
Im bernsteinfarbenen Wasser eines flachen Tümpels schwebt man auf Augenhöhe mit einem *Daphnia magna*, dessen zweiklappiger Chitinpanzer vom gelösten Huminsäuren zu einem warmen Honigton gefärbt wird und das Innere des Tieres wie durch getöntes Museumsglas freigibt. Das einzelne, große Komplexauge — eine dunkle, facettierte Kugel von erschreckender Präzision — dreht sich minimal in seiner Aufhängung, während das Herzmuskelband dorsal als roséroter Streifen rhythmisch pulsiert und der Darm in leuchtendem Smaragdgrün aufleuchtet, gespeist von chlorophyllreichen Algen. Im hinteren Brutraum drängen sich ein Dutzend Embryonen auf engstem Raum, die jüngsten noch als milchig-opake Kugeln, die älteren bereits mit schemenhaften Gliedmaßenknospen, die wie Hände gegen Mattglas gedrückt erscheinen. Über allem komprimiert das Snell'sche Fenster den gesamten Himmel in ein einziges gleißendes Oval aus warmem Bernsteinlicht, unterbrochen von der flachen, dunkelgrünen Silhouette eines Wasserlinsenlaubs, das wie ein ferner Kontinent über dieser mikroskopischen Welt treibt. Der gesamte Wasserkörper ist in eine träge, goldene Suspension feiner Detrituspartikel getaucht, die in langsamen Konstellationen aus Bronze und Gold durch das flüssige Medium driften — ein Universum aus lebendigem Glas, erfüllt von smaragdenem Feuer.
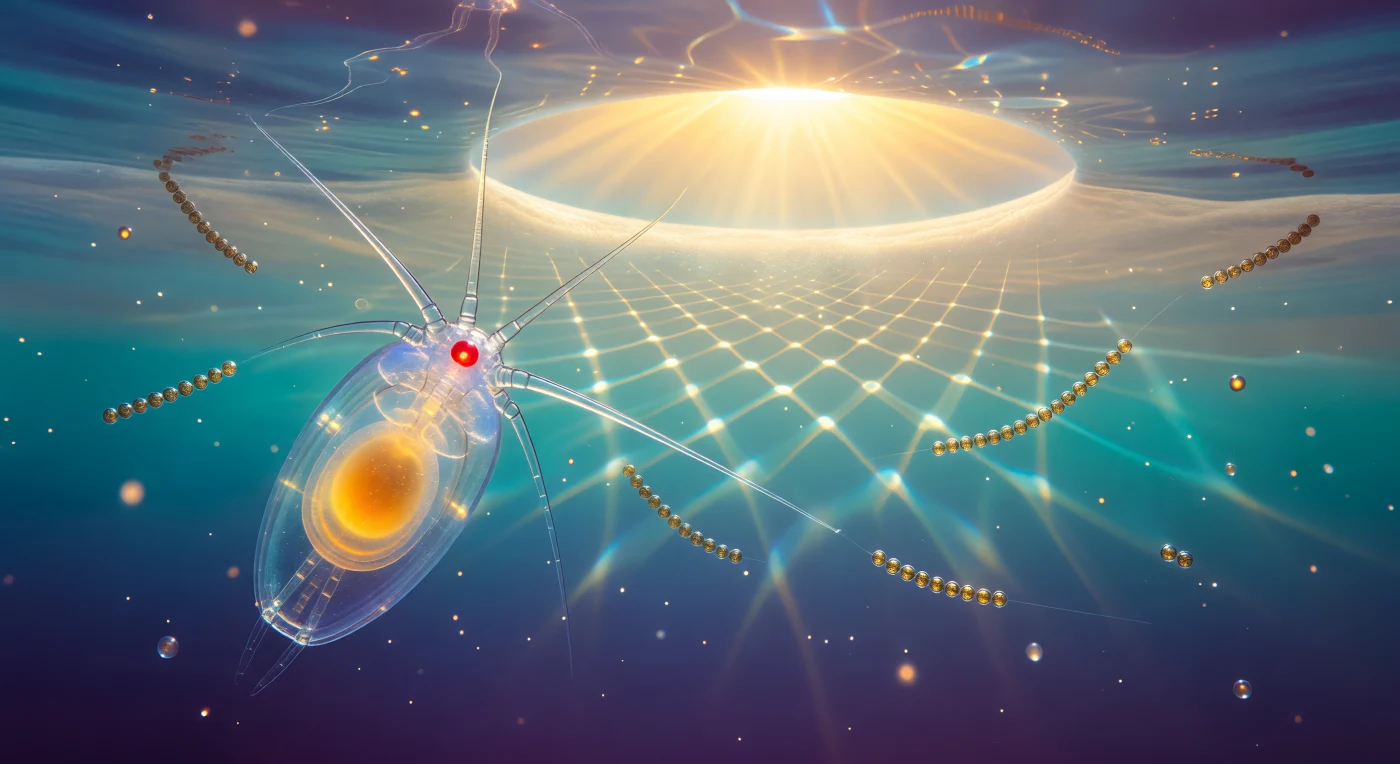
Wer hier schaut, schaut mit dem Auge eines Tieres — von einem einzigen rubinroten Naupliusauge aus, kaum größer als ein Staubkorn, eröffnet sich eine Welt aus flüssigem Glas und bewegtem Licht. Der Körper von *Calanus finmarchicus* füllt den Vordergrund wie eine lebendige Linse: vollkommen durchsichtig, seine Kutikula keine Oberfläche, sondern ein optisches Medium, das das türkisfarbene Wasser bricht und konzentriert, während der Lipidsack im Inneren warm-golden leuchtet — ein gespeicherter Vorrat aus Wachsestern, der dem Tier erlaubt, monatelang ohne Nahrung in die Tiefe abzusinken und wieder aufzusteigen. Über allem öffnet sich Snells Fenster als blazendes Oval aus Weißgold, der gesamte Himmel in einen 97-Grad-Kegel komprimiert durch die Totalreflexion der Wasseroberfläche, von dem aus Kaustik-Gitter — wandernde Netze aus Gold und Eisblau, geworfen von jeder Welle — über den Körper des Tieres ziehen und den Lipidsack für Augenblicke beinahe weiß aufflackern lassen. In dieser epipelagischen Zone bei zehn Metern Tiefe bewegt sich *Calanus* in einer Welt, die von Brechung regiert wird: Kieselalgen-Ketten aus *Chaetoceros* treiben wie gläserne Perlenschnüre im Blau, jede Chloroplastenmembran ein winziges Fenster für das Licht, das in dieser Wassertiefe noch reichlich vorhanden ist, aber mit jedem weiteren Meter exponentiell schwindet — sodass dreißig Zentimeter unterhalb bereits eine Dämmerung aus Indigo beginnt, kalt und still und unermesslich schwer.

