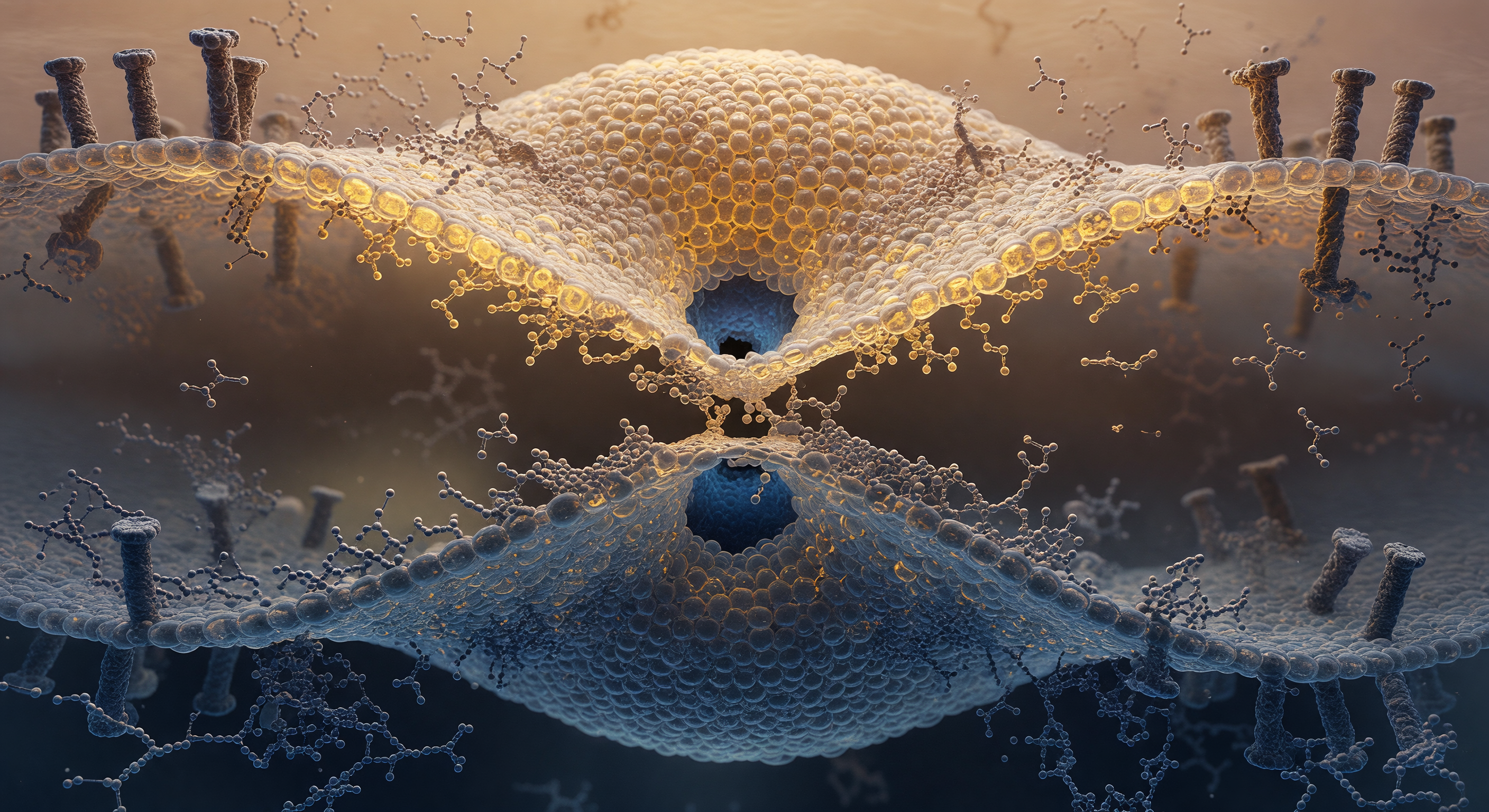
You are suspended at the narrowest point of a molecular hourglass, two nanometers across, where the boundary between a virus and the cell it is invading has temporarily ceased to exist. What surrounds you is not quite membrane and not quite open space but a continuous monolayer of interdigitated lipid — the stalk intermediate of hemifusion, a fleeting topological chimera in which the outer leaflets of a viral envelope above and an endosomal bilayer below have merged into a single disordered throat, their hydrophobic tails briefly exposed at the waist before folding back into the flanking leaflets whose headgroups, individually visible as boulder-sized phosphorylcholine spheres, tremble with violent thermal micro-displacements rather than any smooth or predictable motion. The geometry is stabilized, tenuously, by fusion proteins whose transmembrane anchors are sunk into both bilayers at the stalk periphery, their coiled-coil ectodomains having already snapped into their post-fusion conformation and dragged the two membranes together with forces measured in tens of piconewtons, bending both lipid planes radically inward to feed this junction. At the center of your view, physiological water presses inward from above and below against the last few angstroms of lipid barrier, forming two dark aqueous hemispheres that lean toward each other through the thinning fabric — the pore has not opened yet, but it is becoming inevitable, a thermodynamic commitment unfolding in slow molecular increments that will, within milliseconds, allow viral RNA to cross from one compartment into another and begin the infection of a cell.
Other languages
- Français: Tige de Fusion Membranaire Virale
- Español: Tallo de Fusión Membrana Viral
- Português: Haste de Fusão Membrana Viral
- Deutsch: Viraler Membranfusions-Stiel
- العربية: ساق اندماج الغشاء الفيروسي
- हिन्दी: वायरल झिल्ली संलयन डंठल
- 日本語: ウイルス膜融合茎
- 한국어: 바이러스 막 융합 줄기
- Italiano: Stelo di Fusione Membrana Virale
- Nederlands: Virale Membraanfusie Steel