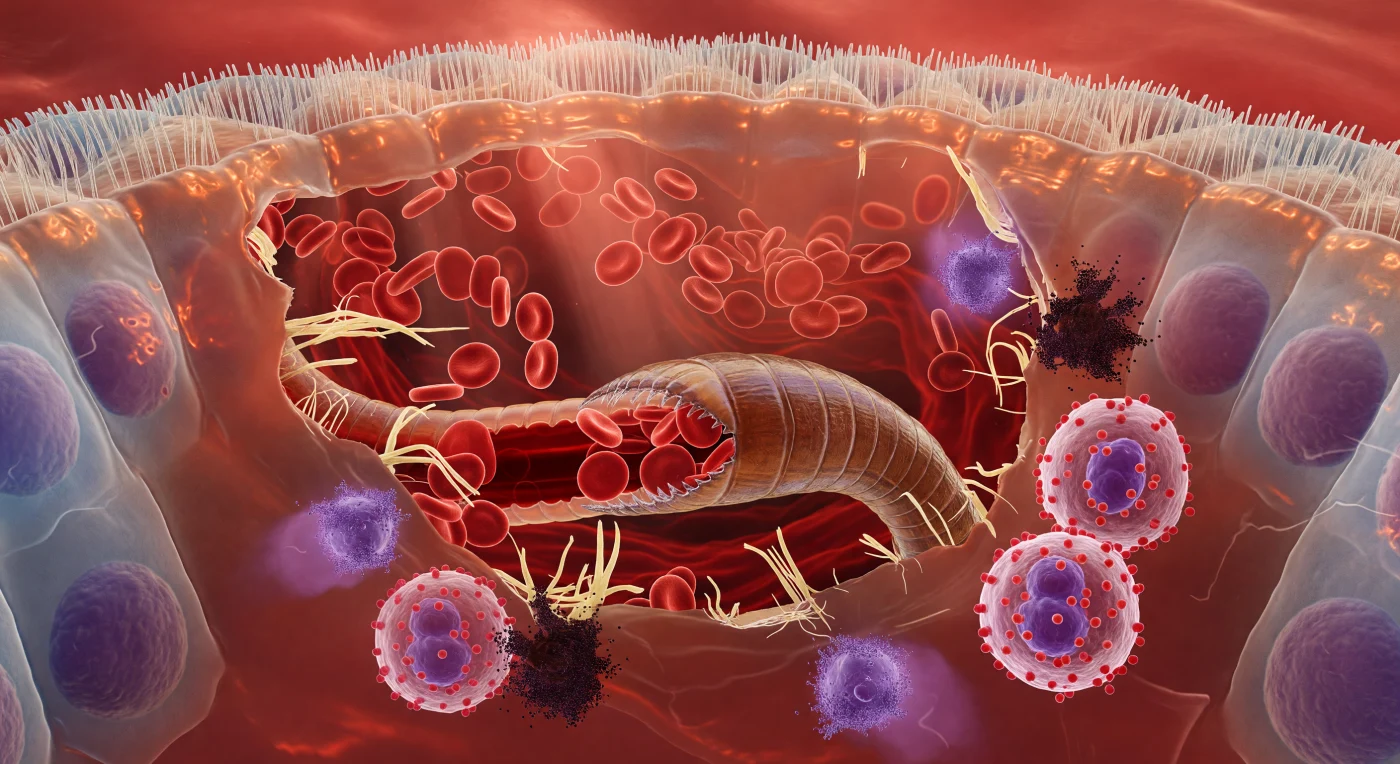
Te encuentras suspendido en el interior cálido y carmesí de una vellosidad intestinal mamífera, a la escala de un nematodo, dentro de una cavidad de herida que se percibe vasta y presurizada como el interior de una gruta inundándose. Sobre ti, el borde en cepillo apical de las células epiteliales columnares forma un techo irregular de adoquines: cada microvilliosidad es una cerda densa y translúcida, apretada hombro con hombro en un dosel ondulante que atrapa la luz ámbar-rojiza difusa del tejido y la refracta en estrías húmedas y doradas, mientras los núcleos ovales violáceos de las células se suspenden en sus tercios basales como linternas oscuras detrás de vidrio esmerilado. En el centro del campo visual, la cápsula bucal del anquilostoma domina como una máquina de asedio hundida en arquitectura blanda: sus paredes quitinosas endurecidas, de un ámbar marrón profundo con placas cortantes curvadas y serradas a escala micrométrica, están presionadas con firmeza en la submucosa desgarrada, mientras los glóbulos rojos biconvexos, cada uno un disco escarlata translúcido que brilla como medallón de vitral bajo la luz posterior del tejido, se vierten desde los capilares rotos en corrientes lentas y densas hacia la cavidad bucal abierta del gusano, cuyo intestino—visible a través de la cutícula iridiscente y corrugada del cuerpo—pulsa como un tubo carmesí arterial con sangre recién ingerida. En la periferia de la herida, el sistema inmunitario se manifiesta como un fenómeno meteorológico: los eosinófilos se agolpan en el margen tisular desgarrado con sus enormes gránulos de un rojo salmón que parecen vibrar con química contenida y sus núcleos bilobulados de un púrpura casi violeta, mientras las mastocitos detonan en nubes de material granular basofílico que se dispersan hacia afuera en lentas volutas púrpura-negras, como tinta caída en agua tibia, difundiéndose en plumas que fragmentan la luz transmitida en halos violetas sobre todo el ambiente suffuso de calor vivo, cada superficie curva y húmeda, sin geometría dura en ningún lugar, solo la suavidad presurizada de la carne bajo estrés y el pulso contráctil rítmico de un parásito alimentándose en el interior oscuro y húmedo de un cuerpo que no sabe que está siendo consumido.