In der tropischen Nacht, drei Meter über dem Korallenschuttergrund, gibt es kein Mondlicht, keine Streustrahlung, keine Orientierung — nur vollständige, salzige, schwere Dunkelheit, und dann die Männchen von *Vargula hilgendorfii*, die in ihr zu schreiben beginnen. Jedes Tier ist ein ovales Kalkschalengehäuse von kaum einem Millimeter Länge, dessen fein geripfte, porenbesetzte Valven von innen her mit einem milchig-perlmuttenen Schimmer glühen, erzeugt durch das eigene sekretierte Luciferin-Luciferase-System der Drüsenzellen — ein lebendiger Lichtsamen, frei schwebend im Nichts. Aus den Rostraldrüsen zwischen den geöffneten Valven werden paarweise Strahlen biolumineszenter Flüssigkeit in die Wassersäule gepumpt, die sich in arttypischen Spiralgeometrien entfalten: an der Quelle ein gesättigtes Aquaweiß, das durch Blaugrün, Blaugrau und Indigoblau in die umgebende Schwärze zerfällt, als wäre Blitzkraft in Wasser gefangen und langsam freigesetzt worden. Wo sich die Leuchtspur zweier Männchen kreuzt, entstehen kurze Knoten intensivierter Strahlung, die aufflackern und verlöschen, während der Korallenschutt darunter — Kalziumkarbonat, Korallinalgenfragmente, tote Acropora-Äste — nur als tiefblaugrüne Silhouette erahnbar ist, von den nächsten Leuchtspuren von unten her angestrahlt, ein lebendiger Sternkarte aus Chemie und Photonen im flüssigen Universum.

In diesem eingefrorenen Augenblick schweben wir auf Augenhöhe mit den Tieren selbst — ein Millimeter groß, getragen von Wasser, das sich wie reines, grüngetöntes Glas anfühlt. Vor uns stürzt sich ein *Mesocyclops leuckarti* auf eine Naupliuslarve: der kompakte, bernsteinfarben-orangerote Körper des Cyclopoiden ist halb opak wie poliertes Keramikpanzer, sein einzelnes, dunkelkarmesinrotes Naupliusauge starrt mit konzentrierter Intensität, während die raptorialen Maxillipeden als chitinöse Wuchtkeile bereits die glasklare Larve treffen — deren Körpermembran an der Kontaktstelle sichtbar eingedrückt wird, die drei Beinpaare in panischer Starre eingefroren. Die Schlagwelle des Aufpralls breitet sich als konzentrische Druckringe aus, bei so niedrigen Reynoldszahlen nicht als scharfe Turbulenz, sondern als zähviskose Verzerrung des Brechungsindex, die die Hintergrundelemente wie durch Linsen schwingen lässt. Im Hintergrund ragen Makrophytenstängel als lebende Säulen einer versunkenen Kathedrale empor, ihre parenchymatischen Zellwände filtern das von der Wasseroberfläche einfallende Licht in ein allgegenwärtiges Jadegrün, das jeden Chitinfortsatz, jede fädige Alge und jeden schwebenden Diatomeenfrustulenfragment in den Rang eines leuchtenden Partikels erhebt. Rechts oben flieht eine zweite Naupliuslarve bereits in einem Fluchtsprung, die Antennen rückwärts gepeitscht, ihr einzelnes rotoranges Auge ein winziger Glutpunkt im grünen Tiefenraum — ein stiller Beweis dafür, dass in dieser Welt Jagd und Flucht auf einer Zeitskala von Millisekunden entschieden werden.

In dieser Welt bist du selbst so klein wie ein Nauplius-Larv — jener erste, freischwimmende Entwicklungsschritt der Krebstiere, kaum 120 Mikrometer lang, oval und beinahe unsichtbar im graustichig-grünen Brackwasser der Chesapeake Bay im Morgengrauen. Was dich umgibt, ist keine Leere, sondern ein dichtes, lebendes Medium: Überall hängen feurig-orange Dreifach-Augenflecken im Trüben, jeder ein Naupliusauge aus verschmolzenen Ommatidien, das einzige farbige Organ in einem sonst geisterhaften, chitinhäutigen Körper, dessen Silhouette sich nur durch die silberfadigen Antennenborsten und den geschwungenen Schatten seines Darms zu erkennen gibt. Schräge, goldgelbe Tyndall-Strahlen des tief stehenden Märzdämmers durchdringen von oben rechts die drei Meter suspensionsreicher Wassersäule — kolloidale Tonminerale, Detritus und Phytoplankton streuen das Licht zu scharfen, obliquen Lichtsäulen, in deren Schnitt die transparenten Larvenkörper kurz aufleuchten und die eingelagerten Eyespots zu Glutpunkten auflodern. Zwischen den Nauplii treiben wie verstreute Bronzenadeln die siliziumwandigen Frusteln der pennaten Diatomee *Nitzschia*, facettiert und lichtbrechend, während der Bildhintergrund sich in ein samtiges Blaugrün aus immer kleineren, schwebenden Rubinfunkeln auflöst — jeder einzelne davon ein unsichtbares Leben, das in diesem einen Kubikmillimeter Salzwasser mit Dutzenden anderen teilt, was für ein kurzes Nauplius-Dasein an Welt zur Verfügung steht.

Der Blick richtet sich nach oben, und die gesamte Welt verdichtet sich in einem einzigen optischen Moment: Die Unterseite des Meereis füllt den Horizont wie eine unermessliche Kathedralendecke aus gefrorenem Licht, ihre unebene Fläche von Diatomeen-Biofilmen in warmem Bernstein und Ocker überzogen — lebendige Flecken, die im kalten Blau-Weiß des durchgefilterten Tageslichts glühen wie Buntglas in einem arktischen Dom. Davor, fast zum Greifen nah, steigt ein *Calanus hyperboreus* auf: Sein transparenter Körper ist von einem gewaltigen Lipidvorrat dominiert, einem Wachsester-Reservoir, das backlit im orangeroten Glutlicht einer Glut erstrahlt — biochemisch gespeicherte Sonnenenergie, die der Copepode während der kurzen arktischen Algenblüte akkumuliert hat, um damit Monate langer Dunkelheit in der Tiefsee zu überdauern. Die ausgestreckten Antennulen tasten nach chemischen Signalen aus der Algendecke darüber, während die federartigen Setae einzeln im Gegenlicht erkennbar sind: ein Sensorium, das auf molekulare Gradienten in einer Welt reagiert, die für das Tier gleichzeitig Nahrungsquelle, Oberfläche und Orientierungspunkt ist. Darunter versammeln sich weitere Tiere als kleiner werdende Orangegluten im Cobaltblau — jede ein winziges, aber vollständiges Feuer auf dem langen vertikalen Weg von der Dunkelheit zum Licht —, während die Wassersäule unter uns in absolutes, drückendes Schwarz versinkt.

Vor uns erstreckt sich eine endlose blasse Ebene aus verdichtetem graubeigem Schlick, dessen Oberfläche wie feines Wildleder unter extremer Vergrößerung wirkt – eine mesopelagische Sedimentlandschaft in 150 Metern Tiefe, wo kein Sonnenstrahl je eindringt und die einzige Lichtquelle das Lebewesen selbst ist. Unmittelbar vor uns überquert ein Ostrakode das Substrat, sein cremefarbenes, zweiklappiges Calcitgehäuse kaum größer als ein Sesamkorn, die glatte Oberfläche von einem regelmäßigen Raster dunkler Porenkanäle durchsetzt, die das Kalzit wie ein fein gebohrtes Gitter strukturieren. Aus dem klaffenden Spalt zwischen den Valven sickert kontinuierlich eine biolumineszente Flüssigkeit – eine kalte, gesättigte blaugrüne Leuchtkraft, erzeugt durch die Luciferin-Luciferase-Reaktion des Tieres –, die den unmittelbar umgebenden Meeresboden in einer flachen Scheibe aus ätherischem Türkislicht illuminiert und hinter jedem Foraminiferen-Test und jedem einzelnen Schlickkörnchen winzige Mikroschatten wirft. Feine bedornte Laufbeine greifen in das Sediment, ihre gebogenen Spitzen hinterlassen haarfeine Eindrücke im Ton, während paarige Tastantennen sich vorwärts in die Dunkelheit strecken und entlang ihrer Länge das biolumineszente Leuchten auffangen, sodass sie wie leuchtende Tasthaare erscheinen, die eine Welt ertasten, die jenseits eines einzigen Zentimeters in absolutem Schwarz versinkt.

In diesem Augenblick schwimmst du entlang der Jagdachse selbst, eingebettet in ein Blau von solcher Sättigung, dass es weniger wie Wasser als wie verflüssigtes Licht wirkt — ein tropisches Pelagial in zehn Metern Tiefe, wo die Sonne noch mit voller Kraft ankommt und durch die sanfte Dünung oben in gleitende Kaustik-Gitter gebrochen wird, die sich wie goldene Filigranarbeit über jede Fläche legen. Im scharfen Vordergrund füllt ein männlicher calanider Copepode dein Blickfeld mit architektonischer Präzision: ein ein bis zwei Millimeter langer Tropfen nahezu vollkommener Transparenz, sein einzelnes Naupliusauge leuchtet rubinrot, der bernsteinfarbene Lipidsack glüht warm gegen das kühle Blau, und die modifizierten Antennulen schwingen vorwärts, ihre fadenfeinen Ästhetasken tasten das Pheromonfeld ab, das das Weibchen — dreißig Körperlängen entfernt, kleiner, ebenso gläsern — in ihrer Kielwasserspur hinterlassen hat. Zwischen beiden windet sich ein interpretatives Helixband aus sanfter ceruleanblauer Lumineszenz, das ihre genaue Fluchtbahn in molekularer Chemie ins Wasser eingeschrieben hat, die Ränder ausgefranst wie ausgeatmeter Atem in kalter Luft — ein Phänomen, das in der kalaniden Reproduktionsbiologie seit den 1980er Jahren belegt ist und bei dem hydrophobe Signalmoleküle in einer kohärenten Spur verbleiben, weil die viskosen Kräfte bei diesen Körpergrößen Diffusion deutlich verlangsamen. Im weiteren blauen Raum treiben goldbraune Dinoflagellaten-Kugeln und zierliche Chaetoceros-Ketten wie geometrisches Treibgut, jede Siliciumdioxid-Frustel ein nahezu unsichtbares Scheibchen mit eigenem Regenbogenrand — die photosynthetische Grundlage jener Nahrungskette, deren dramatischster gegenwärtiger Akt sich in deiner unmittelbaren Nähe vollzieht.

Der Blick führt senkrecht nach oben durch achtzig Meter Atlantik, und die Dunkelheit selbst scheint zu leben: Hunderte von *Calanus*-Ruderfußkrebsen steigen in pulsierenden Strömen empor, jeder Körper kaum zwei bis drei Millimeter lang und nahezu unsichtbar, wäre da nicht der bernsteinfarbene Lipidtropfen im Prosom, der wie eine winzige Kohleglut durch das Blauschwarz brennt. Die nächsten Individuen hängen in kristalliner Schärfe — die gefächerten Antennen wie Federparachute ausgebreitet, das einzelne Naupliusauge ein roter Funken, die Schwimmbeine in einem eingefrorenen Schlagrhythmus — während dahinter Hunderte weitere amber-leuchtender Formen zu einem weichen Schwarm verschwimmen, einer senkrecht gestellten Galaxie in geordneter Wallfahrt. Ihr Ziel ist das Snell'sche Fenster direkt über ihnen: eine präzise geschnittene Kreisscheibe aus komprimiertem Tageslicht, noch silberblass, aber am Rand bereits golden werdend mit der herannahenden Sonne. Zwischen den aufsteigenden Tieren treibt mariner Schnee in der Gegenrichtung herab — blassgraue Schleier aus Schleim, Diatomeenfrustreln und Kotballen, die wie Asche zwischen Glühwürmchen fallen — und zwei der nächsten Copepoden haben ihre Aufwärtswanderung kurz unterbrochen, die Antennen auf eine vorbeitreibende Flocke gerichtet, ihre Fresswerkzeuge erzeugen unsichtbare Mikrowirbel, während hinter ihnen die gesamte kalte, unter Druck stehende Wassersäule von aufsteigendem Licht bevölkert wird.

Der Blick schwebt mitten in der Wassersäule eines norddeutschen Flachsees, umhüllt von einem jadegrünen Leuchten, das aus aufgeschwemmtem Chlorophyll und gelösten organischen Stoffen entsteht — das Wasser selbst ist hier kein unsichtbares Medium, sondern eine lebendige Substanz mit Gewicht, Wärme und Tiefe. Über uns komprimiert Snells Fenster den gesamten Himmel in ein gleißendes Oval aus zerbrochenem Silberlicht, von dem diagonale Kaustikstrahlen herabsinken und an treibenden Diatomeen-Ketten, Mineralstaubteilchen und Bakterienaggregaten aufblitzen wie Funken in einem Kathedralenraum aus lebendigem Licht. Die *Daphnia cucullata* um uns herum sind Objekte von verstörender Schönheit: jede tragt einen turmartigen Helmfortsatz aus so dünn gezogenem Chitin, dass er nur als leuchtende Konturlinie existiert, während durch die glasklare Carapax hindurch der smaragdgrün gefüllte Darm, die orangeroten Karotinoid-Eier im Brutraum und das rhythmisch pulsierende Herz sichtbar sind — eine vollständige Biologie hinter transparenten Wänden. Im Vordergrund hat eine Daphnia ihre Fluchtreaktion ausgelöst, den Körper zu einem harten C gebogen, die Antennen am Scheitelpunkt des Kraftschlags eingefroren, und das verdrängte Wasser um sie herum verbiegt die Hintergrundstrukturen in einer kaum wahrnehmbaren Linsen-Verzerrung, physikalisches Zeugnis einer Beschleunigung, die zu den schnellsten im gesamten Tierreich zählt; dahinter materialisiert sich in warmem Silber-Gold-Bokeh die Fischlarve — ihr enormes, dunkles Auge mit goldenem Rand unscharf genug, um zur Textur zu gehören, aber scharf genug, um Absicht zu verraten.

In einer horizontalen Perspektive, kaum höher als ein Sandkorn über dem Grund, durchzieht warmes Bernsteingold jeden Winkel dieser Sphagnum-Welt: das Moorwasser selbst ist eine gesättigte Lösung aus Huminsäuren und Fulvosäuren, die jeden Lichtstrahl von hellem Honig über tiefes Topas bis hin zu mahagonibraunem Dunkel färbt, sodass die sichtbare Welt bereits nach wenigen Zentimetern in warmem Rost versinkt. Im Zentrum schwebt ein *Acanthocyclops robustus*, sein carotinoidgesättigter orangeroter Körper der einzige brennend gesättigte Farbakzent in dieser monochrom warmen Welt — durch das halbdurchsichtige Chitin des Prosoms schimmert der Darm als dunklerer Roststreifen, das einzelne Naupliusauge leuchtet rubinrot, und die lateral gespreizten Antennulen strahlen in feinen goldenen Seten in die Umgebung aus. Rechts erhebt sich der Sphagnum-Stängel wie eine Fensterfront aus lebendigem Glas: die toten Hyalzellen bilden ein sich wiederholendes Gitter aus bernsteinleuchtendem, wassergefülltem Pergament, unterbrochen vom kühlen Chlorophyllgrün der lebenden Photosynthesezellen — ein rhythmischer Wechsel von kalt und warm, der die ganze Wand in ein mittelalterliches Kirchenfenster verwandelt. Über allem spannt sich die Wasseroberfläche als goldgekräuselter Spiegel der Totalreflexion, in dessen Mitte das Snell'sche Fenster den bedeckten Himmel als kühlen Silberkreis hereinlässt — das einzige kalte Licht in diesem uralten, harzigen, zeitlos bernsteinfarbenen Universum.

Der Blick fällt in eine Architektur, die sich über das gesamte Sichtfeld erstreckt: die Maxillarsetae von *Temora longicornis*, aufgereiht wie die Säulen einer versunkenen Kathedrale, jeder Filament ein zylindrischer Chitinstab von zwei bis fünf Mikrometer Durchmesser, der das blaue Umgebungslicht durch seine Wandung leitet und von innen heraus wie erhitztes Bernsteinglas leuchtet. Zwischen den geometrisch präzise gestaffelten Reihen dieser Setae strömt das Wasser in laminaren Bögen einwärts, trägt goldbraune Diatomeenfragmente und organische Detritusflocken auf einer vorherbestimmten Trajektorie Richtung Mundöffnung — eine biologische Turbine, deren Filterspalt auf den Mikrometer kalibriert ist, eng genug um Phytoplanktonzellen zu fangen, weit genug um gelöste Salze passieren zu lassen. Mittendrin, zwischen zwei benachbarten Setae eingeklemmt und vom hydrodynamischen Druck der zulaufenden Strömung leicht gebogen, hängt eine *Chaetoceros*-Kette: drei längliche Siliziumdioxid-Zellen mit gläsernen Lateralstacheln, die das Blaulicht zu zarten Prismenreflexen brechen, ihre Zellwände in derselben Bernsteintransluzenz leuchtend wie die sie fassenden Filterstrukturen selbst. Im Hintergrund liegt absolutes Kobaltblau-Schwarz, durchzogen vom milchigen Schimmer zu kleiner Partikel, um einzeln aufgelöst zu werden — die ozeanische Abenddämmerung, gegen die sich dieses lebende Präzisionsinstrument als warmes Goldgitter abhebt, während die Muskulatur dahinter in langsamen Kontraktionswellen pulsiert und ihre Schatten durch Chitin und Wasser wirft.

Im blaugrünen Dämmerlicht zwanzig Meter unter der Nordseeoberfläche schwebt eine Calanus helgolandicus-Weibchen reglos im Zentrum unseres Blickfelds – ein gläsernes Schiff in milchig-trüber See, umgeben von einer aquamaringrünen Suspension feinster organischer Partikel, die das gefilterte Oberflächenlicht in diffuse Kälte verwandeln. Durch die transparente Chitinwand des Prosoms sollten die vertrauten inneren Strukturen einer gesunden Calanoidin sichtbar sein: orangerote Ovarialgewebe, ein klarer grüner Darmkanal, die geordnete Geometrie eines lebenden Präzisionsinstruments – stattdessen füllt eine dichte, bernsteinbraun-mahagonifarbene Gregarinenmasse die gesamte Körperhöhle aus, ein apikomplexer Einzeller, der das Darmepithel besetzt und sich als trüb-granulierte Opazität gegen die Integumentwand drückt und das Licht absorbiert statt passieren zu lässt. Diese Gregarinen – obligate intrazelluläre und extraluminale Parasiten aus der Gruppe der Apicomplexa – unterdrücken die Reproduktionsleistung ihres Wirts, reduzieren die Lipidspeicherung und verringern die Überlebensrate, ohne das externe Verhalten sofort zu blockieren: Die Antennen spreizen sich noch immer weit in die Wassersäule, die Schwimmfüße schlagen im rhythmischen Takt ihrer namenlosen Pflicht. Hinter dem Tier treibt eine blasse, gelatinöse Dinoflagellatenkugel in der Strömung und sendet für einen Augenblick einen kaltblauen biolumineszenten Puls aus, dessen chemisches Licht kurz die umgebende Trübe in ein spukhaftes Corona taucht – die sanfte Leuchtreklame eines Ökosystems, das funktioniert, als wäre nichts geschehen.

Im Schwebezustand wenige Zentimeter unterhalb der Wasseroberfläche eines Flachsees begegnen wir einem der intimsten Ereignisse der aquatischen Welt: Ein weiblicher *Cyclops* dominiert das Bild wie ein bernsteinfarbenes architektonisches Monument, sein durchscheinendes Prosoma lässt die fadenförmige Darmschlaufe erahnen, während das einzelne Medianauge wie ein Rubin an der Vorderspitze leuchtet. An seinem Urosome hängen zwei prallgefüllte Eisäcke, karotenoidgesättigte Dotterkonzentrate, deren Membran am linken Sack bereits aufgerissen ist — eine hauchzarte irideszente Haut, die sich in Interferenzfarben von Silber und Aquamarin in die grünlich gefilterte Wassersäule wölbt wie zerberstende Seifenfolie. Drei Naupliuslarven treten gleichzeitig aus diesem Geburtskanal: eine noch in embryonale Membranhäutchen gehüllt, deren Setae wie silbervergoldete Spinnfäden im Gegenlicht schimmern, eine im freien Taumeln mit bereits ausfahrenden Antennen- und Mandibularästen, die ersten Schlagzyklen im hochviskosen Nahfeld tastend erprobend, und eine dritte, die schon frei im leuchtenden Grün schwebt — ihr dreilappiges Medianauge ein glutroter Funke, der die Umgebung abtastet, bevor der erste echte Ruderschlag das Tier in die phytoplanktongetrübte, mit organischen Schwebeteilchen übersäte Wassersäule entlässt.

Im Kegel eines einzigen Stirnlampenlichtstrahls, der schräg durch absolute Finsternis fällt, liegt eine *Pseudocandona* — ein stygobionter Ostrakode von weniger als einem Millimeter Länge — wie ein Elfenbeinfragment auf dem blassen Kalkschotter eines unterirdischen Karstbaches. Das zweiklappige Gehäuse ist vollständig depigmentiert und äugenlos, ein Körperplan, der im Laufe geologischer Zeiträume durch Millionen von Jahren ewiger Nacht geformt wurde; die leicht geöffnete Schale gibt einen Fächer außerordentlich feiner Sinnesborsten frei, sogenannte Aesthetasken, deren haarfeine Schatten mit chirurgischer Präzision auf dem Substrat darunter liegen. Der umgebende Untergrund ist mit einem samtartigen weißen Bakterienrasen bedeckt, der die Kalksteingerölle wie ein lebendiges Tuch überzieht — ein chemoautotrophes Ökosystem, das ohne jeden Sonnenstrahl auskommt. Über uns reflektiert die Wasseroberfläche den Lichtstrahl als zitternde Silberscheibe an die Höhlendecke, die einzige Bewegung in einer sonst geologisch erstarrten Welt, während jenseits des Lichtkegels die Dunkelheit nicht bloß tief, sondern physisch absolut ist.

Vor Ihnen hängt ein Körper im Nichts – ein *Gaussia princeps*, kaum vier Millimeter lang, dessen tiefkarmesinrotes Exoskelett in der absoluten Dunkelheit sechshundert Meter unter der Meeresoberfläche beinahe schwarz wirkt, ein Pigment, das jeden streunenden Lichtblitz verschluckt, bevor er das Tier für einen Räuber sichtbar machen könnte. Dann erfassen die Mechanorezeptoren der langen seitlich ausgestreckten Antennen die Druckwelle Ihrer Annäherung, und das ventrale Urosom antwortet: Eine Reihe biolumineszenter Photophoren zündet nacheinander, jedes Organ öffnet sich als kalter aquamarinblauer Punkt aus reiner Photochemie – kein Wärme, nur Photonen –, und in diesem flüchtigen Halbdunkel werden die Segmente des Chitinpanzers sichtbar, die Muskelstränge darunter, die winzigen Borstenkämme der Schwimmbeine, bevor die Dunkelheit alles zurückfordert. Die Architektur dieses Körpers ist gedrungener und schwerer als die seiner epipelagischen Verwandten, deren Gewebe glasdünn und transparent sind; hier, unter einem Druck von etwa sechzig Atmosphären, sprechen Exoskelett und Muskulatur eine andere Sprache der Anpassung. Im Hintergrund markieren ein Dutzend verstreuter blauer Lichtfunken das endlose Volumen der Wassersäule – die Abwehrblitze anderer Organismen, die vom sinkenden Meeresschnee berührt werden, die einzigen Tiefenhinweise in einem Raum, dessen Ausdehnung sich der Vorstellung entzieht.

Der Blick reicht horizontal durch ein Blau, das weniger eine Farbe als ein Zustand der Materie ist — gesättigtes Kobalt im Oberen, übergehend in ein langsam verdunkelndes Indigo in der Tiefe, durchzogen von zitternden Lichtnetzen, die sich über jede transparente Oberfläche legen wie silberne Kaligraphie. Im rechten oberen Bildfeld brennt Snells Fenster als weißgoldene Scheibe durch die Wasseroberfläche, ihr Rand scharf wie geschliffenes Kristall, darunter eine perfekte Spiegeldecke aus Totalreflexion — das gesamte Himmelsgewölbe in einen einzigen leuchtenden Kreis komprimiert. Im Vordergrund materialisiert sich ein *Clausocalanus*-Copepode mehr als Linsenstörung denn als Tier: sein 0,8-Millimeter-Körper nahezu unsichtbar, die Antennen wie gesponnenes Glas in die Wassersäule ragend, einzig ein rubinroter Punkt des Naupliusauges und ein grünbrauner Darmfaden aus verdauten Diatomeen verraten seine Anwesenheit. Etwas tiefer hängt eine *Oithona* mit paarigen, bernsteinfarben leuchtenden Eisäcken wie Laternen unter ihrem Hinterleib, während drei Naupliuslarven in trägen Bögen trudeln, ihre einzelnen orangeroten Medianaugen wie glühende Funken im blauen Licht. Um sie herum treiben eine goldbraune *Thalassiosira*-Diatomenkette, ein Pteropode mit violett-rosafarbenen Flossenschwingen, das silbern-kristalline Strontiumsulfat-Skelett eines Acantharians und eine grau-weiße Wolke aus marinem Schnee — eine gläserne, schwerelosen Gemeinschaft aus Transparenz, Bernstein und Feuer.

Wir liegen aufgeschlagen auf einer rosa-violetten Krustenalge am Boden eines Gezeitentümpels an der Oregonküste – zwei Zentimeter unter der Wasseroberfläche – und blicken auf eine Welt, die sich anfühlt wie eine weitläufige, geologisch alte Landschaft. Vor uns kriechen zwei *Tigriopus californicus*, harpaktikoide Ruderfußkrebse von strahlendem Orangerot, ihre dorsoventral abgeflachten Körper schmiegen sich dem kalzifizierten Plattenwerk der Alge an, während kurze Antennulen den goldbraunen Diatomeen-Biofilm abtasten – ein irisierendes Mosaik aus Bernstein, Ocker und Kupfer, das sich wie ein byzantinisches Pflaster bis zum fernen Beckenrand erstreckt. Das nachmittägliche Pazifiklicht bricht durch die kaum zwei Zentimeter tiefe, kristallklare Wassersäule und erzeugt rasende Kaustiknetze auf dem Algensubstrat – weiß-goldene Lichtbögen, die sich in Sekundenbruchteilen reformieren und die gesamte Oberfläche in flimmerndes Feuer tauchen. Im Hintergrund ragen Seepocken-Vulkankegel wie grauweiße Kalksteintürme auf, zwischen ihnen hängen leuchtend grüne *Ulva*-Fetzen, und direkt über uns öffnet sich das Snell-Fenster als komprimiertes Oval aus gesättigtem Pazifikblau, eingerahmt vom dunklen Granitsaum des Tümpelrands – ein Blick hinaus in eine andere, größere Welt. In diesem winzigen, von Licht durchströmten Kosmos sind diese Krebstierchen keine Kleinigkeiten, sondern gepanzerte Bewohner einer lebendigen, architektonisch komplexen Landschaft.
